
Genome-Based Reclassification of Two Haloarcula Species and Characterization of Haloarcula montana sp. nov.


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation of Halophilic Archaea
2.2. Phylogenetic Analysis Based on 16S rRNA and RNA Polymerase B Subunit (rpoB’) Genes
2.3. Genomic Characterization and Comparative Analysis
2.4. Phenotypic Determination
3. Results and Discussion
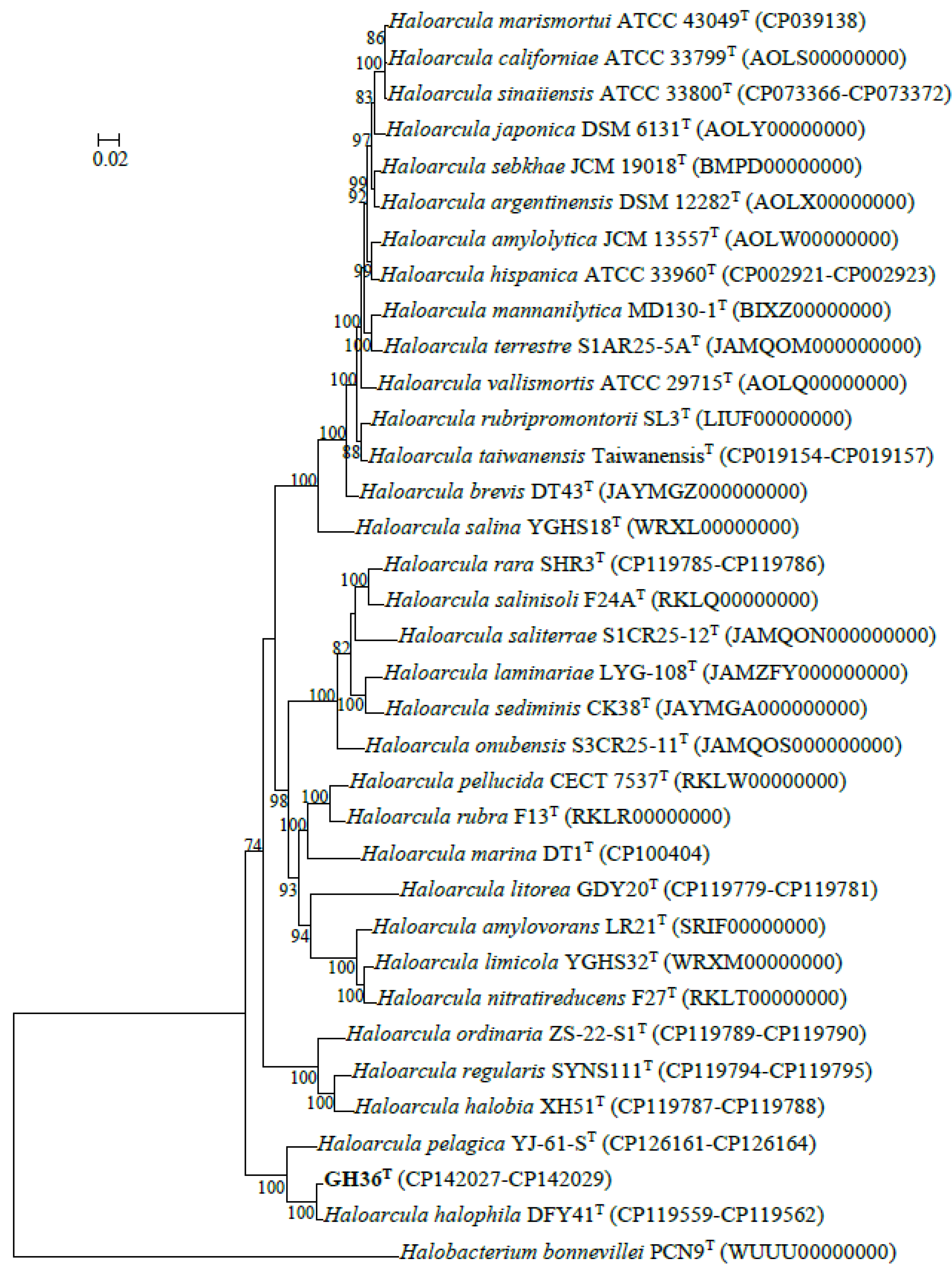
3.1. Phylogenetic Analysis Based on 16S rRNA and rpoB’ Genes
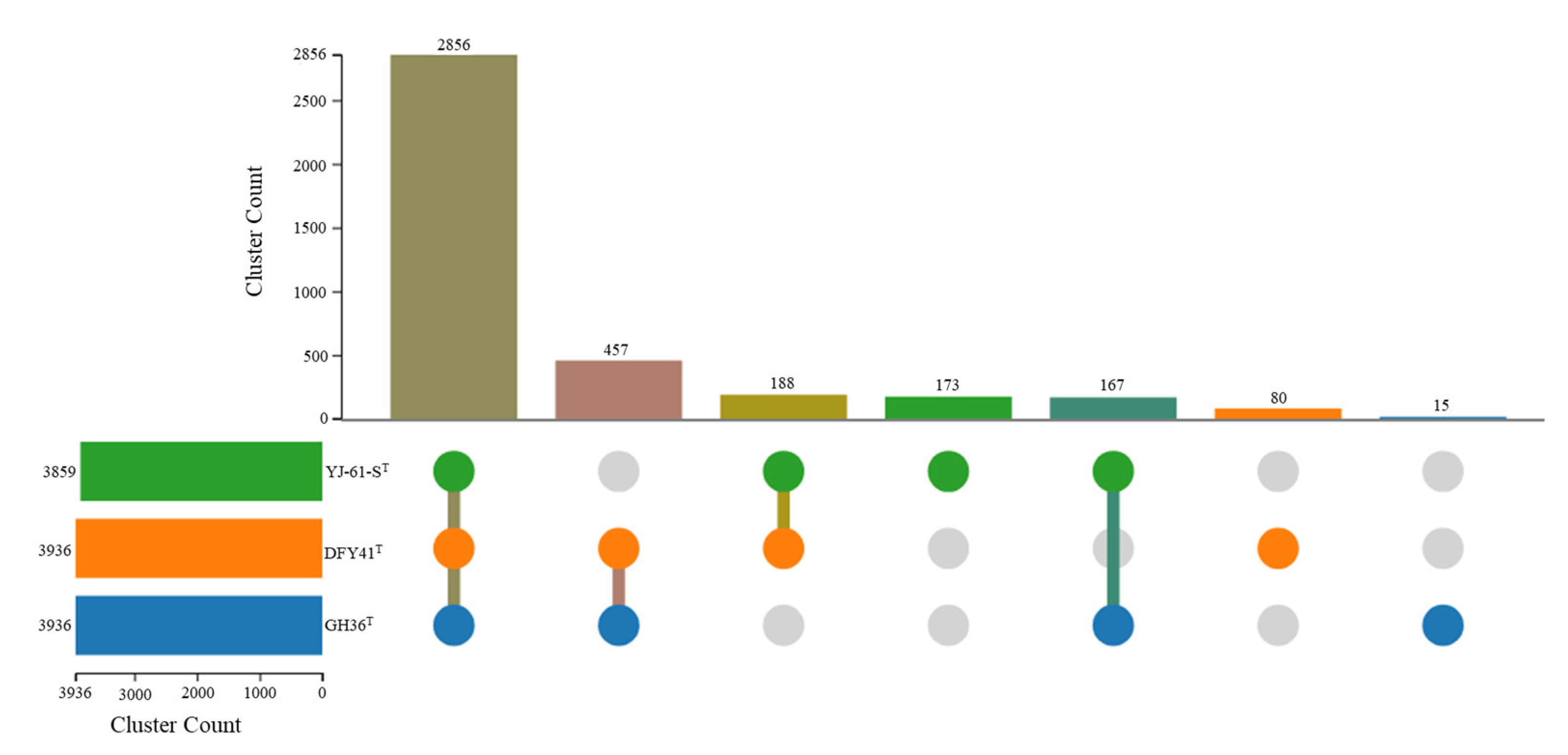
3.2. ORGI and Phylogenomic Analyses
3.3. Genome Annotation and Comparative Genomic Analysis
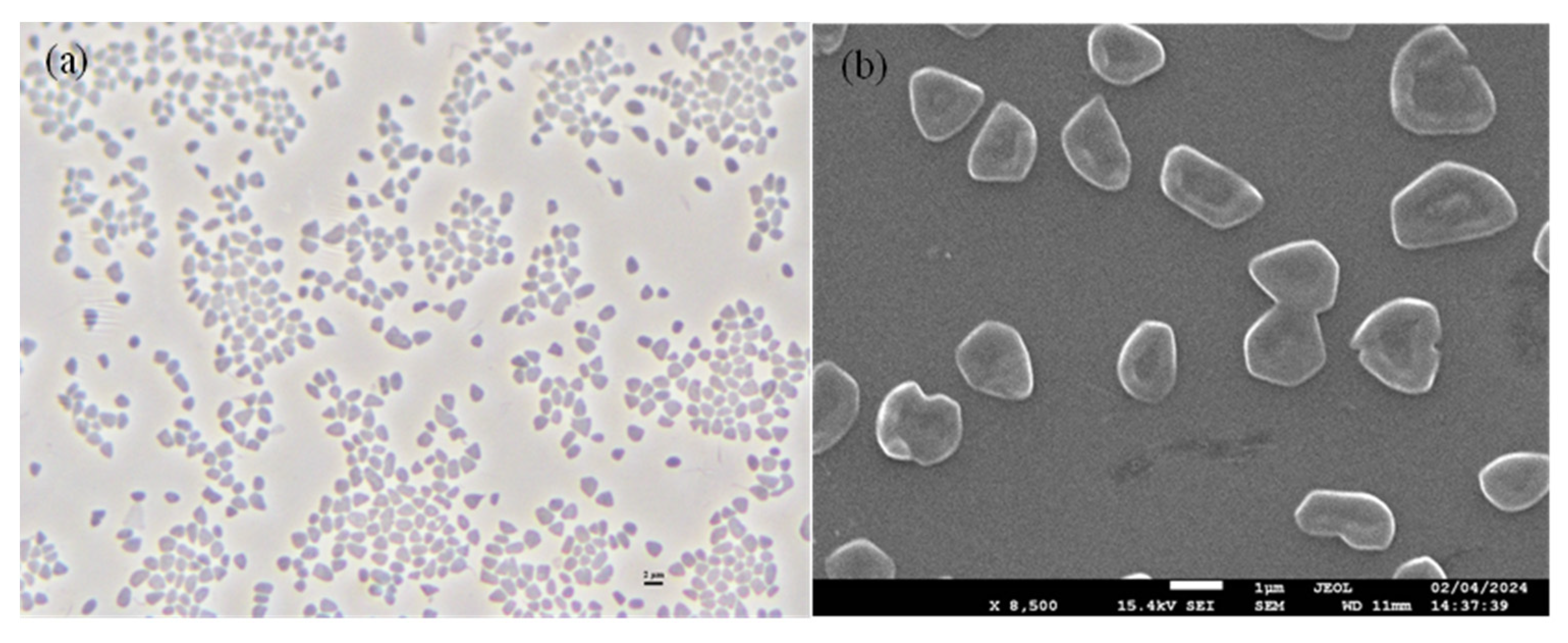
3.4. Phenotypic and Chemotaxonomic Characteristics
4. Taxonomic Conclusions
4.1. Emended Description of Haloarcula marismortui (ex Volcani 1940) Oren et al. 1990
4.2. Description of Haloarcula montana sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.B.; Tang, S.K.; Zhang, Y.G.; Lu, X.H.; Li, L.; Cheng, J.; Zhang, Y.M.; Zhang, L.L.; Li, W.J. Halalkalicoccus paucihalophilus sp. nov., a halophilic archaeon from Lop Nur region in Xinjiang, northwest of China. Antonie Van Leeuwenhoek 2013, 103, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Litchfield, C.D. Potential for industrial products from the halophilic Archaea. J. Ind. Microbiol. Biotechnol. 2011, 38, 1635–1647. [Google Scholar] [CrossRef]
- Oren, A. Diversity of halophilic microorganisms: Environments, phylogeny, physiology, and applications. J. Ind. Microbiol. Biotechnol. 2002, 28, 56–63. [Google Scholar] [CrossRef]
- Wei, W.; Hu, X.; Yang, S.; Wang, K.; Zeng, C.; Hou, Z.; Cui, H.; Liu, S.; Zhu, L. Denitrifying halophilic archaea derived from salt dominate the degradation of nitrite in salted radish during pickling. Food Res. Int. 2022, 152, 110906. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Li, X.X.; Sun, Y.; Li, Y.; Yang, X.Y.; Sun, Y.P.; Cui, H.L. Novel archaeal histamine oxidase from Natronobeatus ordinarius: Insights into histamine degradation for enhancing food safety. J. Agric. Food Chem. 2024, 72, 6519–6525. [Google Scholar] [CrossRef]
