
Study of the Behavioral Characteristics of Lutjanus erythropterus on the Basis of the Distance and Structure of Artificial Reefs



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Culture Conditions
2.2. Experimental Setup
2.2.1. Reef Selection for the Independent Effect Distance
2.2.2. Reef Selection for Behavioral Research
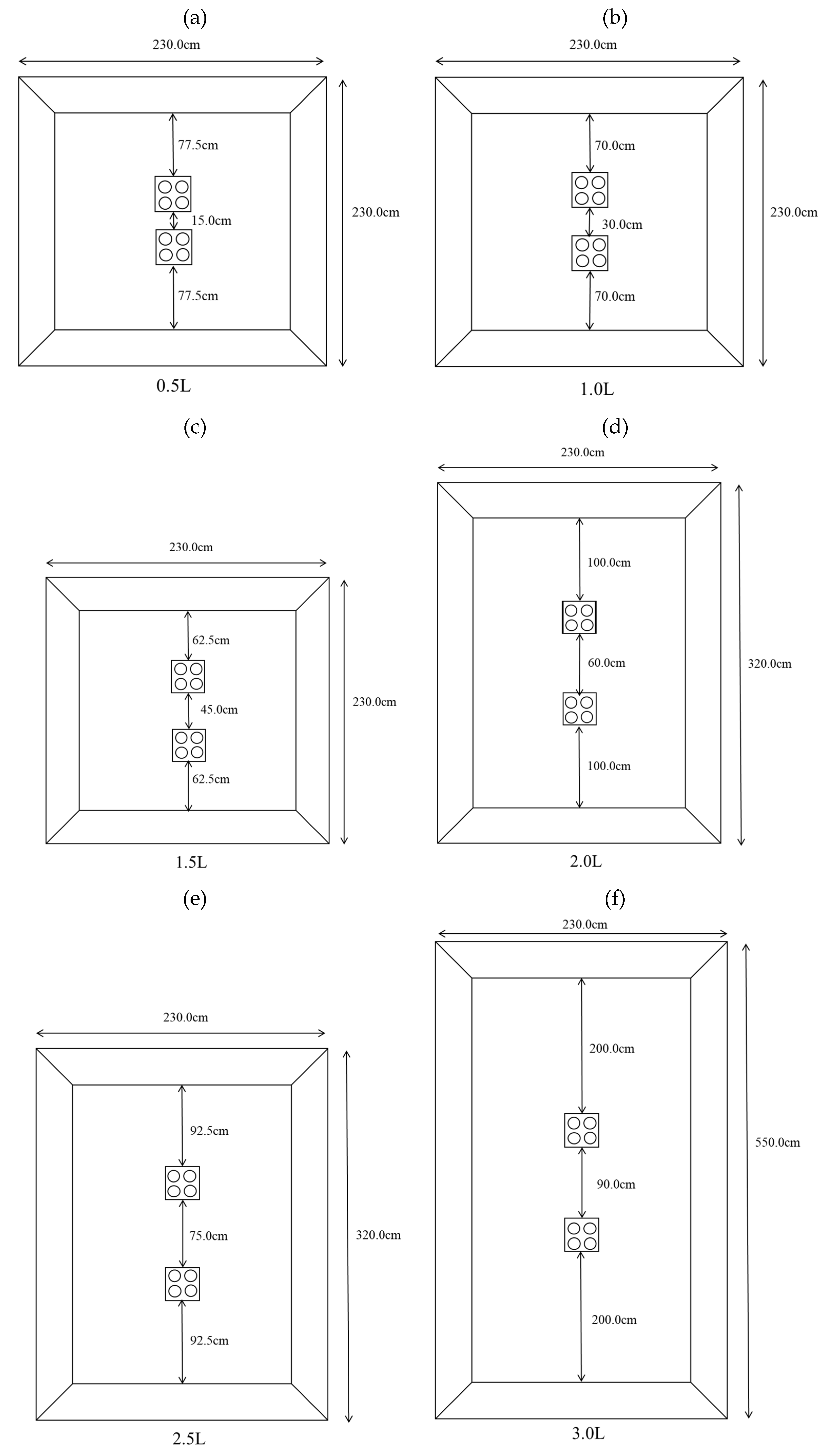
2.3. Experimental Reef Placement
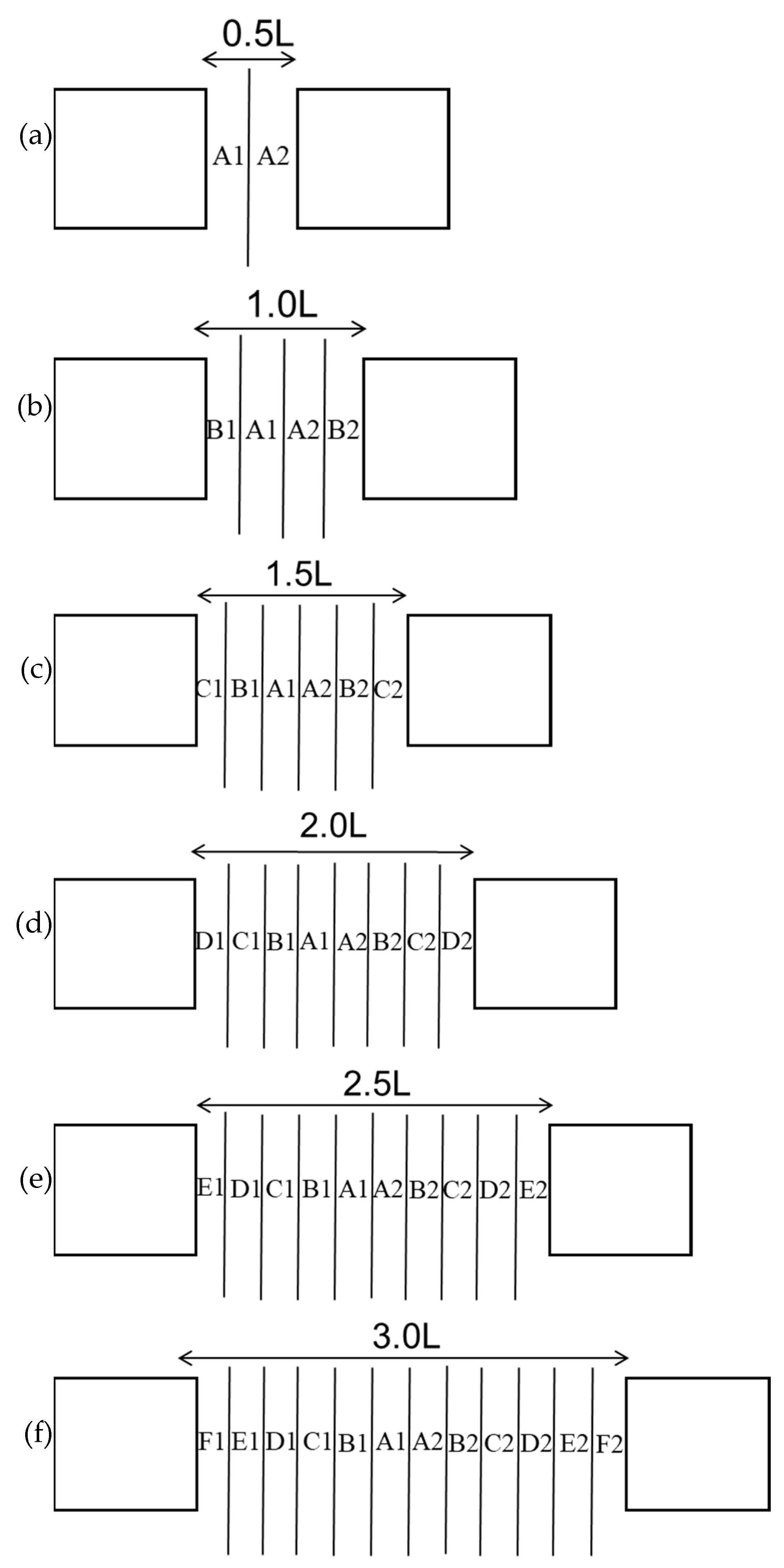
2.3.1. Determination of the Distance at Which Reefs Result in Independent Effects in Fish
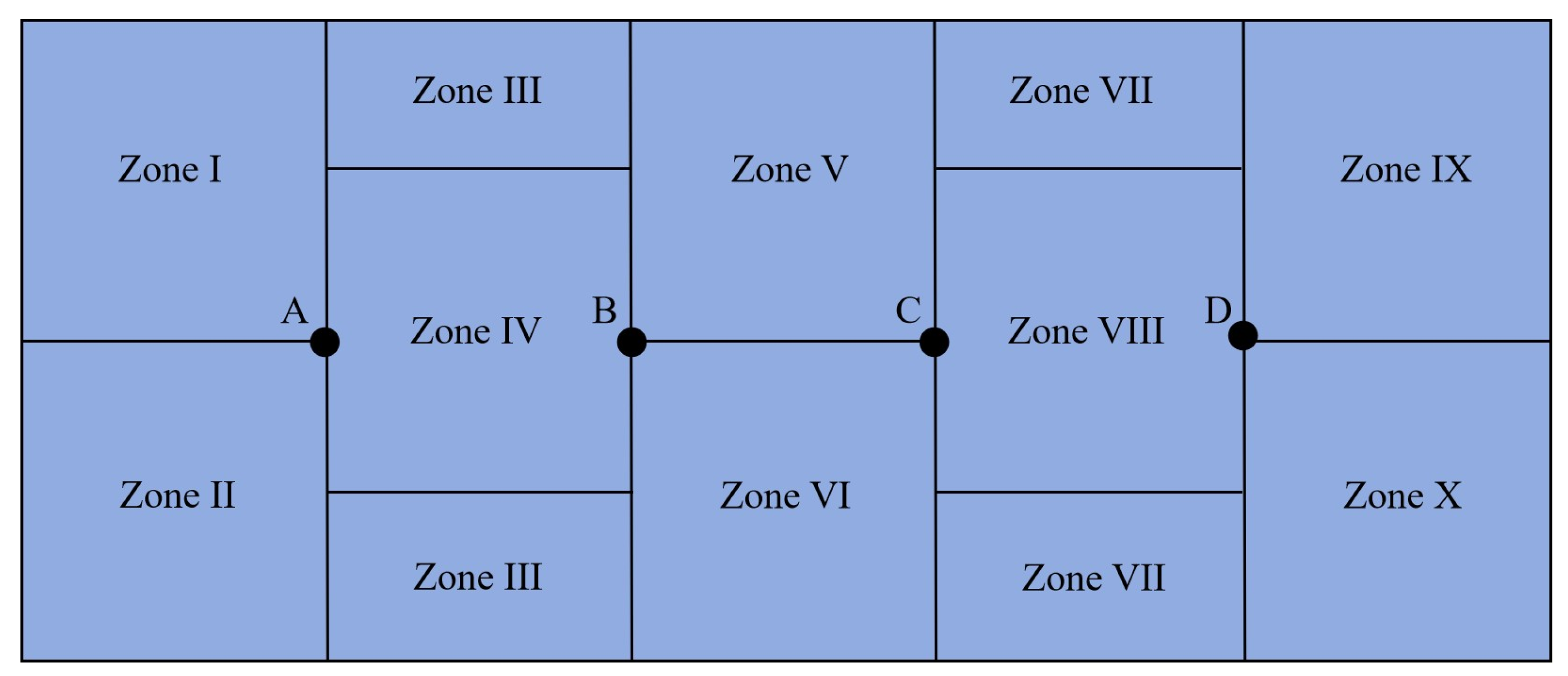
2.3.2. Zoning of Tank
2.4. Experimental Conditions
2.5. Experimental Methods
2.5.1. The Independent Effect Distance of Fish Reefs
2.5.2. Experiments on Two Types of Reef Models
2.5.3. Experiments on Four Types of Reef Models
2.6. Statistical and Analytical Methods
3. Results
3.1. Behavioral Characteristics of Juvenile Lutjanus erythropterus
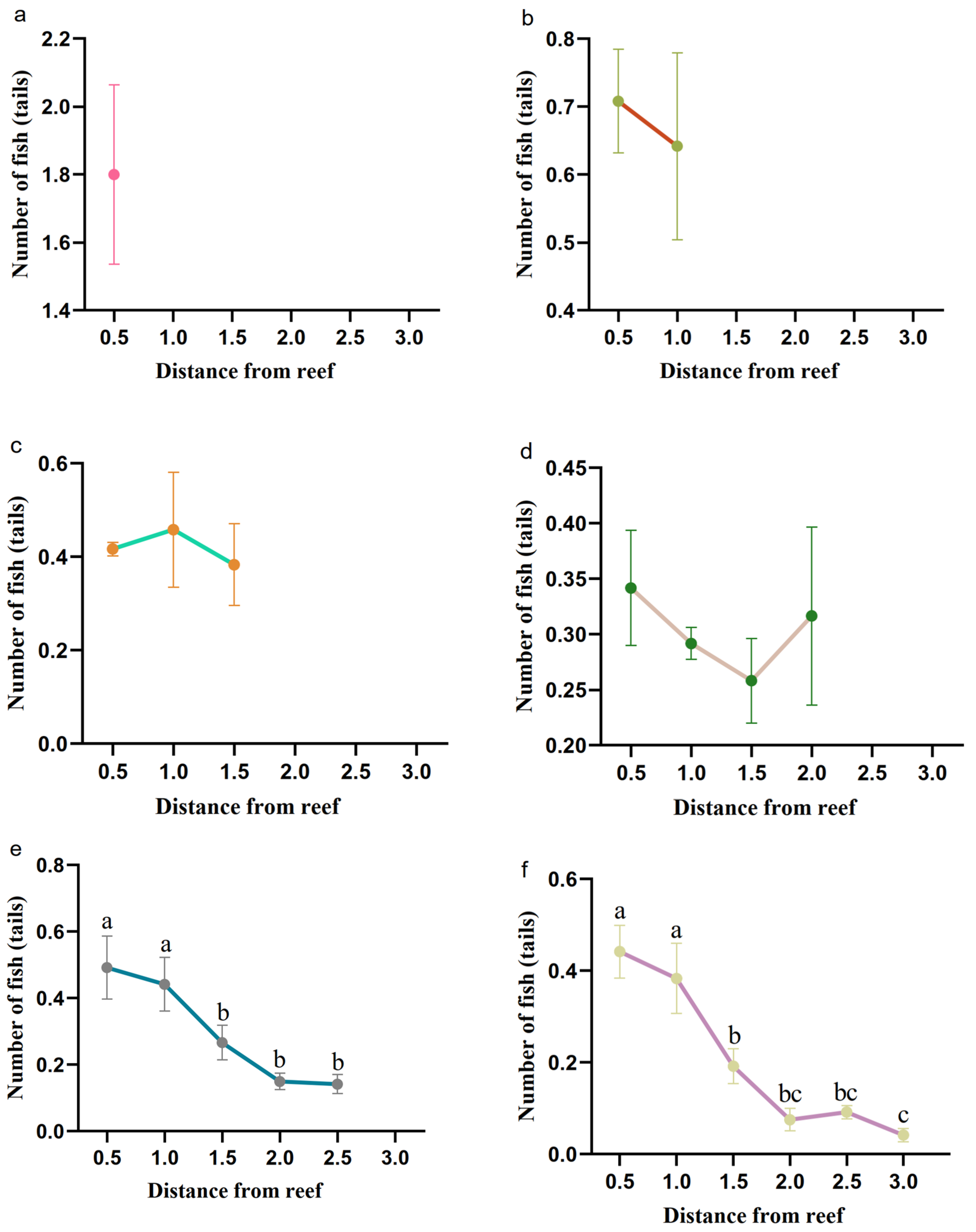
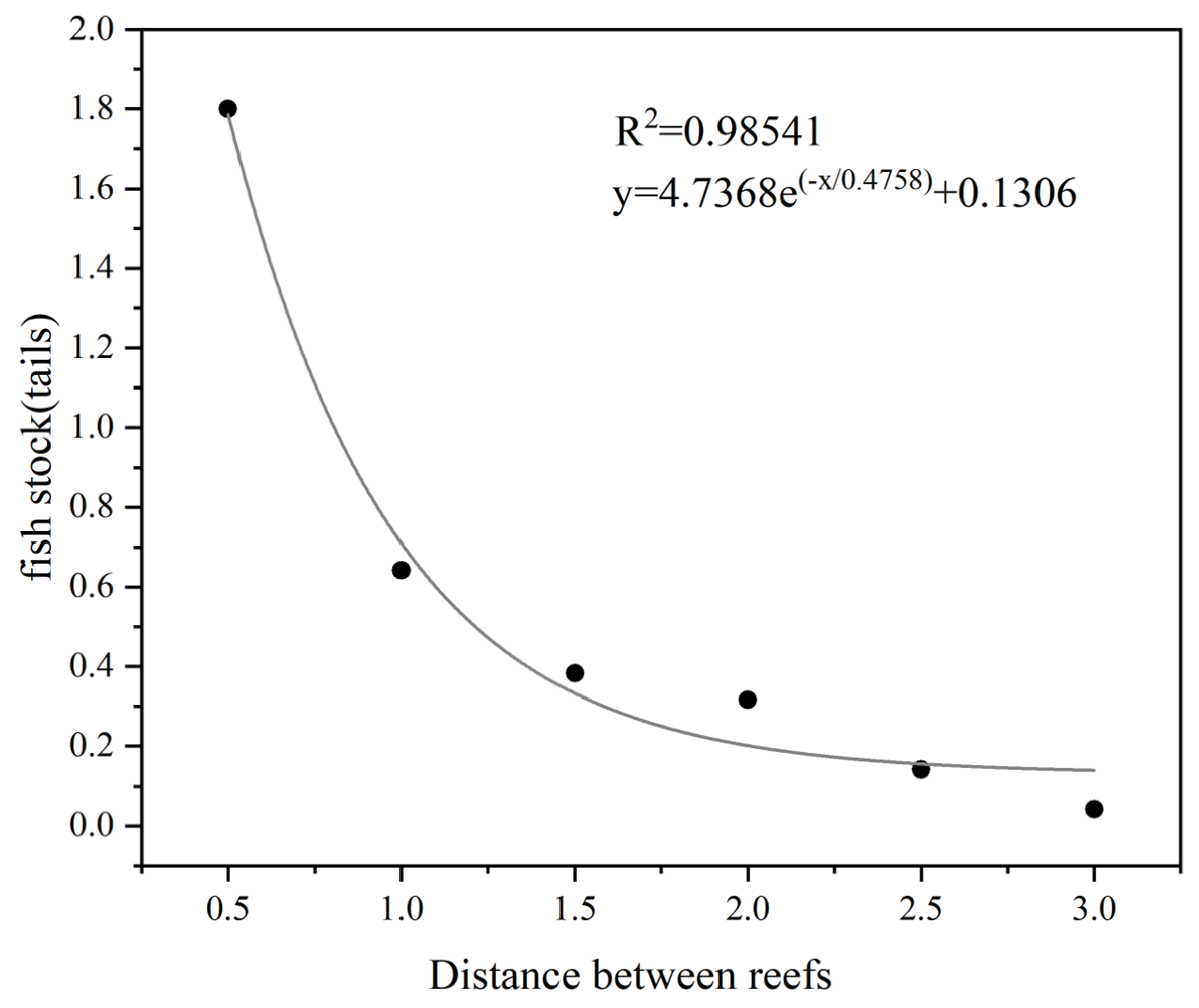
3.1.1. Determination of the Independent Effect Distance of Fish Reefs
3.1.2. Preference of L. erythropterus for Fish Reef
3.2. Determination of the Distances of the Fish Reefs to Retain the Independent Effects
3.3. Behavioral Characteristics of the Two Types of Artificial Fish Reef Models
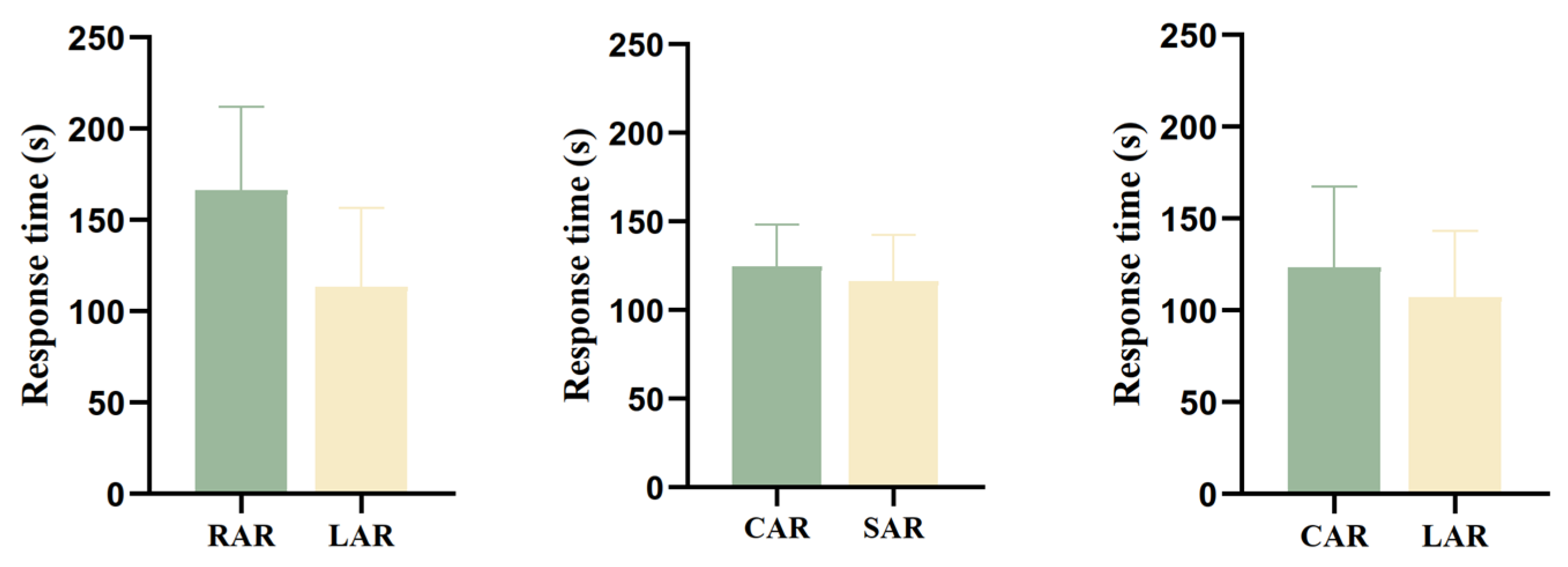
3.3.1. Response Time
3.3.2. Mean Distribution Rate
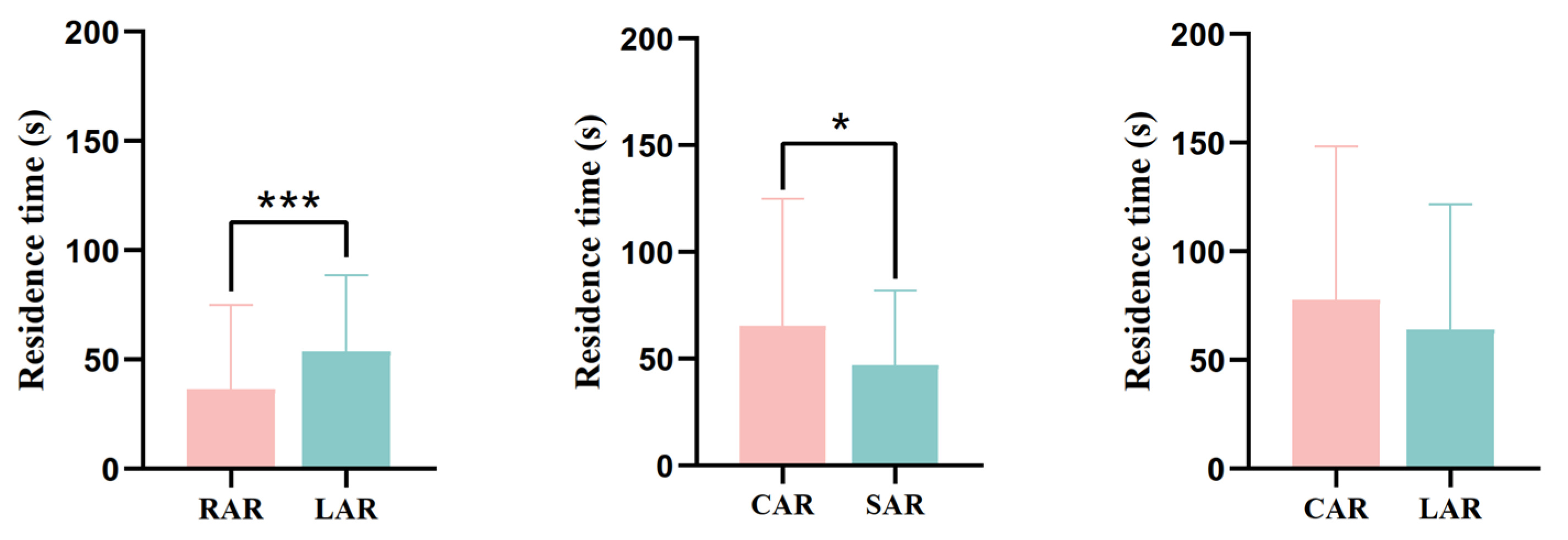
3.3.3. Residence Time
3.4. Behavioral Characteristics of the Four Types of Artificial Fish Reef Models
3.4.1. Response Time
3.4.2. Mean Distribution Rate
3.4.3. Residence Time
4. Discussion
4.1. Determination of the Independent Effect of Distance of Fish Reefs and Analysis of Ecological Effects
4.2. Analysis of the Preference of Juvenile Lutjanus erythropterus for Different Artificial Reef Models
