Advances in Imaging Techniques for Mammalian/Human Ciliated Cell’s Cilia: Insights into Structure, Function, and Dynamics


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Fundamental Properties of Cilia
2.1. Definition, Structure, and Function
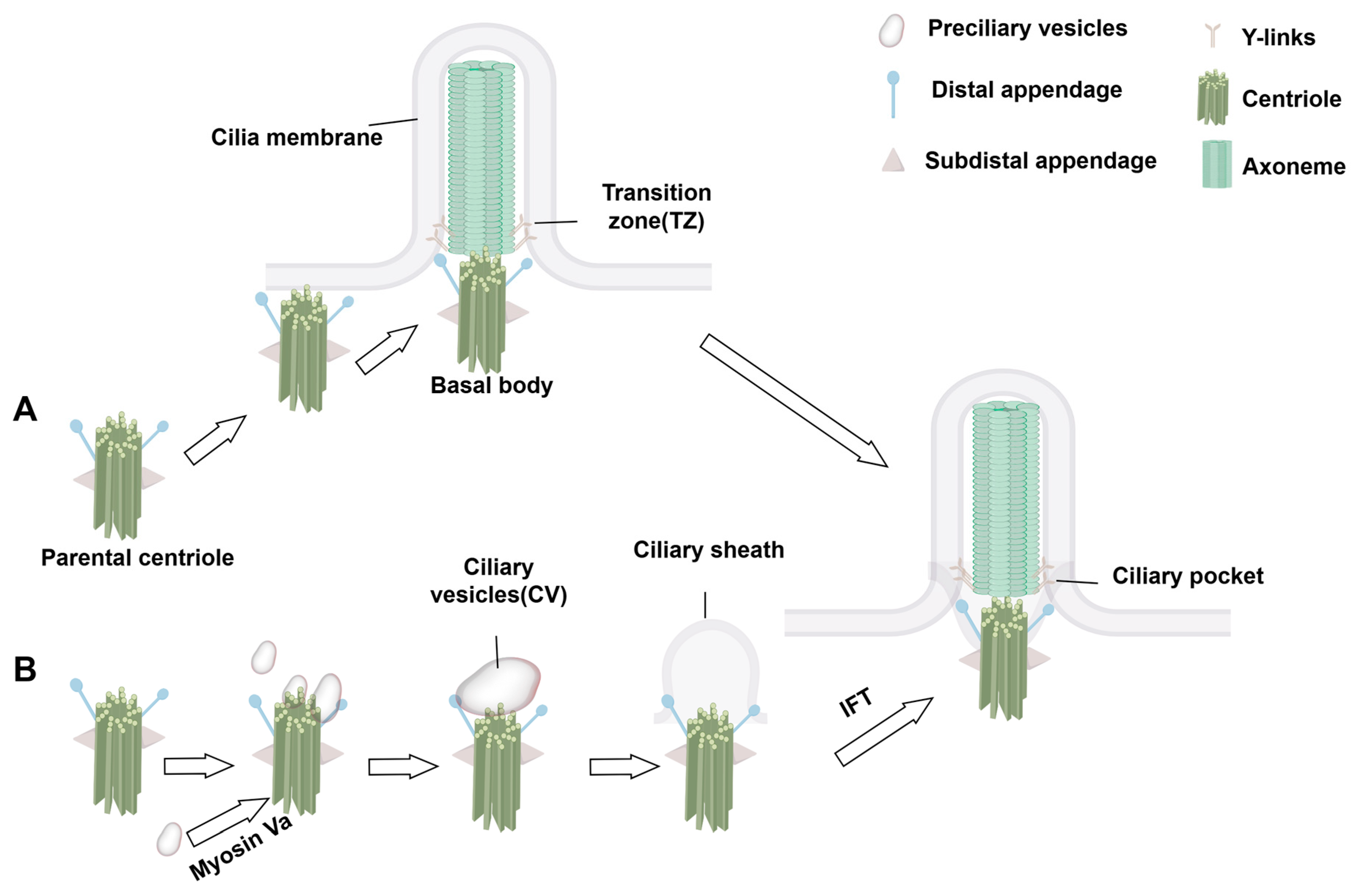
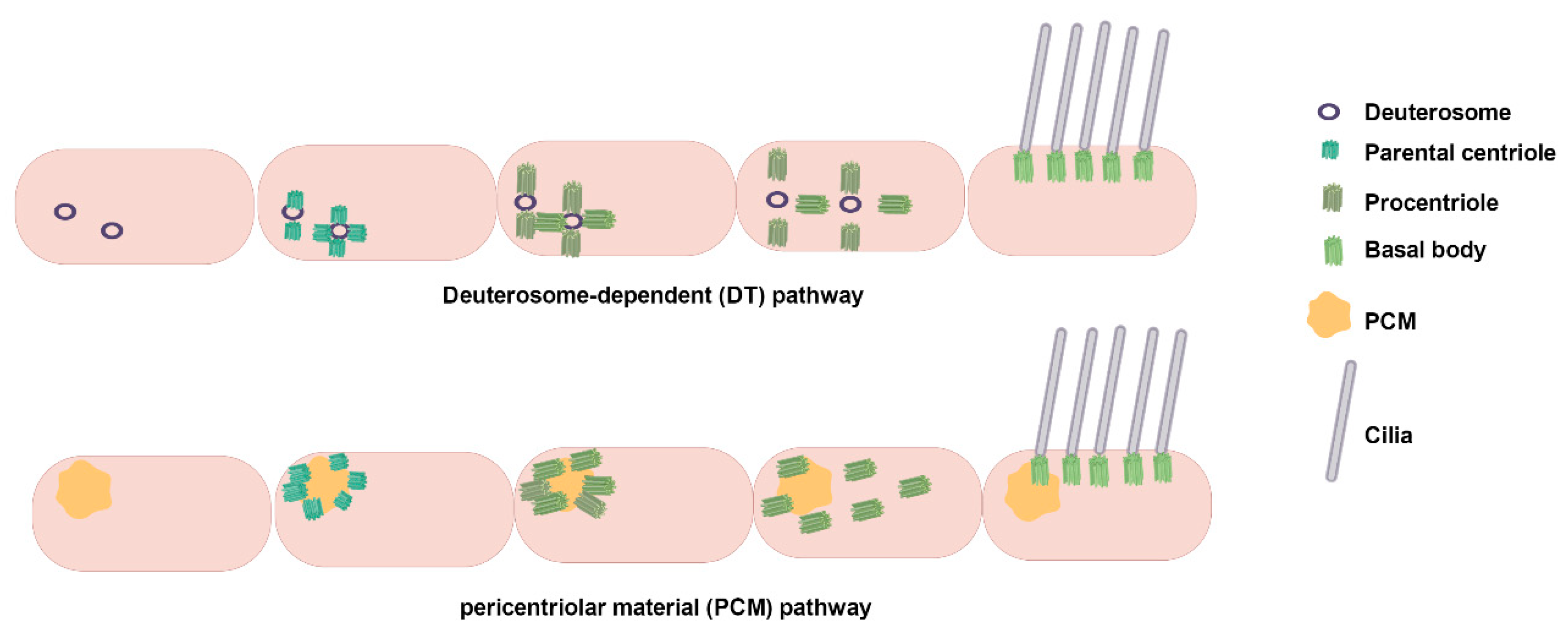
2.2. Assembly Dynamics
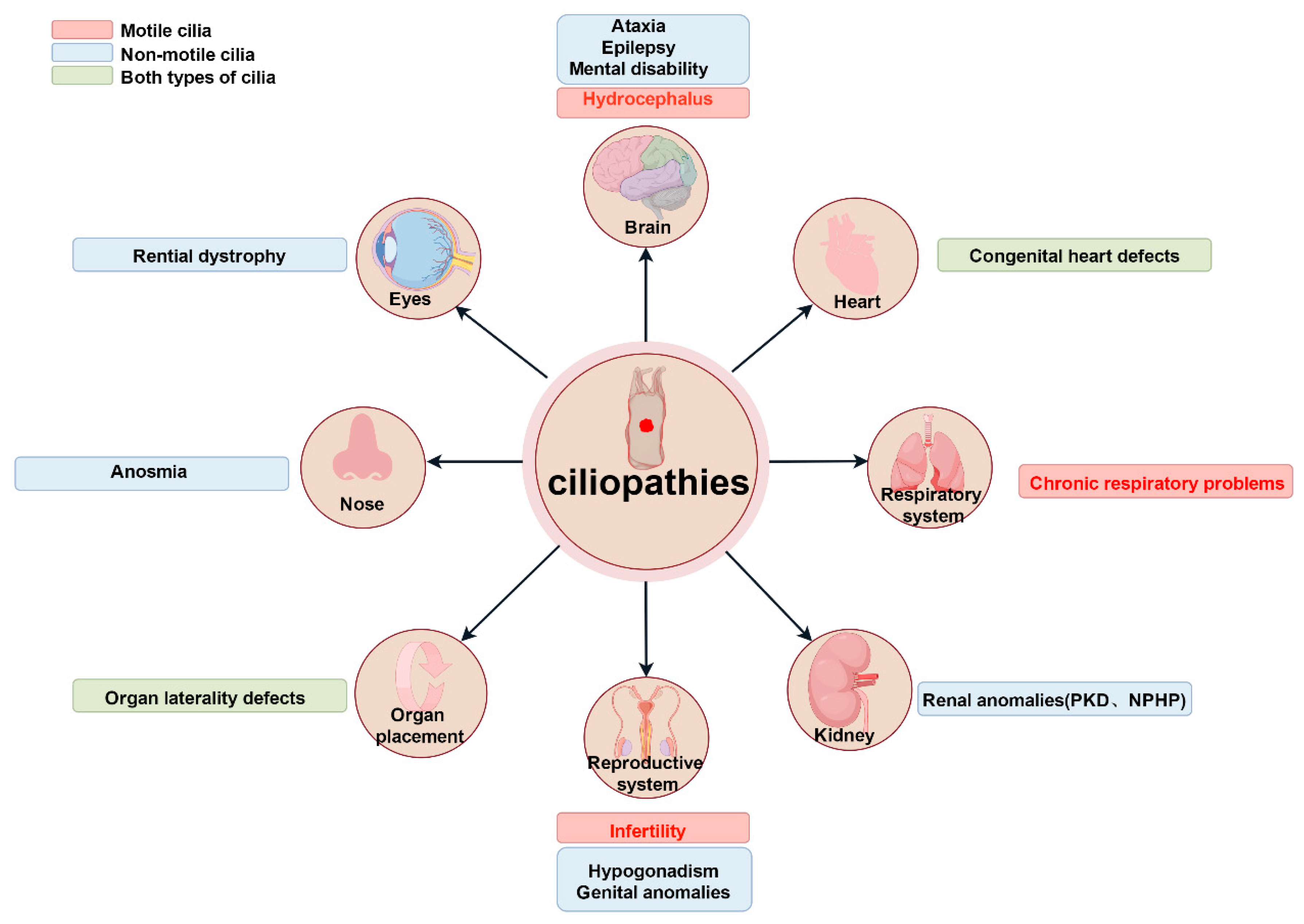
3. Research on Cilia in Organs and Organ Systems
3.1. Role of Cilia in Spermatogenesis
3.2. Sperm Flagellum Structure and Male Infertility
3.3. Ciliay Function in the Efferent Ductules (EDs)
3.4. Ependymal Cell (EC) Cilia and Hydrocephalus
3.5. Neuronal Cilia and Nervous System Diseases
4. Advancements in Cilia Imaging Technology
4.1. Optical Imaging
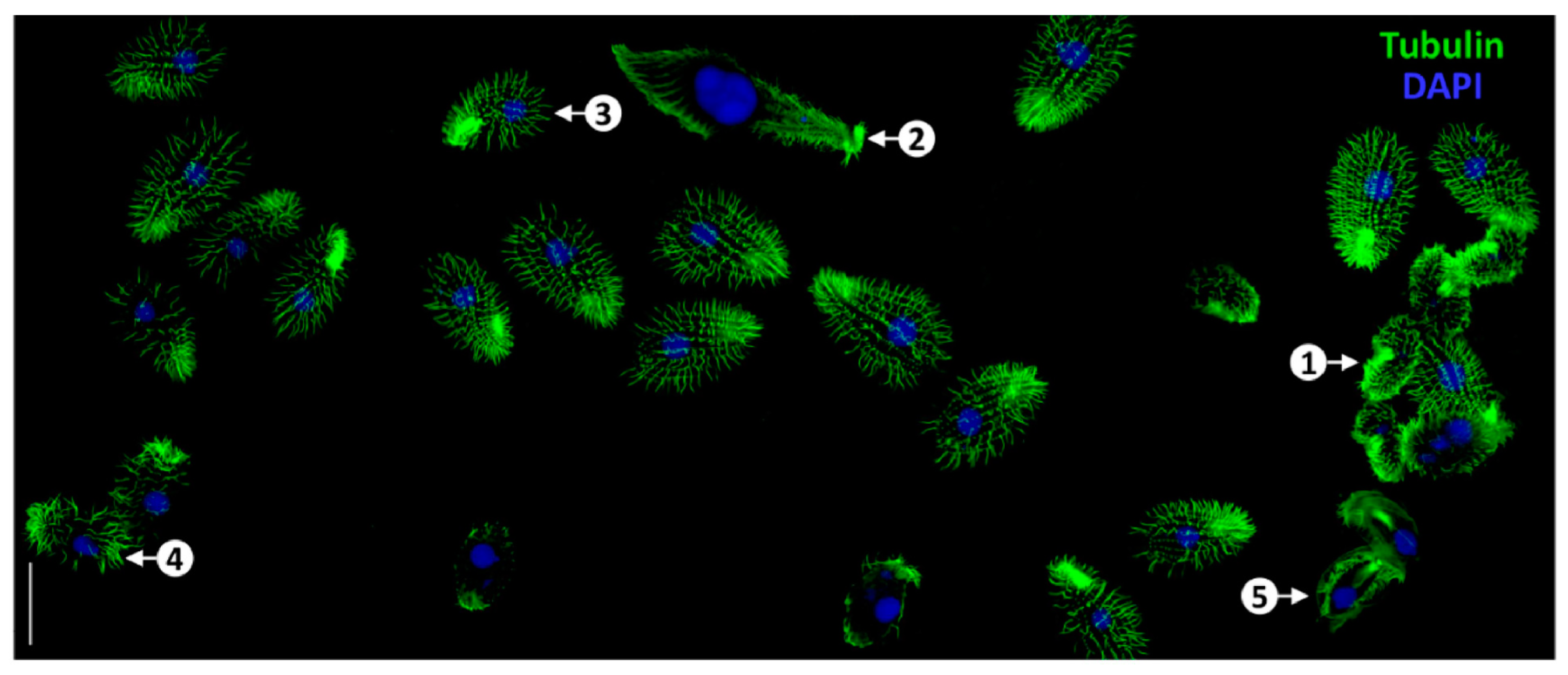
4.1.1. Conventional Optical Imaging
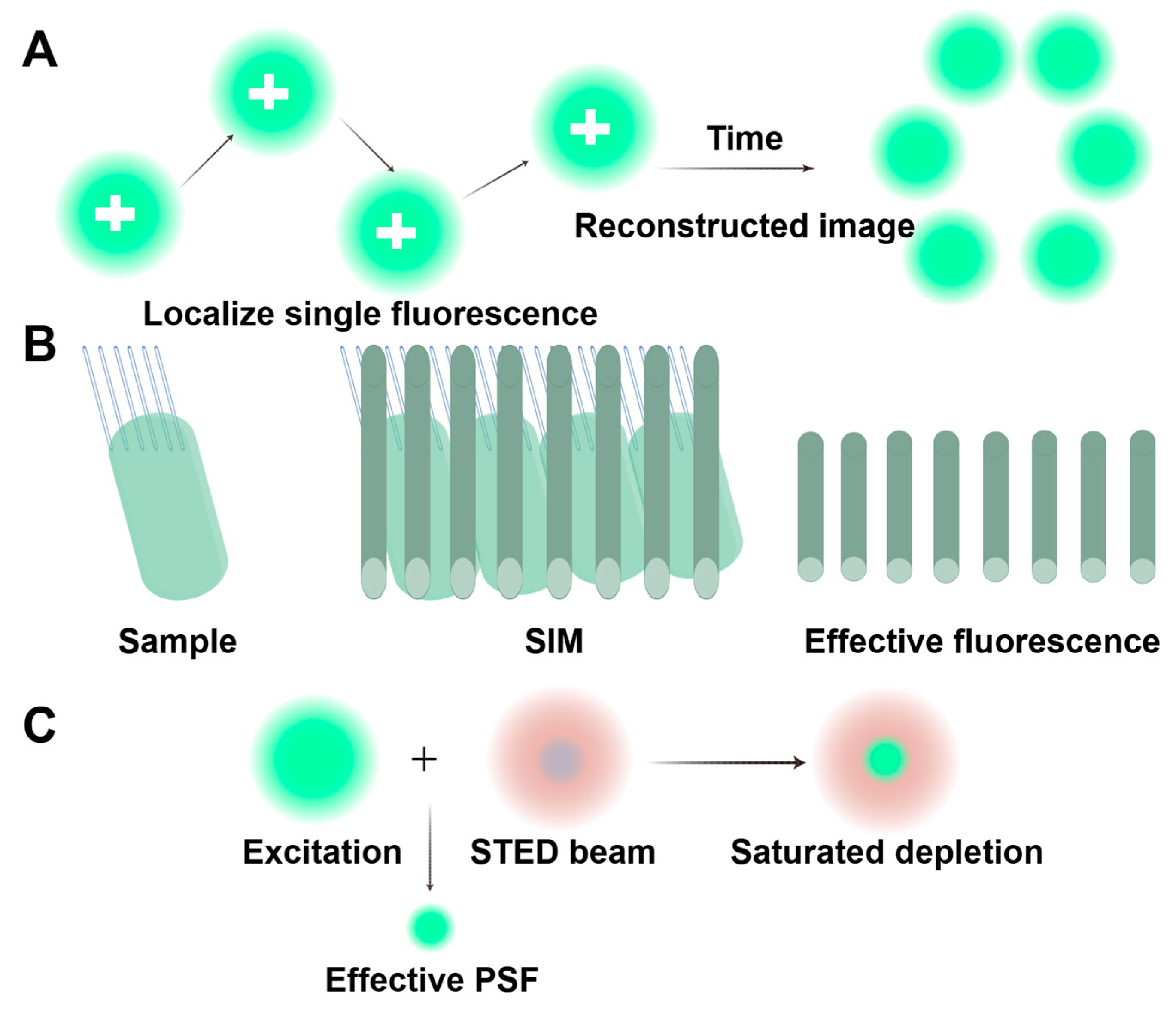
4.1.2. Optical Imaging Technology: Pushing the Limits of Conventional Optical Diffraction
4.2. Electron Microscopy (EM) Imaging
4.2.1. Transmission EM (TEM)
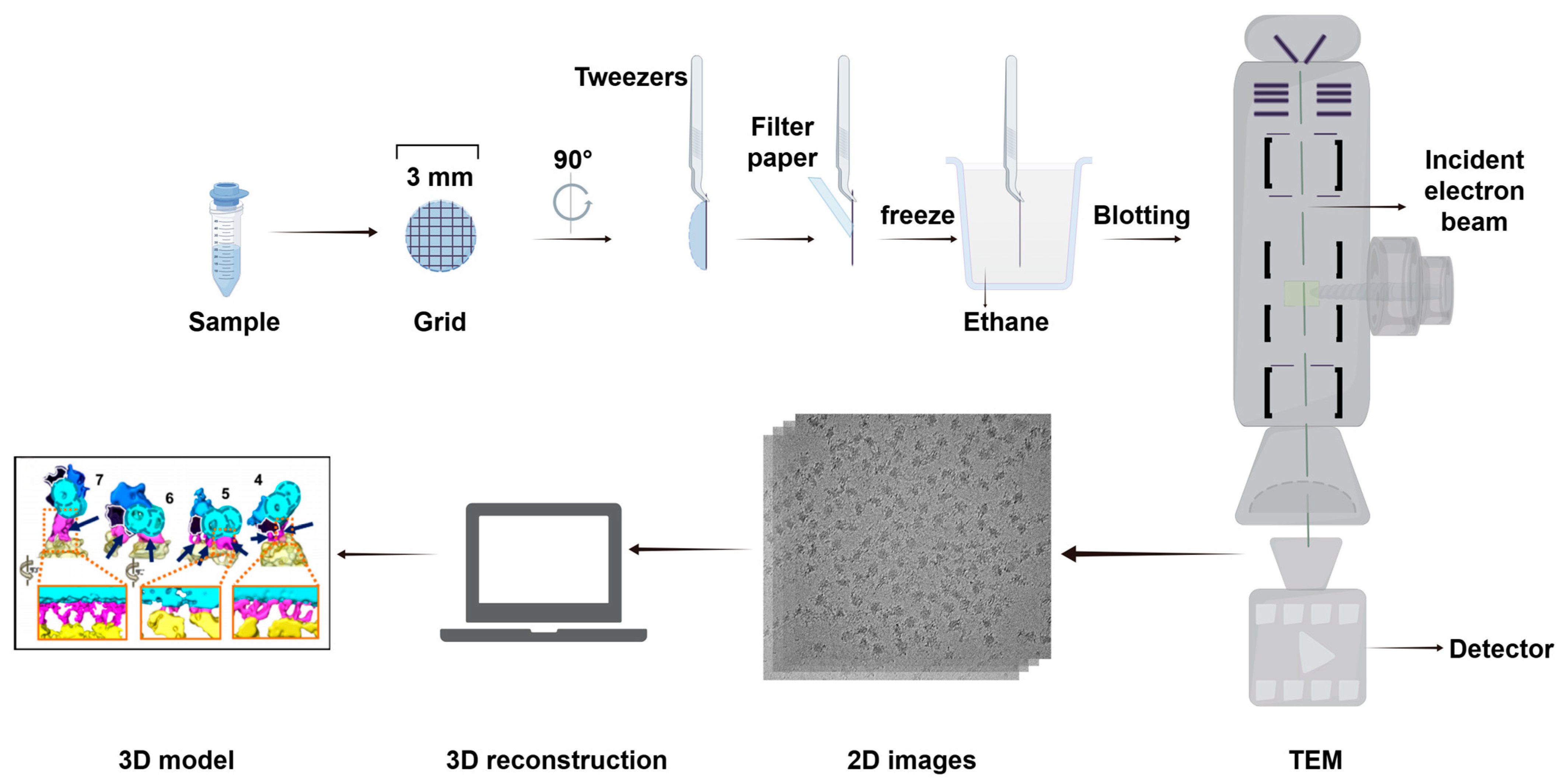
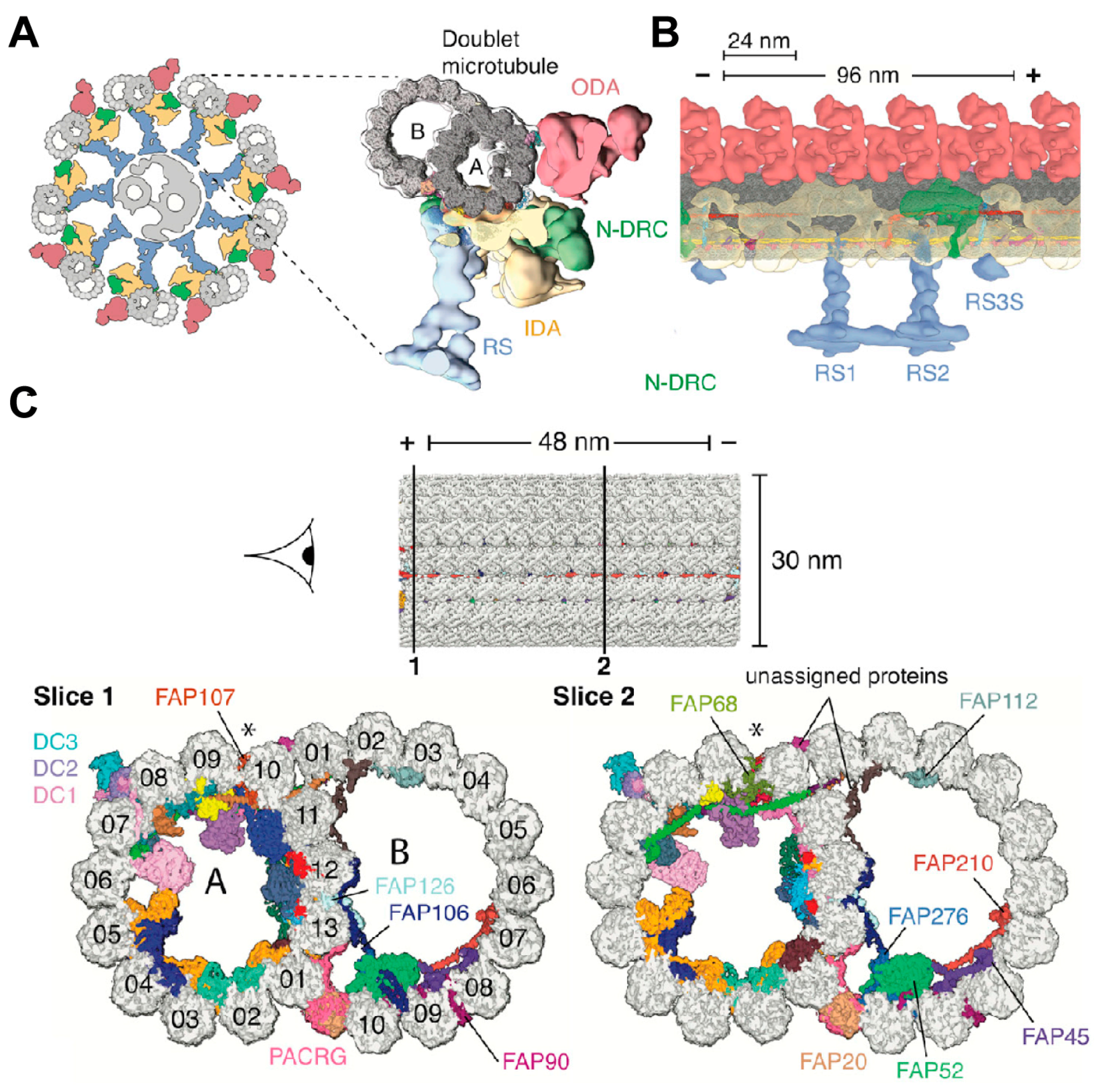
4.2.2. Cryo-EM
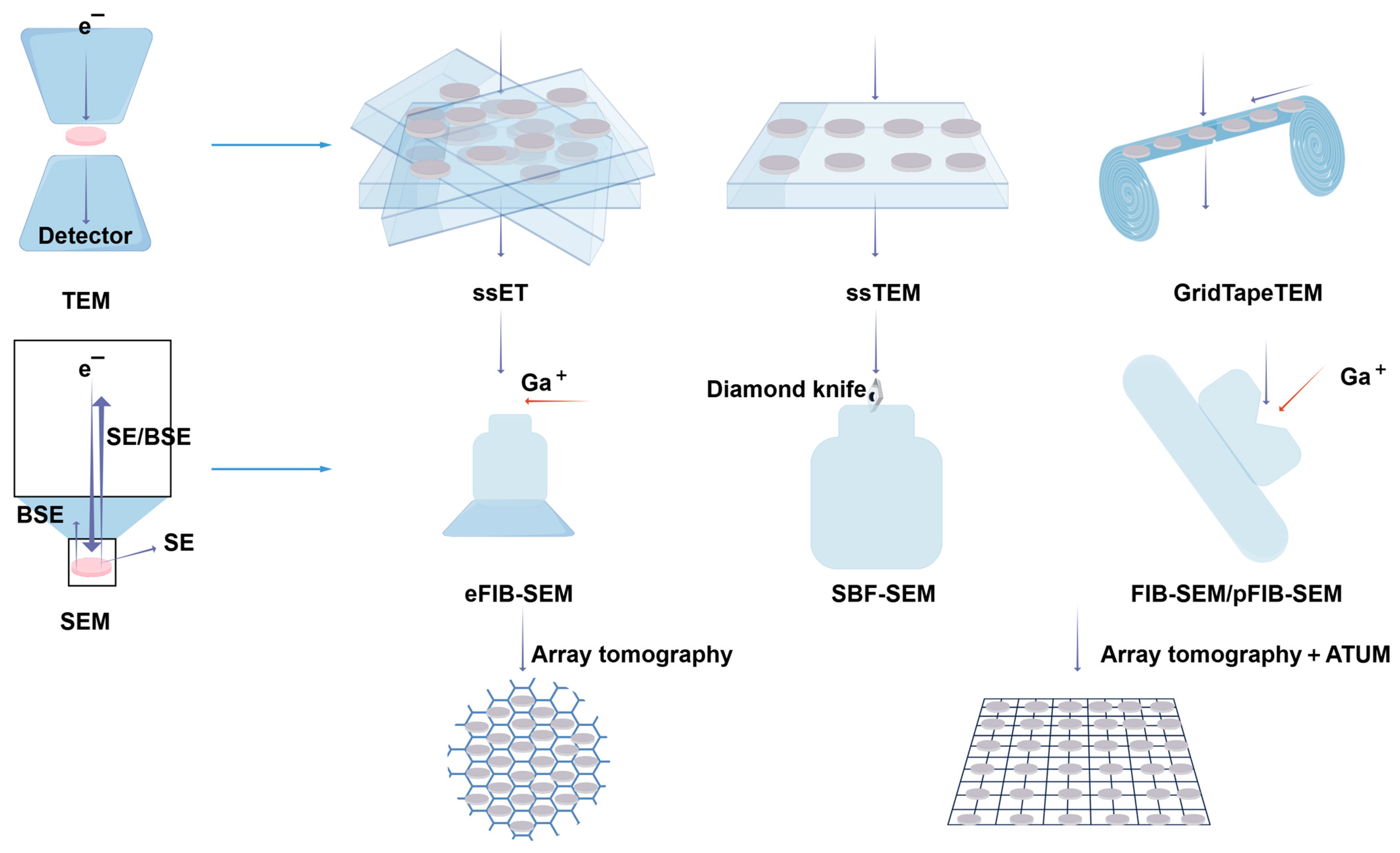
4.2.3. Volume EM (vEM)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sleigh, M.A.; Blake, J.R.; Liron, N. The propulsion of mucus by cilia. Am. Rev. Respir. Dis. 1988, 137, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Modaresi, M.A. Numerical investigation of mucociliary clearance using power law and thixotropic mucus layers under discrete and continuous cilia motion. Biomech. Model. Mechanobiol. 2023, 22, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.L. Aging Diminishes Mucociliary Clearance of the Lung. Adv. Geriatr. Med. Res. 2022, 4, e220005. [Google Scholar] [CrossRef]
- Barton, B.E.; Herrera, G.G.; Anamthathmakula, P.; Rock, J.K.; Willie, A.; Harris, E.A.; Takemaru, K.I.; Winuthayanon, W. Roles of steroid hormones in oviductal function. Reproduction 2020, 159, R125–R137. [Google Scholar] [CrossRef]
- Yoke, H.; Taniguchi, A.; Nonaka, S. Left-right asymmetry is formed in the basal bodies of the mouse node cilia in a cilia motility-dependent manner. bioRxiv 2023. [Google Scholar] [CrossRef]
- Amack, J.D. Salient features of the ciliated organ of asymmetry. Bioarchitecture 2014, 4, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Umair, Z.; Kumar, S.; Goutam, R.S.; Park, S.; Kim, J. The regulatory roles of motile cilia in CSF circulation and hydrocephalus. Fluids Barriers CNS 2021, 18, 31. [Google Scholar] [CrossRef]
- Antony van Leeuwenhoek and his “Little Animals”: Being some Account of the Father of Protozoology and Bacteriology and his Multifarious Discoveries in these Disciplines. Nature 1932, 130, 679–680. [CrossRef]
- Fliegauf, M.; Benzing, T.; Omran, H. When cilia go bad: Cilia defects and ciliopathies. Nat. Rev. Mol. Cell Biol. 2007, 8, 880–893. [Google Scholar] [CrossRef]
- Besschetnova, T.Y.; Kolpakova-Hart, E.; Guan, Y.; Zhou, J.; Olsen, B.R.; Shah, J.V. Identification of signaling pathways regulating primary cilium length and flow-mediated adaptation. Curr. Biol. 2010, 20, 182–187. [Google Scholar] [CrossRef]
- Novas, R.; Cardenas-Rodriguez, M.; Lepanto, P.; Fabregat, M.; Rodao, M.; Fariello, M.I.; Ramos, M.; Davison, C.; Casanova, G.; Alfaya, L.; et al. Kinesin 1 regulates cilia length through an interaction with the Bardet-Biedl syndrome related protein CCDC28B. Sci. Rep. 2018, 8, 3019. [Google Scholar] [CrossRef] [PubMed]
- Reiter, J.F.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Duldulao, N.A.; Li, J.; Sun, Z. Cilia in cell signaling and human disorders. Protein Cell 2010, 1, 726–736. [Google Scholar] [CrossRef]
- Amack, J.D. Structures and functions of cilia during vertebrate embryo development. Mol. Reprod. Dev. 2022, 89, 579–596. [Google Scholar] [CrossRef]
- Ishikawa, T. Axoneme Structure from Motile Cilia. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef]