- Hou, J.; Zhang, Q.K.; Zhang, R.Y.; Li, S.Y.; Liu, Y.Y.; Cui, H.L. A hyperstable, low-salt adapted protease from halophilic archaeon with potential applications in salt-fermented foods. Food Res. Int. 2024, 191, 114738. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, A.K. Haloarchaea: Worth exploring for their biotechnological potential. Biotechnol. Lett. 2017, 39, 1793–1800. [Google Scholar] [CrossRef]
- Ma, X.; Hu, Y.; Li, X.X.; Tan, S.; Cheng, M.; Hou, J.; Cui, H.L. Genome-based taxonomy of genera Haloarcula and Halomicroarcula, and description of six novel species of Haloarcula. Extremophiles 2024, 28, 10. [Google Scholar] [CrossRef]
- Straková, D.; Sánchez-Porro, C.; de la Haba, R.R.; Ventosa, A. Reclassification of Halomicroarcula saliterrae Straková et al. 2024 and Halomicroarcula onubensis Straková et al. 2024 into the genus Haloarcula, as Haloarcula saliterrae comb. nov. and Haloarcula onubensis comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2024, 74, 006510. [Google Scholar] [CrossRef]
- Torreblanca, M.; Rodriguez-Valera, F.; Juez, G.; Ventosa, A.; Kamekura, M.; Kates, M. Classification of non-alkaliphilic halobacteria based on numerical taxonomy and polar lipid composition, and description of Haloarcula gen. nov. and Haloferax gen. nov. Syst. Appl. Microbiol. 1986, 8, 89–99. [Google Scholar] [CrossRef]
- Enomoto, S.; Shimane, Y.; Ihara, K.; Kamekura, M.; Itoh, T.; Ohkuma, M.; Takahashi-Ando, N.; Fukushima, Y.; Yoshida, Y.; Usami, R.; et al. Haloarcula mannanilytica sp. nov., a galactomannan-degrading haloarchaeon isolated from commercial salt. Int. J. Syst. Evol. Microbiol. 2020, 70, 6331–6337. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Hu, Y.; Hou, J.; Zhou, Y.G.; Cui, H.L. Genome-based classification of ‘Haloarcula aidinensis’ and description of three novel halophilic archaea isolated from an inland saline lake, coastal saline soil, and a marine solar saltern. Curr. Microbiol. 2024, 81, 442. [Google Scholar] [CrossRef]