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, M.J.; Guo, Y.; Qin, C.X.; Xin, Y.; Zhao, X.Y.; Yu, G.; Ma, Z.H.; Yang, Y.K. Investigation on behavioral preferences of Lutjanus erythropterus juvenile towards artificial reef models with different pore shapes and sizes. South China Fish. Sci. 2024, 20, 42–52. [Google Scholar]
- Guo, Y.; Zhang, S.Y.; Lin, J. Study on unit reef construction mode based on upwelling effects. South China Fish. Sci. 2020, 16, 71–79. [Google Scholar]
- Mancini, I.; Azzola, A.; Bianchi, C.N.; Capello, M.; Cutroneo, L.; Morri, C.; Oprandi, A.; Montefalcone, M. Habitat Fragmentation Enhances the Difference between Natural and Artificial Reefs in an Urban Marine Coastal Tract. Diversity 2024, 16, 316. [Google Scholar] [CrossRef]
- Komyakova, V.; Chamberlain, D.; Jones, P.G.; Swearer, E.S. Assessing the performance of artificial reefs as substitute habitat for temperate reef fishes: Implications for reef design and placement. Sci. Total Environ. 2019, 668, 139–152. [Google Scholar] [CrossRef]
- Xue, D.W.; Wang, C.Y.; Huang, T.; Pan, Y.; Zhang, N.; Zhang, L.F. Flow field effects and physical stability of pyramidal artificial reef with different slope angles. Ocean Eng. 2023, 283, 115059. [Google Scholar] [CrossRef]
- Hylkema, A.; Debrot, A.O.; Osinga, R.; Bron, P.S.; Heesink, D.B.; Izioka, A.K.; Reid, C.B.; Rippen, J.C.; Treibitz, T.; Yuval, M.; et al. Fish assemblages of three common artificial reef designs during early colonization. Ecol. Eng. 2020, 157, 105994. [Google Scholar] [CrossRef]
- Sentosa, B.W.; Nurruhwati, I.; Apriliani, M.I.; Khan, M.A.A. Distribution of Blacktip Reef Sharks (Carcharinus melanopterus) Based on Habitat Characteristics by the Baited Remote Underwater Video (BRUV) Method in Morotai Waters of North Maluku. Asian J. Fish. Aquat. Res. 2020, 7, 34–50. [Google Scholar] [CrossRef]