- Klena, N.; Pigino, G. Structural Biology of Cilia and Intraflagellar Transport. Annu. Rev. Cell Dev. Biol. 2022, 38, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Christensen, S.T. Overview of structure and function of mammalian cilia. Annu. Rev. Physiol. 2007, 69, 377–400. [Google Scholar] [CrossRef]
- Spassky, N.; Meunier, A. The development and functions of multiciliated epithelia. Nat. Rev. Mol. Cell Biol. 2017, 18, 423–436. [Google Scholar] [CrossRef]
- Yuan, S.; Liu, Y.; Peng, H.; Tang, C.; Hennig, G.W.; Wang, Z.; Wang, L.; Yu, T.; Klukovich, R.; Zhang, Y.; et al. Motile cilia of the male reproductive system require miR-34/miR-449 for development and function to generate luminal turbulence. Proc. Natl. Acad. Sci. USA 2019, 116, 3584–3593. [Google Scholar] [CrossRef]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef]
- Nordgren, T.M.; Wyatt, T.A.; Sweeter, J.; Bailey, K.L.; Poole, J.A.; Heires, A.J.; Sisson, J.H.; Romberger, D.J. Motile cilia harbor serum response factor as a mechanism of environment sensing and injury response in the airway. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 306, L829–L839. [Google Scholar] [CrossRef] [PubMed]
- Bloodgood, R.A. Sensory reception is an attribute of both primary cilia and motile cilia. J. Cell Sci. 2010, 123, 505–509. [Google Scholar] [CrossRef]
- Gopalakrishnan, J.; Feistel, K.; Friedrich, B.M.; Grapin-Botton, A.; Jurisch-Yaksi, N.; Mass, E.; Mick, D.U.; Müller, R.U.; May-Simera, H.; Schermer, B.; et al. Emerging principles of primary cilia dynamics in controlling tissue organization and function. Embo J. 2023, 42, e113891. [Google Scholar] [CrossRef] [PubMed]
- Breslow, D.K.; Holland, A.J. Mechanism and Regulation of Centriole and Cilium Biogenesis. Annu. Rev. Biochem. 2019, 88, 691–724. [Google Scholar] [CrossRef]
- Zhao, H.; Khan, Z.; Westlake, C.J. Ciliogenesis membrane dynamics and organization. Semin. Cell Dev. Biol. 2023, 133, 20–31. [Google Scholar] [CrossRef]
- Ou, G.; Scholey, J.M. Motor Cooperation During Mitosis and Ciliogenesis. Annu. Rev. Cell Dev. Biol. 2022, 38, 49–74. [Google Scholar] [CrossRef]
- Yadav, S.P.; Sharma, N.K.; Liu, C.; Dong, L.; Li, T.; Swaroop, A. Centrosomal protein CP110 controls maturation of the mother centriole during cilia biogenesis. Development 2016, 143, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Dynlacht, B.D. Assembling a primary cilium. Curr. Opin. Cell Biol. 2013, 25, 506–511. [Google Scholar] [CrossRef]
- Pigino, G. Intraflagellar transport. Curr. Biol. 2021, 31, R530–R536. [Google Scholar] [CrossRef]
- Rayamajhi, D.; Roy, S. Multiciliated Cells: Rise and Fall of the Deuterosomes. Trends Cell Biol. 2020, 30, 259–262. [Google Scholar] [CrossRef]
- Liu, H.; Zheng, J.; Zhu, L.; Xie, L.; Chen, Y.; Zhang, Y.; Zhang, W.; Yin, Y.; Peng, C.; Zhou, J.; et al. Wdr47, Camsaps, and Katanin cooperate to generate ciliary central microtubules. Nat. Commun. 2021, 12, 5796. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Marszalek, J.R.; Ruiz-Lozano, P.; Roberts, E.; Chien, K.R.; Goldstein, L.S. Situs inversus and embryonic ciliary morphogenesis defects in mouse mutants lacking the KIF3A subunit of kinesin-II. Proc. Natl. Acad. Sci. USA 1999, 96, 5043–5048. [Google Scholar] [CrossRef]
- Takeda, S.; Yonekawa, Y.; Tanaka, Y.; Okada, Y.; Nonaka, S.; Hirokawa, N. Left-right asymmetry and kinesin superfamily protein KIF3A: New insights in determination of laterality and mesoderm induction by kif3A-/- mice analysis. J. Cell Biol. 1999, 145, 825–836. [Google Scholar] [CrossRef]
- Radhakrishnan, P.; Nayak, S.S.; Shukla, A.; Lindstrand, A.; Girisha, K.M. Meckel syndrome: Clinical and mutation profile in six fetuses. Clin. Genet. 2019, 96, 560–565. [Google Scholar] [CrossRef]
- Melluso, A.; Secondulfo, F.; Capolongo, G.; Capasso, G.; Zacchia, M. Bardet-Biedl Syndrome: Current Perspectives and Clinical Outlook. Ther. Clin. Risk Manag. 2023, 19, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.D.; Maffei, P.; Collin, G.B.; Naggert, J.K. Alström syndrome: Genetics and clinical overview. Curr. Genom. 2011, 12, 225–235. [Google Scholar] [CrossRef]
