
Glycosylation and Acylation: Important Regulators of Immune Cell Fate Decisions


Simple Summary
Abstract
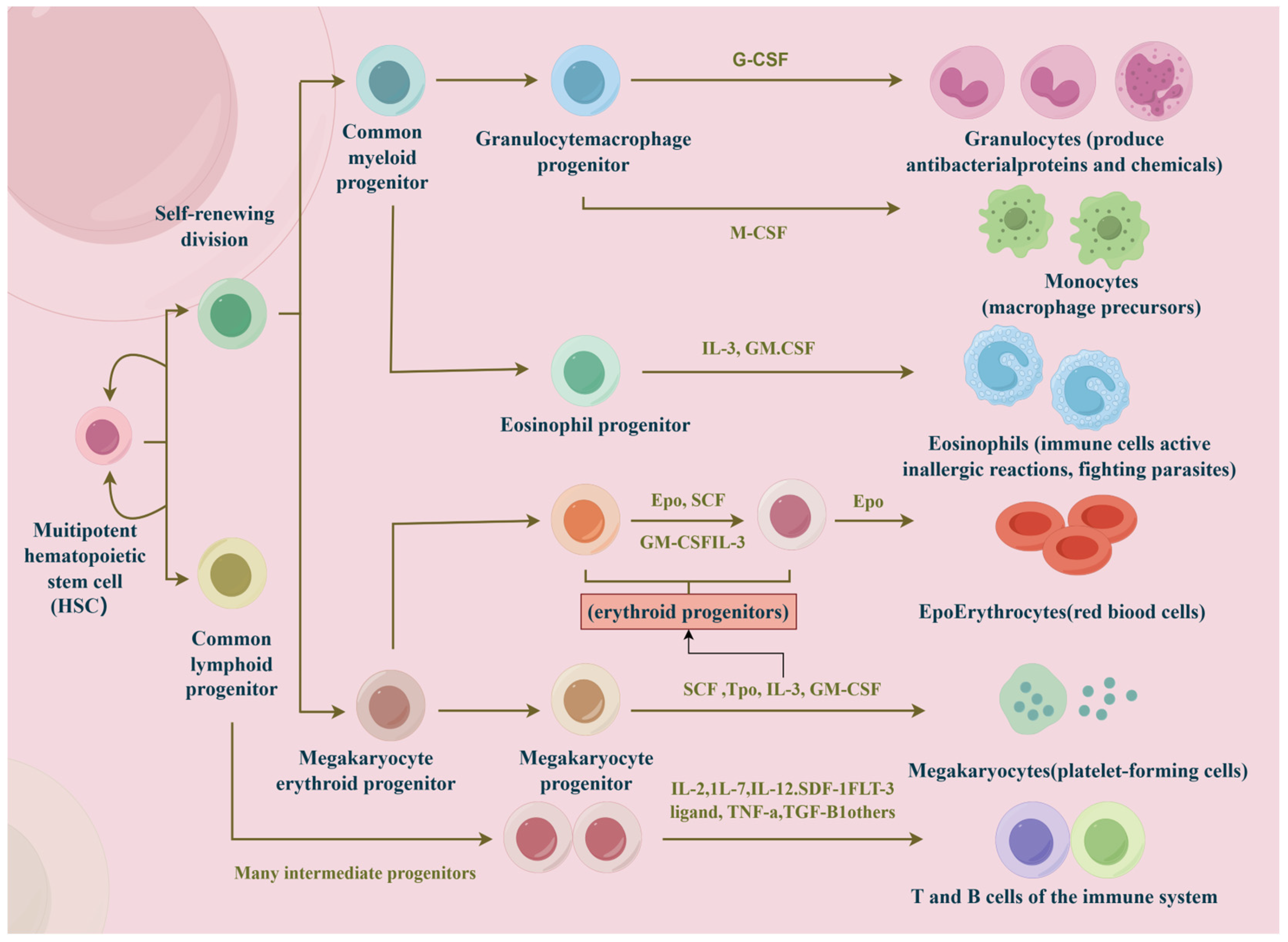
1. Introduction
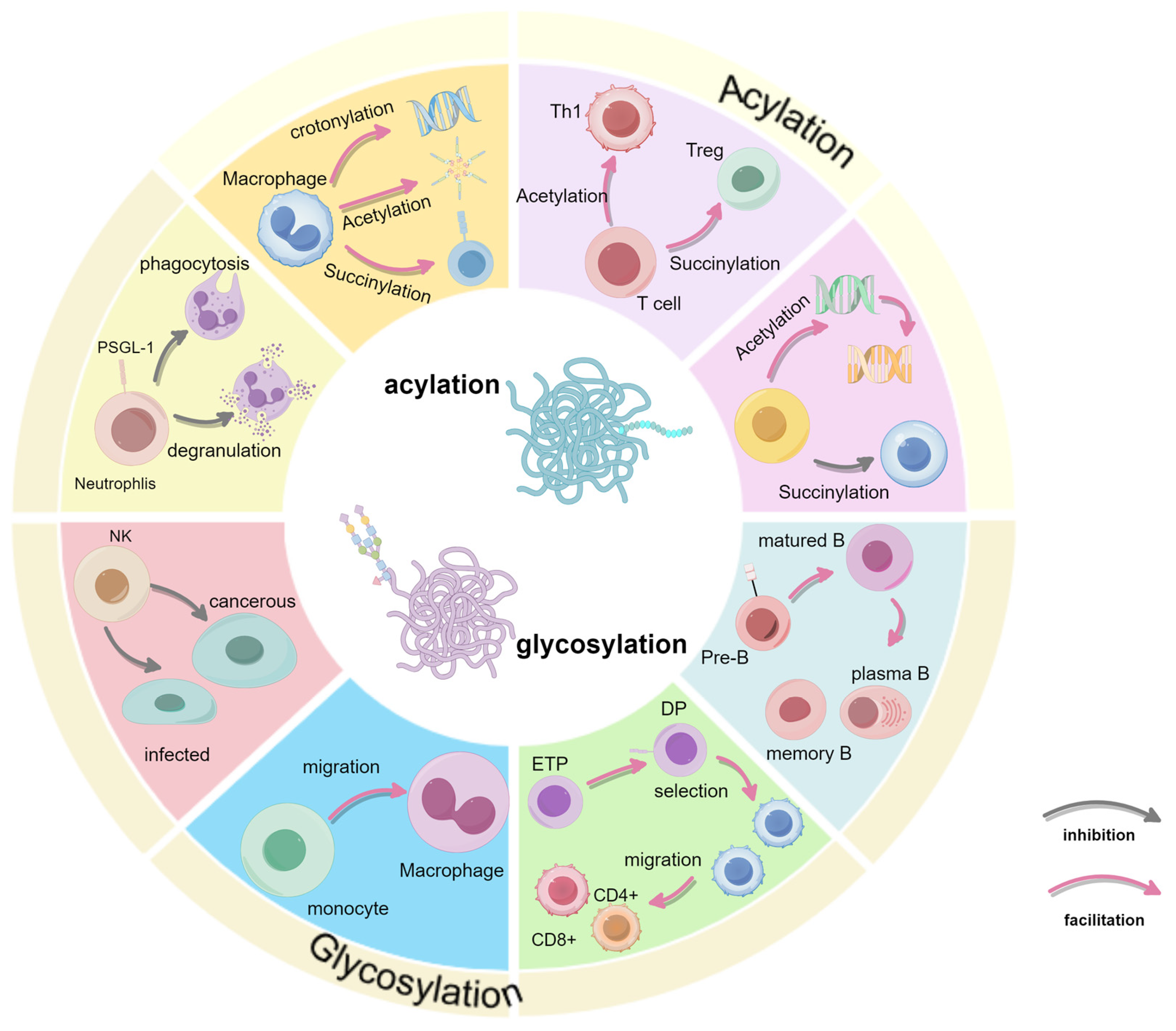
2. Regulation of Glycosylation in Immune Cells
2.1. Innate Immune Cells
2.1.1. Neutrophils
2.1.2. Natural Killer (NK) Cells
2.1.3. Macrophage
2.2. Acquired Immune Cells
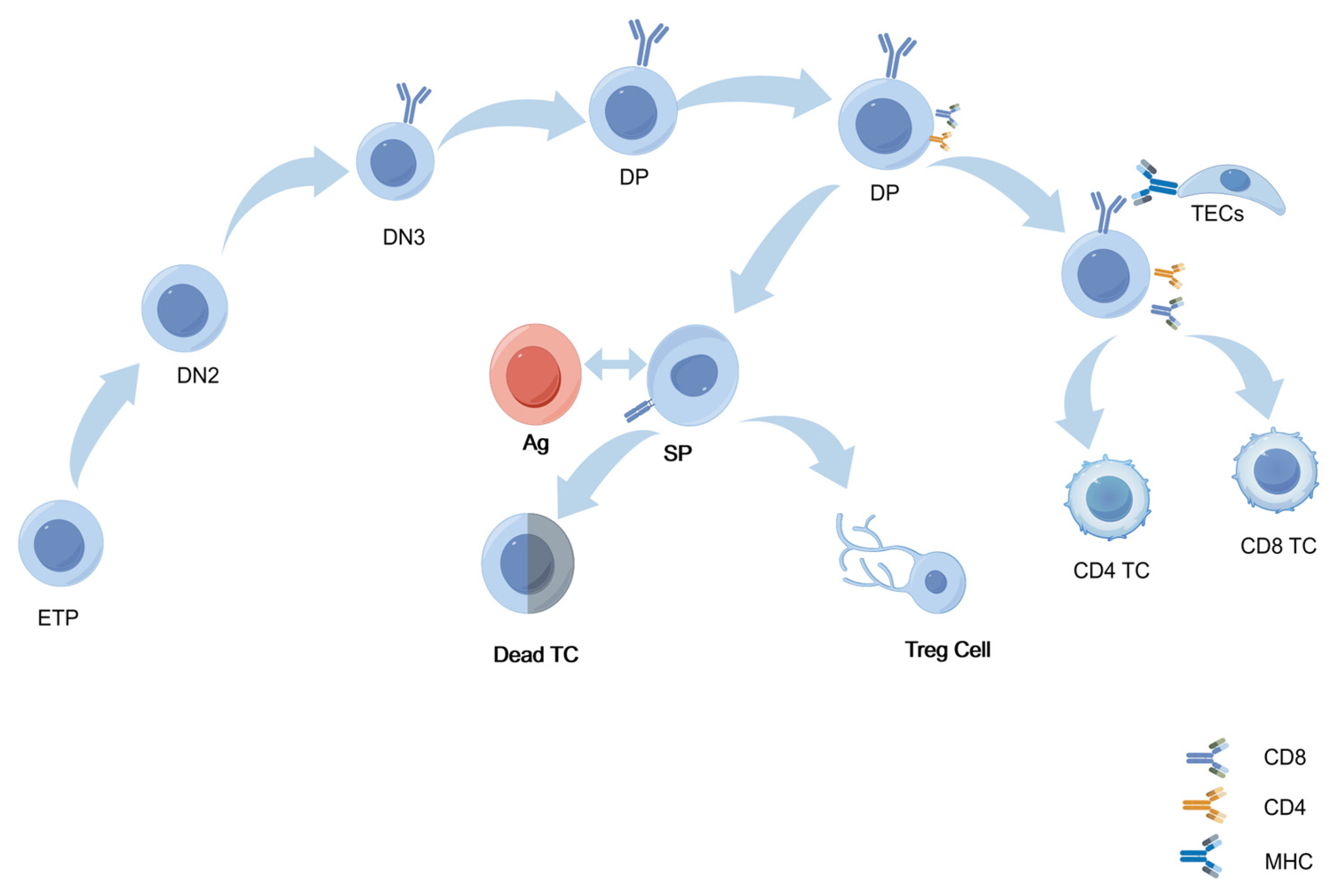
2.2.1. T Cells
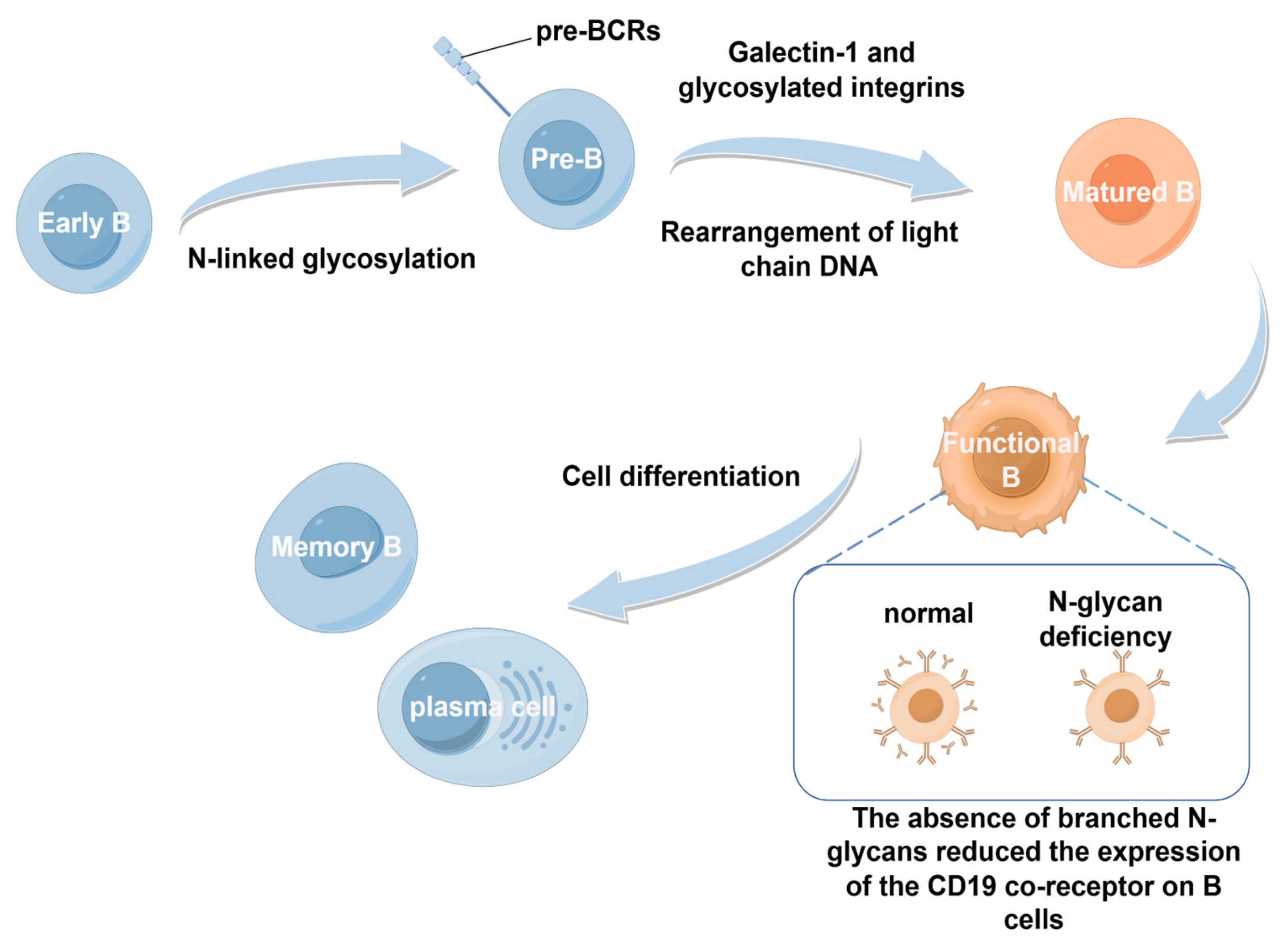
2.2.2. B Cells
2.3. Clinical Studies on Glycosylation
3. Regulation of Acylation in Immune Cells
3.1. Macrophage
3.2. Acquired Immune Cells
3.2.1. T Cells
3.2.2. B Cells
3.3. Clinical Studies on Acylation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Riera Romo, M.; Pérez-Martínez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Smotrys, J.E.; Linder, M.E. Palmitoylation of intracellular signaling proteins: Regulation and function. Annu. Rev. Biochem. 2004, 73, 559–587. [Google Scholar] [CrossRef] [PubMed]
- von Boehmer, H. Unique features of the pre-T-cell receptor alpha-chain: Not just a surrogate. Nat. Rev. Immunol. 2005, 5, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Gan, S.; Zhu, Q.; Dai, D.; Li, N.; Wang, H.; Chen, X.; Hou, D.; Wang, Y.; Pan, Q.; et al. Modulation of M2 macrophage polarization by the crosstalk between Stat6 and Trim24. Nat. Commun. 2019, 10, 4353. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, Y.; Zhang, R.; Hei, H. Advances in post-translational modifications of proteins and cancer immunotherapy. Front. Immunol. 2023, 14, 1229397. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, Y.; Wang, P.; Xu, J. Phosphorylation: A Fast Switch for Checkpoint Signaling. Adv. Exp. Med. Biol. 2020, 1248, 347–398. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Sun, L.; Li, C.-F.; Wang, Y.-H.; Yao, J.; Li, H.; Yan, M.; Chang, W.-C.; Hsu, J.-M.; Cha, J.-H.; et al. Galectin-9 interacts with PD-1 and TIM-3 to regulate T cell death and is a target for cancer immunotherapy. Nat. Commun. 2021, 12, 832. [Google Scholar] [CrossRef]
- Xu, B.; Yuan, L.; Gao, Q.; Yuan, P.; Zhao, P.; Yuan, H.; Fan, H.; Li, T.; Qin, P.; Han, L.; et al. Circulating and tumor-infiltrating Tim-3 in patients with colorectal cancer. Oncotarget 2015, 6, 20592–20603. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Eichler, J. Protein glycosylation. Curr. Biol. 2019, 29, R229–R231. [Google Scholar] [CrossRef] [PubMed]
- Mulagapati, S.; Koppolu, V.; Raju, T.S. Decoding of O-Linked Glycosylation by Mass Spectrometry. Biochemistry 2017, 56, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Ugonotti, J.; Chatterjee, S.; Thaysen-Andersen, M. Structural and functional diversity of neutrophil glycosylation in innate immunity and related disorders. Mol. Asp. Med. 2021, 79, 100882. [Google Scholar] [CrossRef] [PubMed]