- Dominic, A.B.; Consuelo, M.S.; Dan, E.; Erika, G. Assessing Caribbean Shallow and Mesophotic Reef Fish Communities Using Baited-Remote Underwater Video (BRUV) and Diver-Operated Video (DOV) Survey Techniques. PLoS ONE 2016, 11, e0168235. [Google Scholar]
- Grab, H.; Brokaw, J.; Anderson, E.; Gedlinske, L.; Gibbs, J.; Wilson, J.; Loeb, G.; Isaacs, R.; Poveda, K.; Diamond, S. Habitat enhancements rescue bee body size from the negative effects of landscape simplification. J. Appl. Ecol. 2019, 56, 2144–2154. [Google Scholar] [CrossRef]
- Agostini, S.; Harvey, P.B.; Wada, S.; Kon, K.; Milazzo, M.; Inaba, K.; Hall-Spencer, J.M. Ocean acidification drives community shifts towards simplified non-calcified habitats in a subtropical−temperate transition zone. Sci. Rep. 2018, 8, 11354. [Google Scholar] [CrossRef]
- Emadi, M.C.; Bean, T.P.; Mager, M.E. Swimming Performance Assessments of Fish Species of Greatest Conservation Need to Inform Future Stream Crossing Designs in Texas. Fishes 2024, 9, 234. [Google Scholar] [CrossRef]
- Huang, J.L.; Li, J.; Li, Y.; Gong, P.H.; Guan, C.T.; Xia, X. Flow Field Characteristics of Multi-Trophic Artificial Reef Based on Computation Fluid Dynamics. J. Ocean Univ. China 2024, 23, 317–327. [Google Scholar] [CrossRef]
- Villavicencio, B.C.; Cascon, M.H.; Rossi, S. Artificial Reefs around the World: A Review of the State of the Art and a Meta-Analysis of Its Effectiveness for the Restoration of Marine Ecosystems. Environments 2023, 10, 121. [Google Scholar] [CrossRef]
- Layman, C.A.; Allgeier, J.E. An ecosystem ecology perspective on artificial reef production. J. Appl. Ecol. 2020, 57, 2139–2148. [Google Scholar] [CrossRef]
