
Enhanced Lethal Effects of Combined P-tert-Butylcatechol and L-Lysine on Microcystis aeruginosa

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Algae Culture Conditions
2.2. Experimental Design
2.2.1. Individual and Combined Algal Inhibition Effects of TBC and L-Lysine
2.2.2. Research on the Combined Effect of Algal Inhibition Mechanism
2.3. Determination of Algal Cell Density
2.4. Determination of Chlorophyll-a Content and Chlorophyll Fluorescence
2.5. Determination of Cell Survival Status
2.6. Determination of Soluble Protein Contents and Phycobiliprotein
2.7. Determination of Membrane Permeability
2.8. Determination of Enzyme Activity and Microcystins (MCs)
2.9. Statistical Analysis of Data
3. Results
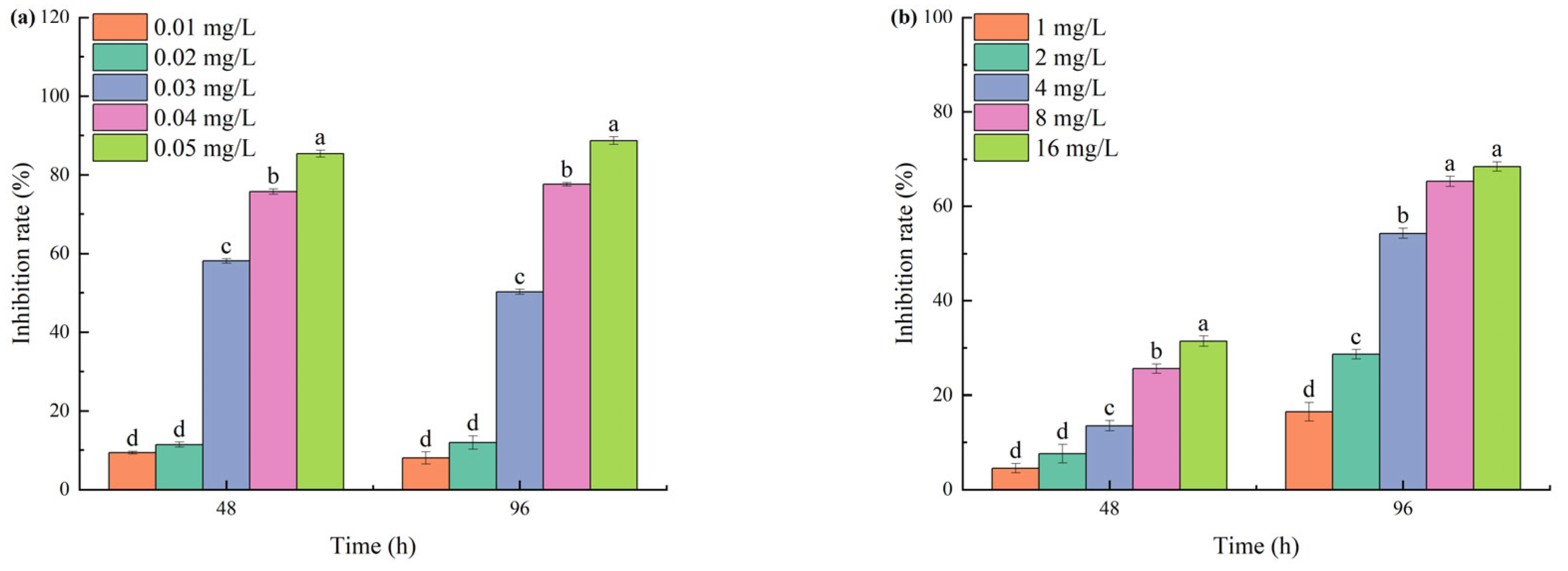
3.1. Effects of TBC and L-Lysine on M. aeruginosa Growth
3.2. Effects of Combined TBC and L-Lysine on M. aeruginosa Growth
3.3. Identification of Viable Microcystis aeruginosa Cells by Flow Cytometry
3.4. Effects of Combined TBC and L-Lysine on the Photosynthetic System of M. aeruginosa
3.4.1. Effects on Chlorophyll Fluorescence Transients
3.4.2. Effects on Chl-a and Soluble Protein
3.4.3. Effects on Phycobiliprotein Content
3.4.4. Effects on PEPC Activity
3.5. Effects of Combined TBC and L-Lysine on the Antioxidant Defense Response of M. aeruginosa
3.6. Effects on Cell Membrane Permeability
3.7. Production and Degradation of MCs
4. Discussion
4.1. Combined Inhibition of M. aeruginosa Growth by TBC and L-Lysine
4.2. Combined TBC and L-Lysine Treatment Impaired the Photosynthetic System of M. aeruginosa
4.3. Oxidative Stress Response of M. aeruginosa to Combined TBC and L-Lysine
4.4. Combined TBC and L-Lysine Altered Membrane Permeability in M. aeruginosa
4.5. TBC and L-Lysine Co-Treatment Modulates Microcystin Synthesis in M. aeruginosa
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Sang, W.L.; Du, C.H.; Liu, X.G.; Ni, L.X.; Li, S.Y.; Xu, J.E.; Chen, X.Q.; Xu, J.; Xu, C. Effect of artemisinin sustained-release algaecide on the growth of Microcystis aeruginosa and the underlying physiological mechanisms. RSC. Adv. 2022, 12, 16094–16104. [Google Scholar] [CrossRef] [PubMed]
- Tanvir, R.U.; Hu, Z.Q.; Zhang, Y.Y.; Lu, J.R. Cyanobacterial community succession and associated cyanotoxin production in hypereutrophic and eutrophic freshwaters. Environ. Pollut. 2021, 290, 118056. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef]
- Lürling, M.; van Oosterhout, F. Effect of selected plant extracts and D- and L-lysine on the cyanobacterium Microcystis aeruginosa. Water 2014, 6, 1807–1825. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Sun, R.; Sun, P.F.; Zhang, J.H.; Esquivel-Elizondo, S.; Wu, Y.H. Microorganisms-based methods for harmful algal blooms control: A review. Bioresour. Technol. 2019, 248, 12–20. [Google Scholar] [CrossRef]
- Luo, Y.; Dao, G.H.; Zhou, G.Q.; Wang, Z.X.; Xu, Z.X.; Lu, X.Y.; Pan, X.J. Effects of low concentration of gallic acid on the growth and microcystin production of Microcystis aeruginosa. Sci. Total Environ. 2024, 916, 169765. [Google Scholar] [CrossRef] [PubMed]