- Luo, F.; Tao, Y.H. Nephronophthisis: A review of genotype-phenotype correlation. Nephrology 2018, 23, 904–911. [Google Scholar] [CrossRef]
- Bergmann, C.; Guay-Woodford, L.M.; Harris, P.C.; Horie, S.; Peters, D.J.M.; Torres, V.E. Polycystic kidney disease. Nat. Rev. Dis. Primers 2018, 4, 50. [Google Scholar] [CrossRef]
- Horani, A.; Ferkol, T.W. Understanding Primary Ciliary Dyskinesia and Other Ciliopathies. J. Pediatr. 2021, 230, 15–22.e1. [Google Scholar] [CrossRef]
- Nygaard, M.B.; Almstrup, K.; Lindbæk, L.; Christensen, S.T.; Svingen, T. Cell context-specific expression of primary cilia in the human testis and ciliary coordination of Hedgehog signalling in mouse Leydig cells. Sci. Rep. 2015, 5, 10364. [Google Scholar] [CrossRef] [PubMed]
- Lara, N.d.L.e.M.; Elsenhans, A.; Dardari, R.; Dobrinski, I. The role of primary cilia in the testis. Curr. Opin. Endocr. Metab. Res. 2024, 34, e100508. [Google Scholar] [CrossRef]
- Xie, H.; Wang, X.; Jin, M.; Li, L.; Zhu, J.; Kang, Y.; Chen, Z.; Sun, Y.; Zhao, C. Cilia regulate meiotic recombination in zebrafish. J. Mol. Cell Biol. 2022, 14, mjac049. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xing, J.; Zhang, X.; Liu, H.; Liu, X.; Jiang, H.; Xu, C.; Zhao, X.; Hu, Z. Control of ciliary transcriptional programs during spermatogenesis by antagonistic transcription factors. Elife 2025, 13, RP94754. [Google Scholar] [CrossRef]
- Mytlis, A.; Kumar, V.; Qiu, T.; Deis, R.; Hart, N.; Levy, K.; Masek, M.; Shawahny, A.; Ahmad, A.; Eitan, H.; et al. Control of meiotic chromosomal bouquet and germ cell morphogenesis by the zygotene cilium. Science 2022, 376, eabh3104. [Google Scholar] [CrossRef]
- Lehti, M.S.; Sironen, A. Formation and function of sperm tail structures in association with sperm motility defects. Biol. Reprod. 2017, 97, 522–536. [Google Scholar] [CrossRef]
- Kumar, N.; Singh, A.K. The anatomy, movement, and functions of human sperm tail: An evolving mystery. Biol. Reprod. 2021, 104, 508–520. [Google Scholar] [CrossRef]
- Gu, N.H.; Zhao, W.L.; Wang, G.S.; Sun, F. Comparative analysis of mammalian sperm ultrastructure reveals relationships between sperm morphology, mitochondrial functions and motility. Reprod. Biol. Endocrinol. 2019, 17, 66. [Google Scholar] [CrossRef]
- Irons, M.J.; Clermont, Y. Formation of the outer dense fibers during spermiogenesis in the rat. Anat. Rec. 1982, 202, 463–471. [Google Scholar] [CrossRef]
- Zhao, W.; Li, Z.; Ping, P.; Wang, G.; Yuan, X.; Sun, F. Outer dense fibers stabilize the axoneme to maintain sperm motility. J. Cell Mol. Med. 2018, 22, 1755–1768. [Google Scholar] [CrossRef]
- Eddy, E.M. The scaffold role of the fibrous sheath. Soc. Reprod. Fertil. Suppl. 2007, 65, 45–62. [Google Scholar] [PubMed]
- Ben Khelifa, M.; Coutton, C.; Zouari, R.; Karaouzène, T.; Rendu, J.; Bidart, M.; Yassine, S.; Pierre, V.; Delaroche, J.; Hennebicq, S.; et al. Mutations in DNAH1, which encodes an inner arm heavy chain dynein, lead to male infertility from multiple morphological abnormalities of the sperm flagella. Am. J. Hum. Genet. 2014, 94, 95–104. [Google Scholar] [CrossRef]
- Touré, A.; Martinez, G.; Kherraf, Z.E.; Cazin, C.; Beurois, J.; Arnoult, C.; Ray, P.F.; Coutton, C. The genetic architecture of morphological abnormalities of the sperm tail. Hum. Genet. 2021, 140, 21–42. [Google Scholar] [CrossRef]
- Nsota Mbango, J.F.; Coutton, C.; Arnoult, C.; Ray, P.F.; Touré, A. Genetic causes of male infertility: Snapshot on morphological abnormalities of the sperm flagellum. Basic. Clin. Androl. 2019, 29, 2. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.; Kim, E.N.; Chen, D.; Li, F.Q.; Takemaru, K.I. Essential Roles of Efferent Duct Multicilia in Male Fertility. Cells 2022, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Hess, R.A. Small tubules, surprising discoveries: From efferent ductules in the turkey to the discovery that estrogen receptor alpha is essential for fertility in the male. Anim. Reprod. 2015, 12, 7–23. [Google Scholar]