- Neven, K.; Frane, M.; Linda, K.L.; Kristijan, H.; Tina, P.P.; Tatjana, M.; Tea, G. Influence of cognitive performance and swimming capacities on selection of youth water polo players to national team. J. Sports Med. Phys. Fit. 2023, 63, 34–41. [Google Scholar]
- Zagars, M.; Ikejima, K.; Arai, N.; Mitamura, H.; Ichikawa, K.; Yokota, T.; Tongnunui, P. Migration patterns of juvenile Lutjanus argentimaculatus in a mangrove estuary in Trang province, Thailand, as revealed by ultrasonic telemetry. Environ. Biol. Fish. 2012, 94, 377–388. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Dong, J.Y.; Zhang, X.M.; Zhang, X.M. Effects of environmental enrichment on the distribution of Sebastes schlegelii in early developmental stages. Acta Ecol. Sin. 2018, 38, 8223–8233. [Google Scholar]
- Granneman, J.E.; Steele, M.A. Effects of reef attributes on fish assemblage similarity between artificial and natural reefs. CES J. Mar. Sci. 2015, 72, 2385–2397. [Google Scholar] [CrossRef]
- Jiang, Z.Y. Numerical Simulation of Hydrodynamics for Artificial Reefs; Ocean University of China: Qingdao, China, 2009. [Google Scholar]
- Harasti, D.; Malcolm, H.; Gallen, C.; Coleman, M.A.; Jordan, A.; Knott, N.A. Appropriate set times to represent patterns of rocky reef fishes using baited video. J. Exp. Mar. Biol. Ecol. 2015, 463, 173–180. [Google Scholar] [CrossRef]
- Lowry, M.; Folpp, H.; Gregson, M.; Suthers, I. Comparison of baited remote underwater video (BRUV) and underwater visual census (UVC) for assessment of artificial reefs in estuaries. J. Exp. Mar. Biol. Ecol 2012, 416–417, 243–253. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhao, Q.; Qi, Z.L.; Zhang, X.W.; Ye, M.; Cui, C.; Shi, B.J.; Hou, R. Comparison of stability and fish collection effects of several different types of artificial reefs. Hebei Fish. 2021, 1, 4–10. [Google Scholar]