- Prame Kumar, K.; Nicholls, A.J.; Wong, C.H.Y. Partners in crime: Neutrophils and monocytes/macrophages in inflammation and disease. Cell Tissue Res. 2018, 371, 551–565. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chacko, B.K.; Scott, D.W.; Chandler, R.T.; Patel, R.P. Endothelial surface N-glycans mediate monocyte adhesion and are targets for anti-inflammatory effects of peroxisome proliferator-activated receptor γ ligands. J. Biol. Chem. 2011, 286, 38738–38747. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Huang, N.; Fan, X.; Zaleta-Rivera, K.; Nguyen, T.C.; Zhou, J.; Luo, Y.; Gao, J.; Fang, R.H.; Yan, Z.; Chen, Z.B.; et al. Natural display of nuclear-encoded RNA on the cell surface and its impact on cell interaction. Genome Biol. 2020, 21, 225. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, N.; Tang, W.; Torres, L.; Wang, X.; Ajaj, Y.; Zhu, L.; Luan, Y.; Zhou, H.; Wang, Y.; Zhang, D.; et al. Cell surface RNAs control neutrophil recruitment. Cell 2024, 187, 846–860.e17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yamaji, T.; Teranishi, T.; Alphey, M.S.; Crocker, P.R.; Hashimoto, Y. A small region of the natural killer cell receptor, Siglec-7, is responsible for its preferred binding to alpha 2,8-disialyl and branched alpha 2,6-sialyl residues. A comparison with Siglec-9. J. Biol. Chem. 2002, 277, 6324–6332. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Gonzalez-Gil, A.; Wei, Y.; Fernandes, S.M.; Porell, R.N.; Vajn, K.; Paulson, J.C.; Nycholat, C.M.; Schnaar, R.L. Siglec-8 and Siglec-9 binding specificities and endogenous airway ligand distributions and properties. Glycobiology 2017, 27, 657–668. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Patel, K.R.; Rodriguez Benavente, M.C.; Lorenz, W.W.; Mace, E.M.; Barb, A.W. Fc γ receptor IIIa/CD16a processing correlates with the expression of glycan-related genes in human natural killer cells. J. Biol. Chem. 2021, 296, 100183. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hume, D.A.; Irvine, K.M.; Pridans, C. The Mononuclear Phagocyte System: The Relationship Between Monocytes and Macrophages. Trends Immunol. 2019, 40, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ma, C.; Lu, Y.; Wang, J.; Yun, H.; Jiang, H.; Wu, M.; Feng, X.; Gai, W.; Xu, G.; et al. Rack1 regulates B-cell development and function by binding to and stabilizing the transcription factor Pax5. Cell Mol. Immunol. 2024, 21, 1282–1295. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Swamy, M.; Pathak, S.; Grzes, K.M.; Damerow, S.; Sinclair, L.V.; van Aalten, D.M.; Cantrell, D.A. Glucose and glutamine fuel protein O-GlcNAcylation to control T cell self-renewal and malignancy. Nat. Immunol. 2016, 17, 712–720. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vicente, M.M.; Alves, I.; Fernandes, Â.; Dias, A.M.; Santos-Pereira, B.; Pérez-Anton, E.; Santos, S.; Yang, T.; Correia, A.; Münster-Kühnel, A.; et al. Mannosylated glycans impair normal T-cell development by reprogramming commitment and repertoire diversity. Cell Mol. Immunol. 2023, 20, 955–968. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vicente, M.M.; Leite-Gomes, E.; Pinho, S.S. Glycome dynamics in T and B cell development: Basic immunological mechanisms and clinical applications. Trends Immunol. 2023, 44, 585–597. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bacigalupa, Z.A.; Landis, M.D.; Rathmell, J.C. Nutrient inputs and social metabolic control of T cell fate. Cell Metab. 2024, 36, 10–20. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, C.W.; Chung, E.M.; Yang, R.; Kim, Y.S.; Park, A.H.; Lai, Y.J.; Yang, Y.; Wang, Y.H.; Liu, J.; et al. Targeting Glycosylated PD-1 Induces Potent Antitumor Immunity. Cancer Res. 2020, 80, 2298–2310. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sun, Y.; Li, X.; Wang, T.; Li, W. Core Fucosylation Regulates the Function of Pre-BCR, BCR and IgG in Humoral Immunity. Front. Immunol. 2022, 13, 844427. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Radovani, B.; Gudelj, I. N-Glycosylation and Inflammation; the Not-So-Sweet Relation. Front. Immunol. 2022, 13, 893365. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mortales, C.L.; Lee, S.U.; Demetriou, M. N-Glycan Branching Is Required for Development of Mature B Cells. J. Immunol. 2020, 205, 630–636. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Brown, K.A.; Brain, S.D.; Pearson, J.D.; Edgeworth, J.D.; Lewis, S.M.; Treacher, D.F. Neutrophils in development of multiple organ failure in sepsis. Lancet 2006, 368, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Lizcano, A.; Secundino, I.; Döhrmann, S.; Corriden, R.; Rohena, C.; Diaz, S.; Ghosh, P.; Deng, L.; Nizet, V.; Varki, A. Erythrocyte sialoglycoproteins engage Siglec-9 on neutrophils to suppress activation. Blood 2017, 129, 3100–3110. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Deniset, J.F.; Kubes, P. Recent advances in understanding neutrophils. F1000Res 2016, 5, 2912. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chiewchengchol, D.; Midgley, A.; Sodsai, P.; Deekajorndech, T.; Hirankarn, N.; Beresford, M.W.; Edwards, S.W. The protective effect of GM-CSF on serum-induced neutrophil apoptosis in juvenile systemic lupus erythematosus patients. Clin. Rheumatol. 2015, 34, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Yang, Q.; Li, F.; Zhu, M.; Yang, H.; Tan, T.; Wu, B.; Liu, M.; Xu, C.; Yin, J.; et al. The Glycosylation of Immune Checkpoints and Their Applications in Oncology. Pharmaceuticals 2022, 15, 1451. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rosenstock, P.; Kaufmann, T. Sialic Acids and Their Influence on Human NK Cell Function. Cells 2021, 10, 263. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pereira, M.S.; Alves, I.; Vicente, M.; Campar, A.; Silva, M.C.; Padrão, N.A.; Pinto, V.; Fernandes, Â.; Dias, A.M.; Pinho, S.S. Glycans as Key Checkpoints of T Cell Activity and Function. Front. Immunol. 2018, 9, 2754. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ballesteros-Tato, A.; León, B.; Graf, B.A.; Moquin, A.; Adams, P.S.; Lund, F.E.; Randall, T.D. Interleukin-2 inhibits germinal center formation by limiting T follicular helper cell differentiation. Immunity 2012, 36, 847–856. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Matsuura, A.; Ito, M.; Sakaidani, Y.; Kondo, T.; Murakami, K.; Furukawa, K.; Nadano, D.; Matsuda, T.; Okajima, T. O-linked N-acetylglucosamine is present on the extracellular domain of notch receptors. J. Biol. Chem. 2008, 283, 35486–35495. [Google Scholar] [CrossRef] [PubMed]
- Takaba, H.; Takayanagi, H. The Mechanisms of T Cell Selection in the Thymus. Trends Immunol. 2017, 38, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Rudd, P.M.; Elliott, T.; Cresswell, P.; Wilson, I.A.; Dwek, R.A. Glycosylation and the immune system. Science 2001, 291, 2370–2376. [Google Scholar] [CrossRef] [PubMed]
- Werlen, G.; Li, M.L.; Tottone, L.; da Silva-Diz, V.; Su, X.; Herranz, D.; Jacinto, E. Dietary glucosamine overcomes the defects in αβ-T cell ontogeny caused by the loss of de novo hexosamine biosynthesis. Nat. Commun. 2022, 13, 7404. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dong, W.; Lin, M.; Zhang, R.; Sun, X.; Li, H.; Liu, T.; Xu, Y.; Lv, L. D-mannose targets PD-1 to lysosomal degradation and enhances T cell-mediated anti-tumor immunity. Cancer Lett. 2024, 591, 216883. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, T.; Xu, J. Therapeutic Development of Immune Checkpoint Inhibitors. Adv. Exp. Med. Biol. 2020, 1248, 619–649. [Google Scholar] [CrossRef] [PubMed]
- Mkhikian, H.; Grigorian, A.; Li, C.F.; Chen, H.L.; Newton, B.; Zhou, R.W.; Beeton, C.; Torossian, S.; Tatarian, G.G.; Lee, S.U.; et al. Genetics and the environment converge to dysregulate N-glycosylation in multiple sclerosis. Nat. Commun. 2011, 2, 334. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Scherer, H.U.; van der Woude, D.; Toes, R.E.M. From risk to chronicity: Evolution of autoreactive B cell and antibody responses in rheumatoid arthritis. Nat. Rev. Rheumatol. 2022, 18, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Mallory, T.; Satomura, S. AFP-L3: A new generation of tumor marker for hepatocellular carcinoma. Clin. Chim. Acta 2001, 313, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-T.; Zhang, C.; Wu, J.; Lu, P.; Xu, L.; Yuan, H.; Feng, Y.; Chen, Z.-S.; Wang, N. Biomarkers for diagnosis and therapeutic options in hepatocellular carcinoma. Mol. Cancer 2024, 23, 189. [Google Scholar] [CrossRef]