- Hess, R.A. Estrogen in the adult male reproductive tract: A review. Reprod. Biol. Endocrinol. 2003, 1, 52. [Google Scholar] [CrossRef]
- Clulow, J.; Jones, R.C.; Hansen, L.A.; Man, S.Y. Fluid and electrolyte reabsorption in the ductuli efferentes testis. J. Reprod. Fertil. Suppl. 1998, 53, 1–14. [Google Scholar]
- Hess, R.A.; Bunick, D.; Lee, K.H.; Bahr, J.; Taylor, J.A.; Korach, K.S.; Lubahn, D.B. A role for oestrogens in the male reproductive system. Nature 1997, 390, 509–512. [Google Scholar] [CrossRef]
- Hansen, L.A.; Clulow, J.; Jones, R.C. The role of Na+-H+ exchange in fluid and solute transport in the rat efferent ducts. Exp. Physiol. 1999, 84, 521–527. [Google Scholar]
- Mirzadeh, Z.; Han, Y.G.; Soriano-Navarro, M.; García-Verdugo, J.M.; Alvarez-Buylla, A. Cilia organize ependymal planar polarity. J. Neurosci. 2010, 30, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Mirzadeh, Z.; Merkle, F.T.; Soriano-Navarro, M.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Neural stem cells confer unique pinwheel architecture to the ventricular surface in neurogenic regions of the adult brain. Cell Stem Cell 2008, 3, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Ohata, S.; Nakatani, J.; Herranz-Pérez, V.; Cheng, J.; Belinson, H.; Inubushi, T.; Snider, W.D.; García-Verdugo, J.M.; Wynshaw-Boris, A.; Alvarez-Buylla, A. Loss of Dishevelleds disrupts planar polarity in ependymal motile cilia and results in hydrocephalus. Neuron 2014, 83, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, X.; Guo, J.; Yu, C.; Yang, J. Molecular Mechanisms and Risk Factors for the Pathogenesis of Hydrocephalus. Front. Genet. 2021, 12, 777926. [Google Scholar] [CrossRef]
- Wallmeier, J.; Dallmayer, M.; Omran, H. The role of cilia for hydrocephalus formation. Am. J. Med. Genet. C Semin. Med. Genet. 2022, 190, 47–56. [Google Scholar] [CrossRef]
- Tully, H.M.; Dobyns, W.B. Infantile hydrocephalus: A review of epidemiology, classification and causes. Eur. J. Med. Genet. 2014, 57, 359–368. [Google Scholar] [CrossRef]
- Liu, X.Y.; Song, X.; Czosnyka, M.; Robba, C.; Czosnyka, Z.; Summers, J.L.; Yu, H.J.; Gao, G.Y.; Smielewski, P.; Guo, F.; et al. Congenital hydrocephalus: A review of recent advances in genetic etiology and molecular mechanisms. Mil. Med. Res. 2024, 11, 54. [Google Scholar] [CrossRef]
- Wu, J.Y.; Cho, S.J.; Descant, K.; Li, P.H.; Shapson-Coe, A.; Januszewski, M.; Berger, D.R.; Meyer, C.; Casingal, C.; Huda, A.; et al. Mapping of neuronal and glial primary cilia contactome and connectome in the human cerebral cortex. Neuron 2024, 112, 41–55.e3. [Google Scholar] [CrossRef]
- Ott, C.M.; Torres, R.; Kuan, T.S.; Kuan, A.; Buchanan, J.; Elabbady, L.; Seshamani, S.; Bodor, A.L.; Collman, F.; Bock, D.D.; et al. Nanometer-scale views of visual cortex reveal anatomical features of primary cilia poised to detect synaptic spillover. bioRxiv 2023. [Google Scholar] [CrossRef]
- Ott, C.M.; Torres, R.; Kuan, T.S.; Kuan, A.; Buchanan, J.; Elabbady, L.; Seshamani, S.; Bodor, A.L.; Collman, F.; Bock, D.D.; et al. Ultrastructural differences impact cilia shape and external exposure across cell classes in the visual cortex. Curr. Biol. 2024, 34, 2418–2433.e4. [Google Scholar] [CrossRef]
- Senatore, E.; Iannucci, R.; Chiuso, F.; Delle Donne, R.; Rinaldi, L.; Feliciello, A. Pathophysiology of Primary Cilia: Signaling and Proteostasis Regulation. Front. Cell Dev. Biol. 2022, 10, 833086. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.X.; Miao, Y.T.; Zhang, X.; Huang, M.Z.; Jahangir, M.; Luo, S.; Lang, B. Primary cilia-associated protein IFT172 in ciliopathies. Front. Cell Dev. Biol. 2023, 11, 1074880. [Google Scholar] [CrossRef]