- Zhou, Y.B.; Cai, W.G.; Chen, H.G.; Chen, P.M.; Lv, G.M.; Jia, X.P. Attraction effect of various artificial reef models on Plectorhynchus cinctus. J. Trop. Oceanogr. 2010, 29, 103–107. [Google Scholar]
- Chen, Y.; Yu, C.Q.; Zhang, G.S.; Zhang, S. The environmental function and fish gather effect of atificial reefs. J. Dalian Ocean Univ. 2002, 17, 64–69. [Google Scholar]
- Lin, J.; Zhang, S.Y. Research advance on physical stability and ecological effectsof artificial reef. Mar. Fish. 2006, 28, 257–262. [Google Scholar]
- Campbell, M.D.; Rose, K.; Boswell, K.; Cowan, J. Individual-based modeling of an artificial reef fish community: Effects of habitat quantity and degree of refuge. Ecol. Model. 2011, 222, 3895–3909. [Google Scholar] [CrossRef]
- Liu, T.-L.; Su, D.-T. Numerical analysis of the influence of reef arrangements on artificial reef flow fields. Ocean Eng. 2013, 74, 81–89. [Google Scholar] [CrossRef]
- Zhang, J.T.; Zhu, L.X.; Liang, Z.L.; Sun, L.Y.; Nie, Z.Y.; Wang, J.H.; Xie, W.D.; Jiang, Z.Y. Numerical study of efficiency indices to evaluate the effect of layout mode of artificial reef unit on flow field. J. Mar. Sci. Eng. 2021, 9, 770. [Google Scholar] [CrossRef]
- Yu, D.Y.; Yang, Y.Y.; Li, Y.J. Research on Hydrodynamic Characteristics and Stability of Artificial Reefs with Different Opening Ratios. Period. Ocean Univ. China 2019, 49, 128–136. [Google Scholar]
- Yu, D.Y.; Zhao, W.; Wang, F.Y.; Wang, S.L. Trapezoid artificial reefs in different deployment spacing: Physical and numerical simulations. Oceanol. Limnol. Sin. 2020, 51, 283–292. [Google Scholar]