- Li, B.H.; Yin, Y.J.; Kang, L.F.; Feng, L.; Liu, Y.Z.; Du, Z.W.; Tian, Y.J.; Zhang, L.Q. A review: Application of allelochemicals in water ecological restoration--algal inhibition. Chemosphere. 2021, 267, 128869. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Liu, B.Y.; Wang, J.; Gao, Y.N.; Wu, Z.B. Study on the mechanism of allelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte (Myriophyllum spicatum) and its secretion. Aquat. Toxicol. 2010, 98, 196–203. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M.; Murakami, A. Myriophyllum spicatum-released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa. Water Res. 2000, 34, 3026–3032. [Google Scholar] [CrossRef]
- Li, J.M.; Cao, L.R.; Guo, Z.H. Joint effects and mechanisms of luteolin and kaempferol on toxigenic Microcystis growth-Comprehensive analysis on two isomers interaction in binary mixture. J. Environ. Manag. 2022, 312, 114904. [Google Scholar] [CrossRef]
- Laska, E.M.; Meisner, M.; Tang, D.I. Classification of the effectiveness of combination treatments. Stat Med. 1997, 16, 2211–2228. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kouchiwa, T.; Hodoki, Y.; Hotta, K.; Uchida, H.; Harada, K.-I. Distribution and identification of actinomycetes lysing cyanobacteria in a eutrophic lake. J. Appl. Phycol. 1998, 10, 391–397. [Google Scholar] [CrossRef]
- Hehmann, A.; Kaya, K.; Watanabe, M.M. Selective control of Microcystis using an amino acid-a laboratory assay. J. Appl. Phycol. 2002, 14, 85–89. [Google Scholar] [CrossRef]
- Qiu, Y.T.; Wang, Z.H.; Liu, F.; Liu, J.X.; Tan, K.T.; Ji, R.B. Inhibition of Scenedesmus quadricauda on Microcystis flos-aquae. Appl. Microbiol. Biotechnol. 2019, 103, 5907–5916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Sun, R.Y.; Du, W.; Meng, N.; Shao, Y.; Li, C.L. Catalytic Synthesis of p-tert-Butylcatechol by Ionic Liquid. Energy Procedia 2012, 16, 1090–1094. [Google Scholar] [CrossRef]
- Dahedl, E.K.; Hancock, T.L.; Kratz, M.A.; Urakawa, H. A combination cyanobacterial treatment approach using hydrogen peroxide and L-lysine successfully improved the removal efficiency of toxic cyanobacteria. J. Environ. Manag. 2025, 373, 123503. [Google Scholar] [CrossRef]
- Kong, F.C.; Zhao, Q.M.; Wen, W.K.; He, P.M.; Shao, L. Allelopathic effects and mechanism of kaempferol on controlling Microcystis aeruginosa blooms. Mar. Pollut. Bull. 2025, 217, 118116. [Google Scholar] [CrossRef]
- Zhang, M.; Kong, F.X.; Xing, P.; Tan, X. Effects of interspecific interactions between Microcystis aeruginosa and Chlorella pyrenoidosa on their growth and physiology. Int. Rev. Hydrobiol. 2007, 92, 281–290. [Google Scholar] [CrossRef]
- He, L.; Lin, Z.Y.; Wang, Y.M.; He, X.J.; Zhou, J.; Guan, M.Q.; Zhou, J. Facilitating harmful algae removal in fresh water via joint effects of multi-species algicidal bacteria. J. Hazard. Mater. 2021, 403, 123662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.H.; Chen, W.; Li, H.Q.; Yang, J.Y.; Zha, D.M.; Duan, Y.Q.; Hozzein, W.N.; Xiao, M.; Gao, R.; Li, W.J. L-valine, an antialgal amino acid from Streptomyces jiujiangensis JXJ 0074T. Appl. Microbiol. Biotechnol. 2016, 100, 4627–4636. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.T.; Jiang, X.X.; Xu, H.Z.; Liu, C.R.; Song, Y.Z.; Pan, K.Q.; Wang, L.X.; Du, L.; Liu, H.X. Effects of 4-tert-butylpyrocatechol and tea polyphenol on growth, physiology and antioxidant responses in Microcystis aeruginosa. Aquat. Toxicol. 2023, 260, 106541. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, M.; Park, W. Unlocking the mystery of lysine toxicity on Microcystis aeruginosa. J. Hazard. Mater. 2023, 448, 130932. [Google Scholar] [CrossRef]