- Pfirrmann, T.; Franco, B.; Kopinke, D.; Gerhardt, C. Editorial: Regulation of proteostasis and cellular energy homeostasis at the primary cilium. Front. Cell Dev. Biol. 2023, 11, 1285237. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; Goyal, R.; Panoutsopoulos, A.A.; Balashova, O.A.; Lee, D.; Borodinsky, L.N. Calcium dynamics at the neural cell primary cilium regulate Hedgehog signaling-dependent neurogenesis in the embryonic neural tube. Proc. Natl. Acad. Sci. USA 2023, 120, e2220037120. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.Q.; Li, S.; Xu, Y.L.; Zhang, Y.C.; Li, P.Y.; Liang, L.Y.; Song, G.P.; Jian, X.X.; Wu, M.; Song, Z.Q.; et al. Rhythmic cilia changes support SCN neuron coherence in circadian clock. Science 2023, 380, 972–979. [Google Scholar] [CrossRef]
- Wang, L.; Guo, Q.; Acharya, S.; Zheng, X.; Huynh, V.; Whitmore, B.; Yimit, A.; Malhotra, M.; Chatterji, S.; Rosin, N.; et al. Primary cilia signaling in astrocytes mediates development and regional-specific functional specification. Nat. Neurosci. 2024, 27, 1708–1720. [Google Scholar] [CrossRef]
- Ray, S.; Gurung, P.; Manning, R.S.; Kravchuk, A.A.; Singhvi, A. Neuron cilia restrain glial KCC-3 to a microdomain to regulate multisensory processing. Cell Rep. 2024, 43, 113844. [Google Scholar] [CrossRef] [PubMed]
- Rivagorda, M.; Romeo-Guitart, D.; Blanchet, V.; Mailliet, F.; Boitez, V.; Barry, N.; Milunov, D.; Siopi, E.; Goudin, N.; Moriceau, S.; et al. A primary cilia-autophagy axis in hippocampal neurons is essential to maintain cognitive resilience. Nat. Aging 2025, 5, 450–467. [Google Scholar] [CrossRef]
- Yang, J.; Dong, Y.; Liu, J.; Peng, Y.; Wang, D.; Li, L.; Hu, X.; Li, J.; Wang, L.; Chu, J.; et al. Primary ciliary protein kinase A activity in the prefrontal cortex modulates stress in mice. Neuron 2025, 113, 1276–1289.e5. [Google Scholar] [CrossRef]
- Oya, M.; Miyasaka, Y.; Nakamura, Y.; Tanaka, M.; Suganami, T.; Mashimo, T.; Nakamura, K. Age-related ciliopathy: Obesogenic shortening of melanocortin-4 receptor-bearing neuronal primary cilia. Cell Metab. 2024, 36, 1044–1058.e10. [Google Scholar] [CrossRef]
- Jiang, Y.; Xun, Y.; Zhang, Z. Central regulation of feeding and body weight by ciliary GPR75. J. Clin. Investig. 2024, 134, e182121. [Google Scholar] [CrossRef] [PubMed]
- Satir, P. Landmarks in cilia research from Leeuwenhoek to us. Cell Motil. Cytoskelet. 1995, 32, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Amako, K. [Little animals observed by Antony van Leeuwenhoek]. Nihon Saikingaku Zasshi 2014, 69, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Bloodgood, R.A. From central to rudimentary to primary: The history of an underappreciated organelle whose time has come. The primary cilium. Methods Cell Biol. 2009, 94, 3–52. [Google Scholar] [CrossRef]
- Satir, P. CILIA: Before and after. Cilia 2017, 6, 1. [Google Scholar] [CrossRef]
- Fujimoto, J.G.; Pitris, C.; Boppart, S.A.; Brezinski, M.E. Optical coherence tomography: An emerging technology for biomedical imaging and optical biopsy. Neoplasia 2000, 2, 9–25. [Google Scholar] [CrossRef]
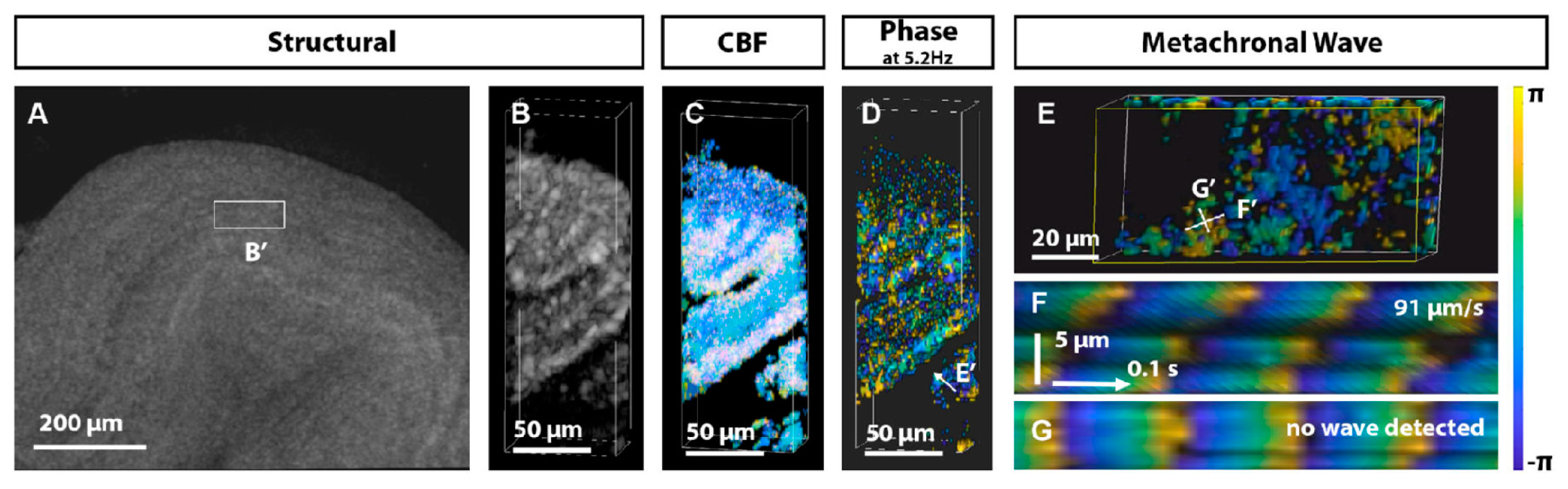
- Xia, T.; Umezu, K.; Scully, D.M.; Wang, S.; Larina, I.V. In vivo volumetric depth-resolved imaging of cilia metachronal waves using dynamic optical coherence tomography. Optica 2023, 10, 1439–1451. [Google Scholar] [CrossRef]
- Luo, P.G.; Sahu, S.; Yang, S.T.; Sonkar, S.K.; Wang, J.; Wang, H.; LeCroy, G.E.; Cao, L.; Sun, Y.P. Carbon “quantum” dots for optical bioimaging. J. Mater. Chem. B 2013, 1, 2116–2127. [Google Scholar] [CrossRef]
- Crivat, G.; Taraska, J.W. Imaging proteins inside cells with fluorescent tags. Trends Biotechnol. 2012, 30, 8–16. [Google Scholar] [CrossRef]
- Stender, A.S.; Marchuk, K.; Liu, C.; Sander, S.; Meyer, M.W.; Smith, E.A.; Neupane, B.; Wang, G.; Li, J.; Cheng, J.X.; et al. Single cell optical imaging and spectroscopy. Chem. Rev. 2013, 113, 2469–2527. [Google Scholar] [CrossRef]
- Phua, S.C.; Chiba, S.; Suzuki, M.; Su, E.; Roberson, E.C.; Pusapati, G.V.; Schurmans, S.; Setou, M.; Rohatgi, R.; Reiter, J.F.; et al. Dynamic Remodeling of Membrane Composition Drives Cell Cycle through Primary Cilia Excision. Cell 2019, 178, 261. [Google Scholar] [CrossRef]