- Lin, M.J.; Ji, X.R.; Wang, F.X. Flow field effects and hydrodynamic analysis of square composite reefs. Nat. Sci. J. Hainan Univ. 2022, 40, 66–75. [Google Scholar]
- Zhang, Z.; Dong, J.Y.; Sun, X.; Zhang, Y.Y.; Zhan, Q.P.; Zhang, Z.H.; Shen, F.Y.; Ding, X.Y.; Zhang, P.D.; Zhang, X.M. Trophic structure of macrobenthos in artificial reef area of Furong Island, Laizhou Bay. J. Fish. China 2023, 47, 62–75. [Google Scholar]
- Einbinder, S.; Perelberg, A.; Ben-Shaprut, O.; Foucart, M.H.; Shashar, N. Effects of artificial reefs on fish grazing in their vicinity: Evidence from algae presentation experiments. Mar. Environ. 2006, 61, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.K.; Henderson, B.; Spieler, R.E. Fish assemblages associated with artificial reefs of concrete aggregates or quarry stone offshore Miami Beach, Florida, USA. Aquat. Living Resour. 2002, 15, 95–105. [Google Scholar] [CrossRef]
- Zhou, Y.B.; Cai, W.G.; Chen, H.G.; Chen, P.M.; Lv, G.M.; Jia, X.P. Attractive effect of combination of artificial reef models on young Lutjanus argentimaculatus in experimental tank. J. Oceanogr. Taiwan Strait 2012, 31, 231–237. [Google Scholar]
- Chen, H.; Kong, l. Review of Redfish Resources in the Beibu Gulf and Countermeasures for their Conservation. China Fish. 1991, 12, 12–13. [Google Scholar]
- Wang, T.; Shi, J.; Yu, Y.F.; Zhao, J.F.; Xiao, Y.Y.; Liu, Y.; Li, C.H. Progress of research on coral reef fishes in Xisha and recommendations for their protection. Chin. J. Ecol. 2023, 42, 1755–1763. [Google Scholar]
- He, D.R.; Ding, Y. Gathering effect of artificial reef model on garrupa (Epinephelus akaara). J. Oceanogr. Taiwan Strait 1995, 14, 394–398. [Google Scholar]
- He, D.R.; Shi, Y.M. Attractive Effect of Fish Reef Model on Black Porgy (Sparus macrocephalus). J. Xiamen Univ. Nat. Sci. 1995, 34, 653–658. [Google Scholar]