- Ibrahim, N.K.; Murray, J.L. Clinical Development of the STn-KLH Vaccine (Theratope®). Clin. Breast Cancer 2003, 3, S139–S143. [Google Scholar] [CrossRef]
- Temesfői, V.; Molnár, K.; Kaltenecker, P.; Réger, B.; Szomor, Á.; Horváth-Szalai, Z.; Alizadeh, H.; Kajtár, B.; Kőszegi, T.; Miseta, A.; et al. O-GlcNAcylation in early stages of chronic lymphocytic leukemia: Protocol development for flow cytometry. Cancer Biomark. 2021, 32, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Natoni, A.; Cerreto, M.; De Propris, M.S.; Del Giudice, I.; Soscia, R.; Peragine, N.; Intoppa, S.; Milani, M.L.; Guarini, A.; Foà, R. Sialylation regulates migration in chronic lymphocytic leukemia. Haematologica 2023, 108, 1851–1860. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shang, S.; Liu, J.; Hua, F. Protein acylation: Mechanisms, biological functions and therapeutic targets. Signal Transduct. Target. Ther. 2022, 7, 396. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tan, M.; Luo, H.; Lee, S.; Jin, F.; Yang, J.S.; Montellier, E.; Buchou, T.; Cheng, Z.; Rousseaux, S.; Rajagopal, N.; et al. Identification of 67 histone marks and histone lysine crotonylation as a new type of histone modification. Cell 2011, 146, 1016–1028. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, Y.; Luo, L.; Xu, X.; Wu, J.; Wang, F.; Lu, Y.; Zhang, N.; Ding, Y.; Lu, B.; Zhao, K. Acetylation is required for full activation of the NLRP3 inflammasome. Nat. Commun. 2023, 14, 8396. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Le Cam, L.; Linares, L.K.; Paul, C.; Julien, E.; Lacroix, M.; Hatchi, E.; Triboulet, R.; Bossis, G.; Shmueli, A.; Rodriguez, M.S.; et al. E4F1 Is an Atypical Ubiquitin Ligase that Modulates p53 Effector Functions Independently of Degradation. Cell 2006, 127, 775–788. [Google Scholar] [CrossRef]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, L.; Xiang, T.; Guo, J.; Guo, F.; Wu, Y.; Feng, H.; Liu, J.; Tao, S.; Fu, P.; Ma, L. Inhibition of ACSS2-mediated histone crotonylation alleviates kidney fibrosis via IL-1β-dependent macrophage activation and tubular cell senescence. Nat. Commun. 2024, 15, 3200. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yu, H.; Bu, C.; Liu, Y.; Gong, T.; Liu, X.; Liu, S.; Peng, X.; Zhang, W.; Peng, Y.; Yang, J.; et al. Global crotonylome reveals CDYL-regulated RPA1 crotonylation in homologous recombination-mediated DNA repair. Sci. Adv. 2020, 6, eaay4697. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, Y.; Sabari, B.R.; Panchenko, T.; Wen, H.; Zhao, D.; Guan, H.; Wan, L.; Huang, H.; Tang, Z.; Zhao, Y.; et al. Molecular Coupling of Histone Crotonylation and Active Transcription by AF9 YEATS Domain. Mol. Cell 2016, 62, 181–193. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jing, X.; Lyu, J.; Xiong, J. Acetate regulates GAPDH acetylation and T helper 1 cell differentiation. Mol. Biol. Cell 2023, 34, br10. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Weinert, B.T.; Schölz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.L. The complex language of chromatin regulation during transcription. Nature 2007, 447, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.; Tora, L. Distinct GCN5/PCAF-containing complexes function as co-activators and are involved in transcription factor and global histone acetylation. Oncogene 2007, 26, 5341–5357. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Nakayama, M.; Kawai, C.; Kuribayashi, F.; Mimuro, H.; Imajoh-Ohmi, S.; Nishitoh, H.; Takami, Y.; Nakayama, T. Histone acetyltransferase p300/CBP-associated factor is an effective suppressor of secretory immunoglobulin synthesis in immature B cells. Microbiol. Immunol. 2015, 59, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Gamez-Garcia, A.; Espinosa-Alcantud, M.; Bueno-Costa, A.; Alari-Pahissa, E.; Marazuela-Duque, A.; Thackray, J.K.; Ray, C.; Berenguer, C.; Kumari, P.; Bech, J.J.; et al. A SIRT7-dependent acetylation switch regulates early B cell differentiation and lineage commitment through Pax5. Nat. Immunol. 2024, 25, 2308–2319. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xu, Y.; Harder, K.W.; Huntington, N.D.; Hibbs, M.L.; Tarlinton, D.M. Lyn tyrosine kinase: Accentuating the positive and the negative. Immunity 2005, 22, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Liu, Y.; Yan, J.; Zhao, L.; Zhou, Y.; Shen, X.; Chen, Y.; Chen, Y.; Meng, X.; Zhang, X.; et al. Fumarate suppresses B-cell activation and function through direct inactivation of LYN. Nat. Chem. Biol. 2022, 18, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, Y.; Hu, Y.; Li, Q.; Lan, Y.; Li, Y. Per-cell histone acetylation is associated with terminal differentiation in human T cells. Clin. Epigenetics 2024, 16, 21. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Si, X.; Shao, M.; Teng, X.; Huang, Y.; Meng, Y.; Wu, L.; Wei, J.; Liu, L.; Gu, T.; Song, J.; et al. Mitochondrial isocitrate dehydrogenase impedes CAR T cell function by restraining antioxidant metabolism and histone acetylation. Cell Metab. 2024, 36, 176–192.e10. [Google Scholar] [CrossRef] [PubMed]
- Geiger, R.; Rieckmann, J.C.; Wolf, T.; Basso, C.; Feng, Y.; Fuhrer, T.; Kogadeeva, M.; Picotti, P.; Meissner, F.; Mann, M.; et al. L-Arginine Modulates T Cell Metabolism and Enhances Survival and Anti-tumor Activity. Cell 2016, 167, 829–842.e13. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Medvedovic, J.; Ebert, A.; Tagoh, H.; Busslinger, M. Pax5: A master regulator of B cell development and leukemogenesis. Adv. Immunol. 