- Straková, D.; Galisteo, C.; de la Haba, R.R.; Ventosa, A. Characterization of Haloarcula terrestris sp. nov. and reclassification of a Haloarcula species based on a taxogenomic approach. Int. J. Syst. Evol. Microbiol. 2023, 73, 006157. [Google Scholar] [CrossRef]
- Han, D.; Cui, H.L. Halostella pelagica sp. nov. and Halostella litorea sp. nov., isolated from salted brown alga Laminaria. Int. J. Syst. Evol. Microbiol. 2020, 70, 1969–1976. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ma, X.; Li, X.X.; Tan, S.; Cheng, M.; Hou, J.; Cui, H.L. Halomicrococcus gelatinilyticus sp. nov. and Halosimplex aquaticum sp. nov., halophilic archaea isolated from saline soil and an inland solar saltern. Int. J. Syst. Evol. Microbiol. 2024, 74, 006231. [Google Scholar] [CrossRef]
- Cui, H.L.; Zhou, P.J.; Oren, A.; Liu, S.J. Intraspecific polymorphism of 16S rRNA genes in two halophilic archaeal genera, Haloarcula and Halomicrobium. Extremophiles 2009, 13, 31–37. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Rodriguez, R.L.; Konstantinidis, K.T. MyTaxa: An advanced taxonomic classifier for genomic and metagenomic sequences. Nucleic Acids Res. 2014, 42, e73. [Google Scholar] [CrossRef]
- Cui, C.; Han, D.; Hou, J.; Cui, H.L. Genome-based classification of the class Halobacteria and description of Haladaptataceae fam. nov. and Halorubellaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2023, 73, 005984. [Google Scholar] [CrossRef]
- Rinke, C.; Chuvochina, M.; Mussig, A.J.; Chaumeil, P.A.; Davín, A.A.; Waite, D.W.; Whitman, W.B.; Parks, D.H.; Hugenholtz, P. A standardized archaeal taxonomy for the Genome Taxonomy Database. Nat. Microbiol. 2021, 6, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, Y.; Fan, G.; Sun, D.; Zhang, X.; Yu, Z.; Wang, J.; Wu, L.; Shi, W.; Ma, J. IPGA: A handy integrated prokaryotes genome and pan-genome analysis web service. iMeta 2022, 1, e55. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Cui, H.L.; Hou, J.; Amoozegar, M.A.; Dyall-Smith, M.L.; de la Haba, R.R.; Minegishi, H.; Montalvo-Rodriguez, R.; Oren, A.; Sanchez-Porro, C.; Ventosa, A.; et al. Proposed minimal standards for description of new taxa of the class Halobacteria. Int. J. Syst. Evol. Microbiol. 2024, 74, 006290. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Zhu, L.R.; Zhang, Q.K.; Dong, X.Y.; Hou, J.; Cui, H.L. Genome-based classification of the family Haloferacaceae and description of five novel species of Halobaculum. Extremophiles 2024, 28, 22. [Google Scholar] [CrossRef]
- Dussault, H.P. An improved technique for staining red halophilic bacteria. J. Bacteriol. 1955, 70, 484–485. [Google Scholar] [CrossRef]
- Wang, B.B.; Bao, C.X.; Sun, Y.P.; Hou, J.; Cui, H.L. Halobacterium wangiae sp. nov. and Halobacterium zhouii sp. nov., two extremely halophilic archaea isolated from sediment of a salt lake and saline soil of an inland saltern. Int. J. Syst. Evol. Microbiol. 2023, 73, 005922. [Google Scholar] [CrossRef]
- Tao, C.Q.; Ding, Y.; Zhao, Y.J.; Cui, H.L. Natronorubrum halophilum sp. nov. isolated from two inland salt lakes. J. Microbiol. 2020, 58, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.L.; Gao, X.; Yang, X.; Xu, X.W. Halorussus rarus gen. nov., sp. nov., a new member of the family Halobacteriaceae isolated from a marine solar saltern. Extremophiles 2010, 14, 493–499. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Oren, A.; Ginzburg, M.; Ginzburg, B.Z.; Hochstein, L.I.; Volcani, B.E. Haloarcula marismortui (Volcani) sp. nov., nom. rev., an extremely halophilic bacterium from the Dead Sea. Int. J. Syst. Bacteriol. 1990, 40, 209–210. [Google Scholar] [CrossRef]
- Javor, B.; Requadt, C.; Stoeckenius, W. Box-shaped halophilic bacteria. J. Bacteriol. 1982, 151, 1532–1542. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Optimum NaCl (M) | 3.1 | 2.6 | 4.8 | 3.4 | 3.1 | 3.4 | 4.8 | 2.6 | 2.6 | 3.4 |