- Zhou, Y.B.; Cai, W.G.; Chen, H.G.; Chen, P.M.; Tang, Z.F.; Lv, G.M.; Jia, X.P. Attraction effect of various artificial reef models on Sebastiscus marmoratus (Cuvier et Valenciennes). Guangdong Agric. Sci. 2011, 38, 8–10+33. [Google Scholar]
- Li, l.; Chen, D.; Peng, J.X.; Zhang, J.M.; Wang, L.; Chen, S.; Quan, W.M.; Jiang, M.; Jiang, Y.Z. On the attractive effects of three artificial reef models for Sparus macrocephlus. Mar. Fish. 2018, 40, 625–631. [Google Scholar]
- Jones, N.A.; Spence, R.; Jones, F.A.; Spence-Jones, H.C. Shade as enrichment: Testing preferences for shelter in two model fish species. J. Fish. Biol. 2019, 95, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, X.D.; Wu, X.Y.; Shi, G.F. Distribution of schlegel’s rockfish (Sebastes schlegeli Hilgendorf) in different artificial reef models. J. Dalian Ocean Univ. 2006, 21, 153–157. [Google Scholar]
- Wu, J.; Zhang, S.; Sun, M.C.; Chen, Y. Experiment on the distribution of different artificial reef models for Paralichthys olivaceus. Mar. Fish. 2004, 26, 271–276. [Google Scholar]
- Shi, J.; Qian, W.G.; Tang, Z.Z.; Han, Z.X.; Zhang, S. Acoustic taming of dusty stingfish Sebastisous marmoratus by 150 Hz rectangular interval sound. J. Dalian Ocean Univ. 2014, 29, 514–519. [Google Scholar]
- Partridge, B.L.; Pitcher, T.J. The sensory basis of fish schools: Relative roles of lateral line and vision. J. Comp. Physiol. 1980, 135, 315–325. [Google Scholar] [CrossRef]
- Hackradt, C.W.; Félix-Hackradt, F.C.; García-Charton, J.A. Influence of habitat structure on fish assemblage of an artificial reef in southern Brazil. Mar. Environ. 2011, 72, 235–247. [Google Scholar] [CrossRef]