- Toro-Tapia, G.; Das, R.M. Primary cilium remodeling mediates a cell signaling switch in differentiating neurons. Sci. Adv. 2020, 6, eabb0601. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, D.; Colin, E.; Louie, C.M.; Williams, D.S. Live-cell imaging evidence for the ciliary transport of rod photoreceptor opsin by heterotrimeric kinesin-2. J. Neurosci. 2012, 32, 10587–10593. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Breslow, D.K.; Koslover, E.F.; Spakowitz, A.J.; Nelson, W.J.; Nachury, M.V. Single molecule imaging reveals a major role for diffusion in the exploration of ciliary space by signaling receptors. Elife 2013, 2, e00654. [Google Scholar] [CrossRef]
- Gautier, A.; Juillerat, A.; Heinis, C.; Corrêa, I.R., Jr.; Kindermann, M.; Beaufils, F.; Johnsson, K. An engineered protein tag for multiprotein labeling in living cells. Chem. Biol. 2008, 15, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Los, G.V.; Encell, L.P.; McDougall, M.G.; Hartzell, D.D.; Karassina, N.; Zimprich, C.; Wood, M.G.; Learish, R.; Ohana, R.F.; Urh, M.; et al. HaloTag: A novel protein labeling technology for cell imaging and protein analysis. ACS Chem. Biol. 2008, 3, 373–382. [Google Scholar] [CrossRef]
- Lu, Q.; Insinna, C.; Ott, C.; Stauffer, J.; Pintado, P.A.; Rahajeng, J.; Baxa, U.; Walia, V.; Cuenca, A.; Hwang, Y.S.; et al. Early steps in primary cilium assembly require EHD1/EHD3-dependent ciliary vesicle formation. Nat. Cell Biol. 2015, 17, 228–240. [Google Scholar] [CrossRef]
- Diener, D. Analysis of cargo transport by IFT and GFP imaging of IFT in Chlamydomonas. Methods Cell Biol. 2009, 93, 111–119. [Google Scholar] [CrossRef]
- Ishikawa, H.; Marshall, W.F. Efficient live fluorescence imaging of intraflagellar transport in mammalian primary cilia. Methods Cell Biol. 2015, 127, 189–201. [Google Scholar] [CrossRef]
- Pan, T.; Zhang, J.; Hu, C.; Yan, G.; Qin, W.; Cheng, Z.; Yuan, D.; Zhou, F.; Xiong, J.; Miao, W.; et al. An improved method for tubulin staining of ciliated eukaryotes. Water Biol. Secur. 2024, 3, 100274. [Google Scholar] [CrossRef]
- Shimada, H.; Lu, Q.; Insinna-Kettenhofen, C.; Nagashima, K.; English, M.A.; Semler, E.M.; Mahgerefteh, J.; Cideciyan, A.V.; Li, T.; Brooks, B.P.; et al. In Vitro Modeling Using Ciliopathy-Patient-Derived Cells Reveals Distinct Cilia Dysfunctions Caused by CEP290 Mutations. Cell Rep. 2017, 20, 384–396. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Tan, C.; Wang, F.; Wang, Y.; Zhou, R.; Cui, D.; You, W.; Zhao, H.; Ren, J.; Feng, B. Knock-in of large reporter genes in human cells via CRISPR/Cas9-induced homology-dependent and independent DNA repair. Nucleic Acids Res. 2016, 44, e85. [Google Scholar] [CrossRef]
- Frigault, M.M.; Lacoste, J.; Swift, J.L.; Brown, C.M. Live-cell microscopy—Tips and tools. J. Cell Sci. 2009, 122, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.; Ansari, S.; Gebauer, L.; Müller, S.F.; Lowjaga, K.; Geyer, J.; Tzvetkov, M.V.; Brockmöller, J. A double-Flp-in method for stable overexpression of two genes. Sci. Rep. 2020, 10, 14018. [Google Scholar] [CrossRef]
- Lü, Q.; Zhang, C.; Westlake, C.J. Live-cell fluorescence imaging of ciliary dynamics. Biophys. Rep. 2021, 7, 101–110. [Google Scholar] [CrossRef]
- Yoon, J.; Comerci, C.J.; Weiss, L.E.; Milenkovic, L.; Stearns, T.; Moerner, W.E. Revealing Nanoscale Morphology of the Primary Cilium Using Super-Resolution Fluorescence Microscopy. Biophys. J. 2019, 116, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, W.; Xu, Z. [Comparison and progress review of various super-resolution fluorescence imaging techniques]. Se Pu 2021, 39, 1055–1064. [Google Scholar] [CrossRef]
- Sherman, S.; Nachmias, D.; Elia, N. A simple, straightforward correlative live-cell-imaging-structured-illumination-microscopy approach for studying organelle dynamics. Microsc. Res. Tech. 2015, 78, 777–783. [Google Scholar] [CrossRef]