- Zimbai, P.V.; Dionigi, C.P.; Brashear, S.S. Selective toxicity of exogenous L-lysine to cyanobacteria, relative to a chlorophyte and a diatom. Phycologia 2001, 40, 483–486. [Google Scholar] [CrossRef]
- Noh, W.; Kim, J.; Lee, S.J.; Ryu, B.G.; Kang, C.M. Harvesting and contamination control of microalgae Chlorella ellipsoidea using the bio-polymeric flocculant A-poly-L-lysine. Bioresour. Technol. 2018, 249, 206–211. [Google Scholar] [CrossRef]
- Zuo, S.P.; Zhou, S.B.; Ye, L.; Ma, S.M. Synergistic and antagonistic interactions among five allelochemicals with antialgal effects on bloom-forming Microcystis aeruginosa. Ecol. Eng. 2016, 97, 486–492. [Google Scholar] [CrossRef]
- Inderjit; Streibig, J.C.; Olofsdotter, M. Joint action of phenolic acid mixtures and its significance in allelopathy research. Physiol. Plant. 2002, 114, 422–428. [Google Scholar] [CrossRef]
- Gao, Y.N.; Liu, B.Y.; Ge, F.J.; He, Y.; Lu, Z.Y.; Zhou, Q.H.; Zhang, Y.Y.; Wu, Z.B. Joint effects of allelochemical nonanoic acid, N-phenyl-1-naphtylamine and caffeic acid on the growth of Microcystis aeruginosa. Allelopathy J. 2015, 32, 249–258. [Google Scholar]
- Yang, L.H.; Ying, G.G.; Su, H.C.; Stauber, J.L.; Adams, M.S.; Binet, M.T. Growth inhibiting effects of 12 antibacterial agents and their mixtures on the freshwater microalga Pseudokirchneriella subcapitata. Environ. Toxicol. Chem. 2008, 27, 1201–1208. [Google Scholar] [CrossRef]
- Ouyang, P.; Wang, C.; Wang, P.; Gan, X.; Wang, X.; Yang, C. Effects of mixed allelochemicals on the growth of Microcystis aeruginosa, microcystin production, extracellular polymeric substances, and water quality. Water 2020, 12, 1861. [Google Scholar] [CrossRef]
- Wang, R.; Hua, M.; Yu, Y.; Zhang, M.; Xian, Q.M.; Yin, D.Q. Evaluating the effects of allelochemical ferulic acid on Microcystis aeruginosa by pulse-amplitude-modulated (PAM) fluorometry and flow cytometry. Chemosphere 2016, 147, 264–271. [Google Scholar] [CrossRef]
- Huang, S.Z.; Zhu, J.Y.; Zhang, L.; Peng, X.; Zhang, X.Y.; Ge, F.J.; Liu, B.Y.; Wu, Z.B. Combined Effects of Allelopathic Polyphenols on Microcystis aeruginosa and Response of Different Chlorophyll Fluorescence Parameters. Front. Microbiol. 2020, 11, 614570. [Google Scholar] [CrossRef]
- Shao, J.H.; Liu, D.M.; Gong, D.X.; Zeng, Q.R.; Yan, Z.Y.; Gu, J.D. Inhibitory effects of sanguinarine against the cyanobacterium Microcystis aeruginosa NIES-843 and possible mechanisms of action. Aquat. Toxicol. 2013, 142–143, 257–263. [Google Scholar] [CrossRef]
- Sekar, N.; Ramasamy, R.P. Recent advances in photosynthetic energy conversion. J. Photochem. Photobiol. C Photochem. Rev. 2015, 22, 19–33. [Google Scholar] [CrossRef]
- Tian, L.L.; Chen, M.; Ren, C.Y.; Wang, Y.Y.; Li, L. Anticyanobacterial effect of L-lysine on Microcystis aeruginosa. RSC Adv. 2018, 8, 21606–21612. [Google Scholar] [CrossRef]