2011, 111, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Delogu, A.; Schebesta, A.; Sun, Q.; Aschenbrenner, K.; Perlot, T.; Busslinger, M. Gene repression by Pax5 in B cells is essential for blood cell homeostasis and is reversed in plasma cells. Immunity 2006, 24, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Cobaleda, C.; Schebesta, A.; Delogu, A.; Busslinger, M. Pax5: The guardian of B cell identity and function. Nat. Immunol. 2007, 8, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Schebesta, A.; McManus, S.; Salvagiotto, G.; Delogu, A.; Busslinger, G.A.; Busslinger, M. Transcription factor Pax5 activates the chromatin of key genes involved in B cell signaling, adhesion, migration, and immune function. Immunity 2007, 27, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Revilla, I.D.R.; Bilic, I.; Vilagos, B.; Tagoh, H.; Ebert, A.; Tamir, I.M.; Smeenk, L.; Trupke, J.; Sommer, A.; Jaritz, M.; et al. The B-cell identity factor Pax5 regulates distinct transcriptional programmes in early and late B lymphopoiesis. Embo J. 2012, 31, 3130–3146. [Google Scholar] [CrossRef] [PubMed Central]
- Kumari, P.; Tarighi, S.; Braun, T.; Ianni, A. SIRT7 Acts as a Guardian of Cellular Integrity by Controlling Nucleolar and Extra-Nucleolar Functions. Genes 2021, 12, 1361. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mann, B.S.; Johnson, J.R.; Cohen, M.H.; Justice, R.; Pazdur, R. FDA Approval Summary: Vorinostat for Treatment of Advanced Primary Cutaneous T-Cell Lymphoma. Oncologist 2007, 12, 1247–1252. [Google Scholar] [CrossRef]
- Vogl, D.T.; Raje, N.; Jagannath, S.; Richardson, P.; Hari, P.; Orlowski, R.; Supko, J.G.; Tamang, D.; Yang, M.; Jones, S.S.; et al. Ricolinostat, the First Selective Histone Deacetylase 6 Inhibitor, in Combination with Bortezomib and Dexamethasone for Relapsed or Refractory Multiple Myeloma. Clin. Cancer Res. 2017, 23, 3307–3315. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bates, S.E.; Zhan, Z.; Steadman, K.; Obrzut, T.; Luchenko, V.; Frye, R.; Robey, R.W.; Turner, M.; Gardner, E.R.; Figg, W.D.; et al. Laboratory correlates for a phase II trial of romidepsin in cutaneous and peripheral T-cell lymphoma. Br. J. Haematol. 2010, 148, 256–267. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Greshock, T.J.; Johns, D.M.; Noguchi, Y.; Williams, R.M. Improved total synthesis of the potent HDAC inhibitor FK228 (FR-901228). Org. Lett. 2008, 10, 613–616. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, J.; Zhao, F.; Qu, Y. Lactylation: A Novel Post-Translational Modification with Clinical Implications in CNS Diseases. Biomolecules 2024, 14, 1175. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Virág, D.; Dalmadi-Kiss, B.; Vékey, K.; Drahos, L.; Klebovich, I.; Antal, I.; Ludányi, K. Current Trends in the Analysis of Post-translational Modifications. Chromatographia 2020, 83, 1–10. [Google Scholar] [CrossRef]