| Temperature optimum (℃) | 37 | 42 | 40 | 37 | 41 | 35 | 37 | 40 | 37 | 35 |
| Optimum pH | 7.5 | 7.0 | 7.0 | 7.0 | 7.5 | 7.0 | 7.0 | 7.0 | 7.0 | 7.0 |
| Anaerobic growth with nitrate | + | + | + | + | + | − | − | − | − | − |
| Anaerobic growth with arginine | + | + | − | − | − | − | − | − | − | − |
| Anaerobic growth with DMSO | − | + | − | + | − | − | − | − | − | + |
| Utilization of: | ||||||||||
| d-Mannose | + | + | + | + | + | − | + | + | + | − |
| d-Galactose | + | + | + | + | + | + | + | + | − | + |
| l-Sorbose | + | − | − | − | − | − | − | − | − | − |
| Maltose | − | + | + | − | + | − | − | − | − | − |
| Sucrose | − | + | + | + | + | + | + | + | + | + |
| Lactose | − | + | + | − | − | − | − | − | − | − |
| Indole formation | − | − | − | − | + | − | − | − | − | − |
| Starch hydrolysis | − | + | − | − | + | − | − | − | − | + |
| Gelatin hydrolysis | − | − | − | − | − | − | − | − | − | − |
| Tween 80 hydrolysis | − | − | − | − | − | − | − | − | − | + |
| H2S formation | − | + | − | − | + | − | − | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.-Q.; Zhu, L.-R.; Mao, Y.-L.; Ma, X.; Hou, J. Genome-Based Reclassification of Two Haloarcula Species and Characterization of Haloarcula montana sp. nov. Biology 2025, 14, 615. https://doi.org/10.3390/biology14060615
Liu J-Q, Zhu L-R, Mao Y-L, Ma X, Hou J. Genome-Based Reclassification of Two Haloarcula Species and Characterization of Haloarcula montana sp. nov. Biology. 2025; 14(6):615. https://doi.org/10.3390/biology14060615
Chicago/Turabian StyleLiu, Ji-Qiang, Ling-Rui Zhu, Ya-Ling Mao, Xue Ma, and Jing Hou. 2025. "Genome-Based Reclassification of Two Haloarcula Species and Characterization of Haloarcula montana sp. nov." Biology 14, no. 6: 615. https://doi.org/10.3390/biology14060615
APA StyleLiu, J.-Q., Zhu, L.-R., Mao, Y.-L., Ma, X., & Hou, J. (2025). Genome-Based Reclassification of Two Haloarcula Species and Characterization of Haloarcula montana sp. nov. Biology, 14(6), 615. https://doi.org/10.3390/biology14060615