- Ko, S.; Jeong, Y.; Cho, S.; Lee, E.; Jeong, B.; Baek, S.; Oh, B.; Ahn, C.; Oh, H.; Cho, B.; et al. Functional role of a novel algicidal compound produced by Pseudoruegeria sp. M32A2M on the harmful algae Alexandrium catenella. Chemosphere 2022, 300, 134535. [Google Scholar] [CrossRef]
- Padgett, M.P.; Krogmann, D.W. Regular paper Large scale preparation of pure phycobiliproteins. Photosynth. Res. 1987, 11, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; Podestá, F.E. The functional organization and control of plant respiration. Crit. Rev. Plant Sci. 2006, 25, 159–198. [Google Scholar] [CrossRef]
- Zhao, Q.M.; Jiang, R.T.; Shi, Y.X.; Shen, A.L.; He, P.M.; Shao, L. Allelopathic Inhibition and Mechanism of Quercetin on Microcystis aeruginosa. Plants 2023, 12, 1808. [Google Scholar] [CrossRef]
- Hong, Y.; Hu, H.Y.; Xie, X.; Sakoda, A.; Sagehashi, M.; Li, F.M. Gramine-induced growth inhibition, oxidative damage and antioxidant responses in freshwater cyanobacterium Microcystis aeruginosa. Aquat. Toxicol. 2009, 91, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.M.; Ren, L.J.; Bi, Z.Q.; Ji, X.J.; Zhao, Q.Y.; Huang, H. Adaptive evolution of microalgae Schizochytrium sp. under high salinity stress to alleviate oxidative damage and improve lipid biosynthesis. Bioresour. Technol. 2018, 267, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Belchik, S.M.; Xun, L. S-glutathionyl-(chloro)hydroquinone reductases: A new class of glutathione transferases functioning as oxidoreductases. Drug Metab. Rev. 2011, 43, 307–316. [Google Scholar] [CrossRef]
- Cano-Europa, E.; Ortiz-Butrón, R.; Gallardo-Casas, C.A.; Blas-Valdivia, V.; Pineda-Reynoso, M.; Olvera-Ramírez, R.; Franco-Colin, M. Phycobiliproteins from Pseudanabaena tenuis rich in c-phycoerythrin protect against HgCl2-caused oxidative stress and cellular damage in the kidney. J. Appl. Phycol. 2010, 22, 495–501. [Google Scholar] [CrossRef]
- He, Q.; Kang, L.; Sun, X.F.; Jia, R.X.; Zhang, Y.; Ma, J.S.; Li, H.; Ai, H.N. Spatiotemporal distribution and potential risk assessment of microcystins in the Yulin River, a tributary of the Three Gorges Reservoir, China. J. Hazard. Mater. 2018, 347, 184–195. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Arora, K.; Kohli, R.K. α-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot. 2006, 98, 1261–1269. [Google Scholar] [CrossRef]
- Luisa, C.; Sgherri, M.; Logcini, B.; Puliga, S.; Navari-Izzot, F. Atioxidant system in Sporobolus stapfianus: Changes in response to desiccation and rehydration. Phytochemistry 1994, 35, 561–565. [Google Scholar] [CrossRef]
- Noctor, G.; Gomez, L.; Lè Ne Vanacker, H.; Foyer, C.H. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. J. Exp. Bot. 2002, 53, 1283–1304. [Google Scholar] [CrossRef]
- Liu, H.Y.; Chen, S.N.; Zhang, H.H.; Wang, N.; Ma, B.; Liu, X.; Niu, L.M.; Yang, F.; Xu, Y.; Zhang, X.L. Effects of copper sulfate algaecide on the cell growth, physiological characteristics, the metabolic activity of Microcystis aeruginosa and raw water application. J. Hazard. Mater. 2023, 445, 130604. [Google Scholar] [CrossRef]
- Liu, D.D.; Liu, H.J.; Wang, S.T.; Chen, J.Z.; Xia, Y.L. The toxicity of ionic liquid 1-decylpyridinium bromide to the algae Scenedesmus obliquus: Growth inhibition, phototoxicity, and oxidative stress. Sci. Total Environ. 2018, 622–623, 1572–1580. [Google Scholar] [CrossRef]