- Brochier, T.; Brehmer, P.; Mbaye, A.; Diop, M.; Watanuki, N.; Terashima, H.; Kaplan, D.; Auger, P. Successful artificial reefs depend on getting the context right due to complex socio-bio-economic interactions. Sci. Rep. 2021, 11, 16698. [Google Scholar]
- Reis, B.; van der Linden, P.; Pinto, I.S.; Almada, E.; Borges, M.T.; Hall, A.E.; Stafford, R.; Herbert, R.J.; Lobo-Arteaga, J.; Gaudêncio, M.J. Artificial reefs in the North–East Atlantic area: Present situation, knowledge gaps and future perspectives. Ocean. Coast. Manag. 2021, 213, 105854. [Google Scholar] [CrossRef]
- Vivier, B.; Dauvin, J.-C.; Navon, M.; Rusig, A.-M.; Mussio, I.; Orvain, F.; Boutouil, M.; Claquin, P. Marine artificial reefs, a meta-analysis of their design, objectives and effectiveness. Glob. Ecol. Conserv. 2021, 27, e01538. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Guo, Y.; Qin, C.; Zhang, J.; Zhang, A. Study of the Behavioral Characteristics of Lutjanus erythropterus on the Basis of the Distance and Structure of Artificial Reefs. Biology 2025, 14, 644. https://doi.org/10.3390/biology14060644
Jiang M, Guo Y, Qin C, Zhang J, Zhang A. Study of the Behavioral Characteristics of Lutjanus erythropterus on the Basis of the Distance and Structure of Artificial Reefs. Biology. 2025; 14(6):644. https://doi.org/10.3390/biology14060644
Chicago/Turabian StyleJiang, Manju, Yu Guo, Chuanxin Qin, Jia Zhang, and Ankai Zhang. 2025. "Study of the Behavioral Characteristics of Lutjanus erythropterus on the Basis of the Distance and Structure of Artificial Reefs" Biology 14, no. 6: 644. https://doi.org/10.3390/biology14060644
APA StyleJiang, M., Guo, Y., Qin, C., Zhang, J., & Zhang, A. (2025). Study of the Behavioral Characteristics of Lutjanus erythropterus on the Basis of the Distance and Structure of Artificial Reefs. Biology, 14(6), 644. https://doi.org/10.3390/biology14060644