- Zhai, L.-H.; Chen, K.-F.; Hao, B.-B.; Tan, M.-J. Proteomic characterization of post-translational modifications in drug discovery. Acta Pharmacol. Sin. 2022, 43, 3112–3129. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune Cells | Factor | Impact | Effect (+: Positive; −: Negative) | References |
|---|---|---|---|---|
| Neutrophils | PSGL-1 | Cell activation, migration | + | [11] |
| AzurocidinCG | Immune response | + | [12,13] | |
| GlycoRNA | Modulation of the inflammatory response | + + | [14,15] | |
| NK cells | Siglec-7, Siglec-9 | Killing activity | − | [16,17] |
| CD16a | Killing activity | + | [18] | |
| Macrophages | Agglutinin (DC-SIGN) | Immune response | − | [10] |
| Pro-inflammatory cytokine | Immune response | + | [13,19] | |
| CD24, CD47 | Phagocytosis | − | [20] | |
| T cells | OGT | Cell development | − | [21] |
| Mgat1 | Cell development | − | [22,23] | |
| IL-2 | Cell differentiation Treg | + | [24] | |
| PD-1 | Cell activation | − | [25,26] | |
| B cells | μ-heavy chain | Pre-BCR | + | [27] |
| Galectin-1 | Cell maturation | + | [28] | |
| CD19 | Cell development | + | [29] |
| Immune Cells | Acylation Type | Factor | Impact | Effect | References |
|---|---|---|---|---|---|
| Macrophages | Acetylation Succinylation Crotonylation | NLRP3 | Inflammation | + | [54] |
| STAT6 | Cell polarization | + | [4] | ||
| CBP | Cell polarization | − | [55,56] | ||
| LPS | Inflammatory | + | [57] | ||
| H3K9cr/ACSS2 | Cell activation | + | [58] | ||
| AF9/YEATS2 | Gene transcription | + | [59] | ||
| T cells | Acetylation Succinylation | Acetate | Cell metabolism | + | [60] |
| Enzyme (KAT2A, SIRT7) | Cancer progress | +/− | [61] | ||
| B cells | Acetylation | PCAF | Cell function | + | [62,63,64] |
| Pax5/SIRT7 | Cell development | + | [65] | ||
| Succinylation | LYN | BCR signal transduction | + | [66,67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Zhang, Y.; Luo, X.; Zheng, X.; Bai, G.; Liu, J. Glycosylation and Acylation: Important Regulators of Immune Cell Fate Decisions. Biology 2025, 14, 611. https://doi.org/10.3390/biology14060611
Wang H, Zhang Y, Luo X, Zheng X, Bai G, Liu J. Glycosylation and Acylation: Important Regulators of Immune Cell Fate Decisions. Biology. 2025; 14(6):611. https://doi.org/10.3390/biology14060611
Chicago/Turabian StyleWang, Han, Yiying Zhang, Xu Luo, Xinxin Zheng, Guangdong Bai, and Junhui Liu. 2025. "Glycosylation and Acylation: Important Regulators of Immune Cell Fate Decisions" Biology 14, no. 6: 611. https://doi.org/10.3390/biology14060611
APA StyleWang, H., Zhang, Y., Luo, X., Zheng, X., Bai, G., & Liu, J. (2025). Glycosylation and Acylation: Important Regulators of Immune Cell Fate Decisions. Biology, 14(6), 611. https://doi.org/10.3390/biology14060611