- Chen, Y.D.; Zhao, C.; Zhu, X.Y.; Zhu, Y.; Tian, R.N. Multiple inhibitory effects of succinic acid on Microcystis aeruginosa: Morphology, metabolomics, and gene expression. Environ. Technol. 2021, 43, 3121–3130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.T.; Zheng, C.Y.; Hu, W.; Xu, W.W.; Wang, H.F. The allelopathy and allelopathic mechanism of phenolic acids on toxic Microcystis aeruginosa. J. Appl. Phycol. 2010, 22, 71–77. [Google Scholar] [CrossRef]
- Kulabhusan, P.K.; Campbell, K. Physico-chemical treatments for the removal of cyanotoxins from drinking water: Current challenges and future trends. Sci. Total Environ. 2024, 917, 170078. [Google Scholar] [CrossRef] [PubMed]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of Microcystis strain PCC 7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef]
- Li, J.M.; Hu, J.Q.; Cao, L.R.; Yuan, Y. Growth, physiological responses and microcystin-production/-release dynamics of Microcystis aeruginosa exposed to various luteolin doses. Ecotoxicol. Environ. Saf. 2020, 196, 110540. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.I.; Ho, L.; Brookes, J.D. Effect of chlorination on Microcystis aeruginosa cell integrity and subsequent microcystin release and degradation. Environ. Sci. Technol. 2007, 41, 4447–4453. [Google Scholar] [CrossRef]
- Martin, W.; Christian, S. Indirect photolysis of cyanotoxins: One possible mechanism for their low persistence. Wat. Res. 1999, 33, 1159–1164. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Liang, W.Y.; Zhao, Y.; Li, F.Z.; Cao, J.C.; Hu, S.J. Generation and release of Microcystin-lR by Microcystis aeruginosa under hydroquinone inhibition. Environ. Sci. 2014, 35, 2294–2299. (In Chinese) [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae-Inhibiting Substances | Concentration (mg/L) | Inhibition Effect | Days (d) | Cost (¥/m3) | References |
|---|---|---|---|---|---|
| Luteolin and kaempferol | 3.5 + 6.5 | 85% | 14 | 197.4 | [12] |
| Artemisinin, nonanoic acid, malonic acid, and ethyl acetate | 3.94 + 6.27 + 8.2 + 6.38 | more than 80% | 7 | 31 | [31] |
| Nonanoic acid and N-Phenyl-1-naphtylamine | 1.25 + 1.25 | 85.43% | 7 | 255 | [29] |
| TBC and L-lysine | 0.04 + 1 | 93.17% | 4 | 0.28 | This research |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, H.; Jiao, G.; Jiang, R.; Shen, Y.; He, P.; Shao, L. Enhanced Lethal Effects of Combined P-tert-Butylcatechol and L-Lysine on Microcystis aeruginosa. Biology 2025, 14, 655. https://doi.org/10.3390/biology14060655
Jiao H, Jiao G, Jiang R, Shen Y, He P, Shao L. Enhanced Lethal Effects of Combined P-tert-Butylcatechol and L-Lysine on Microcystis aeruginosa. Biology. 2025; 14(6):655. https://doi.org/10.3390/biology14060655
Chicago/Turabian StyleJiao, Heyun, Gangwei Jiao, Ruitong Jiang, Yifei Shen, Peimin He, and Liu Shao. 2025. "Enhanced Lethal Effects of Combined P-tert-Butylcatechol and L-Lysine on Microcystis aeruginosa" Biology 14, no. 6: 655. https://doi.org/10.3390/biology14060655
APA StyleJiao, H., Jiao, G., Jiang, R., Shen, Y., He, P., & Shao, L. (2025). Enhanced Lethal Effects of Combined P-tert-Butylcatechol and L-Lysine on Microcystis aeruginosa. Biology, 14(6), 655. https://doi.org/10.3390/biology14060655