- Al Jord, A.; Lemaître, A.I.; Delgehyr, N.; Faucourt, M.; Spassky, N.; Meunier, A. Centriole amplification by mother and daughter centrioles differs in multiciliated cells. Nature 2014, 516, 104–107. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, Y.; Wei, Q.; Huang, Y.; Hu, J.; Ling, K. Phosphatidylinositol phosphate kinase PIPKIγ and phosphatase INPP5E coordinate initiation of ciliogenesis. Nat. Commun. 2016, 7, 10777. [Google Scholar] [CrossRef]
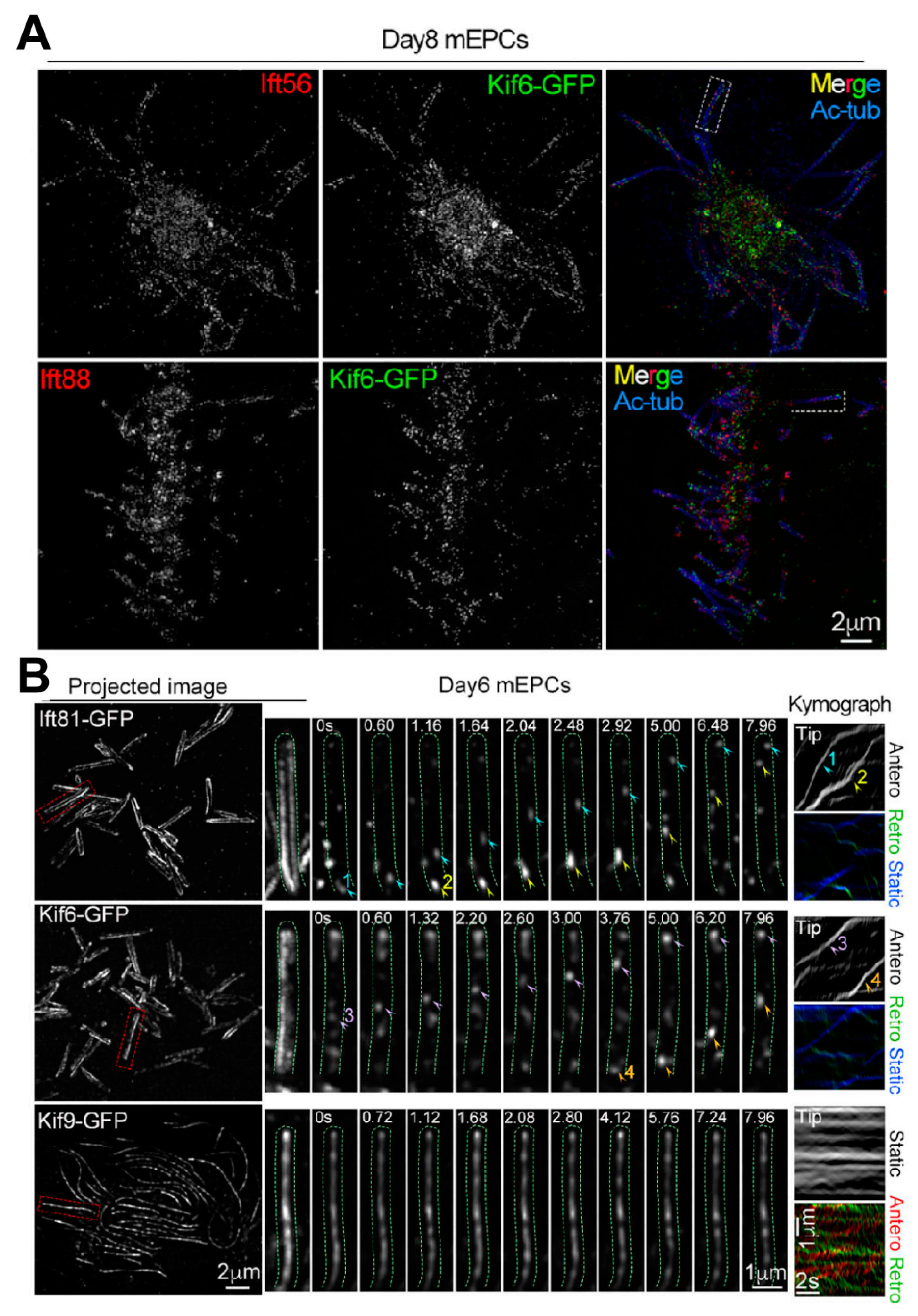
- Fang, C.; Pan, X.; Li, D.; Chen, W.; Huang, Y.; Chen, Y.; Li, L.; Gao, Q.; Liang, X.; Li, D.; et al. Distinct roles of Kif6 and Kif9 in mammalian ciliary trafficking and motility. J. Cell Biol. 2024, 223, e202312060. [Google Scholar] [CrossRef]
- Lewczuk, B.; Szyryńska, N. Field-Emission Scanning Electron Microscope as a Tool for Large-Area and Large-Volume Ultrastructural Studies. Animals 2021, 11, 3390. [Google Scholar] [CrossRef] [PubMed]
- Sjostrand, F.S. The ultrastructure of the innersegments of the retinal rods of the guinea pig eye as revealed by electron microscopy. J. Cell Comp. Physiol. 1953, 42, 45–70. [Google Scholar] [CrossRef]
- Jakus, M.A.; Hall, C.E. Electron microscope observations of the trichocysts and cilia of Paramecium. Biol. Bull. 1946, 91, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W. The study of epithelial cilia and sperm flagella with the electron microscope. Trans. Am. Laryngol. Rhinol. Otol. Soc. 1954, 4, 318–330. [Google Scholar] [CrossRef]
- Satir, P. Studies on Cilia: The Fixation of the Metachronal Wave. J. Cell Biol. 1963, 18, 345–365. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.R. The submicroscopic morphology of protoplasm. Harvey Lect. 1955, 51, 175–228. [Google Scholar]
- Despotes, K.A.; Zariwala, M.A.; Davis, S.D.; Ferkol, T.W. Primary Ciliary Dyskinesia: A Clinical Review. Cells 2024, 13, 974. [Google Scholar] [CrossRef]
- Ishikawa, T. Cryo-electron tomography of motile cilia and flagella. Cilia 2015, 4, 3. [Google Scholar] [CrossRef]
- Oikonomou, C.M.; Jensen, G.J. Cellular Electron Cryotomography: Toward Structural Biology In Situ. Annu. Rev. Biochem. 2017, 86, 873–896. [Google Scholar] [CrossRef]
- Murata, K.; Wolf, M. Cryo-electron microscopy for structural analysis of dynamic biological macromolecules. Biochim. Biophys. Acta. Gen. Subj. 2018, 1862, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Leiro, R.; Scheres, S.H. Unravelling biological macromolecules with cryo-electron microscopy. Nature 2016, 537, 339–346. [Google Scholar] [CrossRef]
- Hughes, L.C.; Ralston, K.S.; Hill, K.L.; Zhou, Z.H. Three-dimensional structure of the Trypanosome flagellum suggests that the paraflagellar rod functions as a biomechanical spring. PLoS ONE 2012, 7, e25700. [Google Scholar] [CrossRef] [PubMed]
- Oda, T. Three-dimensional structural labeling microscopy of cilia and flagella. Microscopy 2017, 66, 234–244. [Google Scholar] [CrossRef]
- Nicastro, D.; Schwartz, C.; Pierson, J.; Gaudette, R.; Porter, M.E.; McIntosh, J.R. The molecular architecture of axonemes revealed by cryoelectron tomography. Science 2006, 313, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Walton, T.; Gui, M.; Velkova, S.; Fassad, M.R.; Hirst, R.A.; Haarman, E.; O’Callaghan, C.; Bottier, M.; Burgoyne, T.; Mitchison, H.M.; et al. Axonemal structures reveal mechanoregulatory and disease mechanisms. Nature 2023, 618, 625–633. [Google Scholar] [CrossRef]
- Song, K.; Awata, J.; Tritschler, D.; Bower, R.; Witman, G.B.; Porter, M.E.; Nicastro, D. In situ localization of N and C termini of subunits of the flagellar nexin-dynein regulatory complex (N-DRC) using SNAP tag and cryo-electron tomography. J. Biol. Chem. 2015, 290, 5341–5353. [Google Scholar] [CrossRef]
- Ma, M.; Stoyanova, M.; Rademacher, G.; Dutcher, S.K.; Brown, A.; Zhang, R. Structure of the Decorated Ciliary Doublet Microtubule. Cell 2019, 179, 909–922.e12. [Google Scholar] [CrossRef]
- Gui, M.; Farley, H.; Anujan, P.; Anderson, J.R.; Maxwell, D.W.; Whitchurch, J.B.; Botsch, J.J.; Qiu, T.; Meleppattu, S.; Singh, S.K.; et al. De novo identification of mammalian ciliary motility proteins using cryo-EM. Cell 2021, 184, 5791–5806.e19. [Google Scholar] [CrossRef]
- Gui, M.; Croft, J.T.; Zabeo, D.; Acharya, V.; Kollman, J.M.; Burgoyne, T.; Höög, J.L.; Brown, A. SPACA9 is a lumenal protein of human ciliary singlet and doublet microtubules. Proc. Natl. Acad. Sci. USA 2022, 119, e2207605119. [Google Scholar] [CrossRef]
- Sun, S.; Fisher, R.L.; Bowser, S.S.; Pentecost, B.T.; Sui, H. Three-dimensional architecture of epithelial primary cilia. Proc. Natl. Acad. Sci. USA 2019, 116, 9370–9379. [Google Scholar] [CrossRef] [PubMed]
- Han, B.G.; Avila-Sakar, A.; Remis, J.; Glaeser, R.M. Challenges in making ideal cryo-EM samples. Curr. Opin. Struct. Biol. 2023, 81, 102646. [Google Scholar] [CrossRef]
- Czymmek, K.J.; Belevich, I.; Bischof, J.; Mathur, A.; Collinson, L.; Jokitalo, E. Accelerating data sharing and reuse in volume electron microscopy. Nat. Cell Biol. 2024, 26, 498–503. [Google Scholar] [CrossRef]
- Xu, C.S.; Pang, S.; Shtengel, G.; Müller, A.; Ritter, A.T.; Hoffman, H.K.; Takemura, S.Y.; Lu, Z.; Pasolli, H.A.; Iyer, N.; et al. An open-access volume electron microscopy atlas of whole cells and tissues. Nature 2021, 599, 147–151. [Google Scholar] [CrossRef]
- Heinrich, L.; Bennett, D.; Ackerman, D.; Park, W.; Bogovic, J.; Eckstein, N.; Petruncio, A.; Clements, J.; Pang, S.; Xu, C.S.; et al. Whole-cell organelle segmentation in volume electron microscopy. Nature 2021, 599, 141–146. [Google Scholar] [CrossRef]
- Cocks, E.; Taggart, M.; Rind, F.C.; White, K. A guide to analysis and reconstruction of serial block face scanning electron microscopy data. J. Microsc. 2018, 270, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qiao, F.; Guo, J.; Jiang, T.; Lou, H.; Li, H.; Xie, G.; Wu, H.; Wang, W.; Pei, R.; et al. In situ architecture of the intercellular organelle reservoir between epididymal epithelial cells by volume electron microscopy. Nat. Commun. 2025, 16, 1664. [Google Scholar] [CrossRef] [PubMed]
- Peddie, C.J.; Genoud, C.; Kreshuk, A.; Meechan, K.; Micheva, K.D.; Narayan, K.; Pape, C.; Parton, R.G.; Schieber, N.L.; Schwab, Y.; et al. Volume electron microscopy. Nat. Rev. Methods Primers 2022, 2, 51. [Google Scholar] [CrossRef]
- Eisenstein, M. Seven technologies to watch in 2023. Nature 2023, 613, 794–797. [Google Scholar] [CrossRef]
- Ding, W.; Wu, Q.; Sun, L.; Pan, N.C.; Wang, X. Cenpj Regulates Cilia Disassembly and Neurogenesis in the Developing Mouse Cortex. J. Neurosci. 2019, 39, 1994–2010. [Google Scholar] [CrossRef]
- Sheu, S.H.; Upadhyayula, S.; Dupuy, V.; Pang, S.; Deng, F.; Wan, J.; Walpita, D.; Pasolli, H.A.; Houser, J.; Sanchez-Martinez, S.; et al. A serotonergic axon-cilium synapse drives nuclear signaling to alter chromatin accessibility. Cell 2022, 185, 3390–3407.e18. [Google Scholar] [CrossRef] [PubMed]
- Polino, A.J.; Sviben, S.; Melena, I.; Piston, D.W.; Hughes, J. Scanning electron microscopy of human islet cilia. bioRxiv 2023. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Huang, S.; Chen, H. Advances in Imaging Techniques for Mammalian/Human Ciliated Cell’s Cilia: Insights into Structure, Function, and Dynamics. Biology 2025, 14, 521. https://doi.org/10.3390/biology14050521
Li J, Huang S, Chen H. Advances in Imaging Techniques for Mammalian/Human Ciliated Cell’s Cilia: Insights into Structure, Function, and Dynamics. Biology. 2025; 14(5):521. https://doi.org/10.3390/biology14050521
Chicago/Turabian StyleLi, Jin, Shiqin Huang, and Hao Chen. 2025. "Advances in Imaging Techniques for Mammalian/Human Ciliated Cell’s Cilia: Insights into Structure, Function, and Dynamics" Biology 14, no. 5: 521. https://doi.org/10.3390/biology14050521
APA StyleLi, J., Huang, S., & Chen, H. (2025). Advances in Imaging Techniques for Mammalian/Human Ciliated Cell’s Cilia: Insights into Structure, Function, and Dynamics. Biology, 14(5), 521. https://doi.org/10.3390/biology14050521