

The Identification of a Glucuronyltransferase-Related Gene, GlcAT-S, with Putative Mucus Protection and Anti-Inflammatory Effects from Gut-Damaged Drosophila by Dextran Sulfate Sodium (DSS)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Drosophila and Feeding Assay
2.2. cDNA Synthesis and Quantitative PCR Analysis
2.3. EC-Specific Knockdown of Selected Genes Using RNA Interference
2.4. Drosophila Gut Immunostaining and Imaging
2.5. Statistical Analysis
3. Results
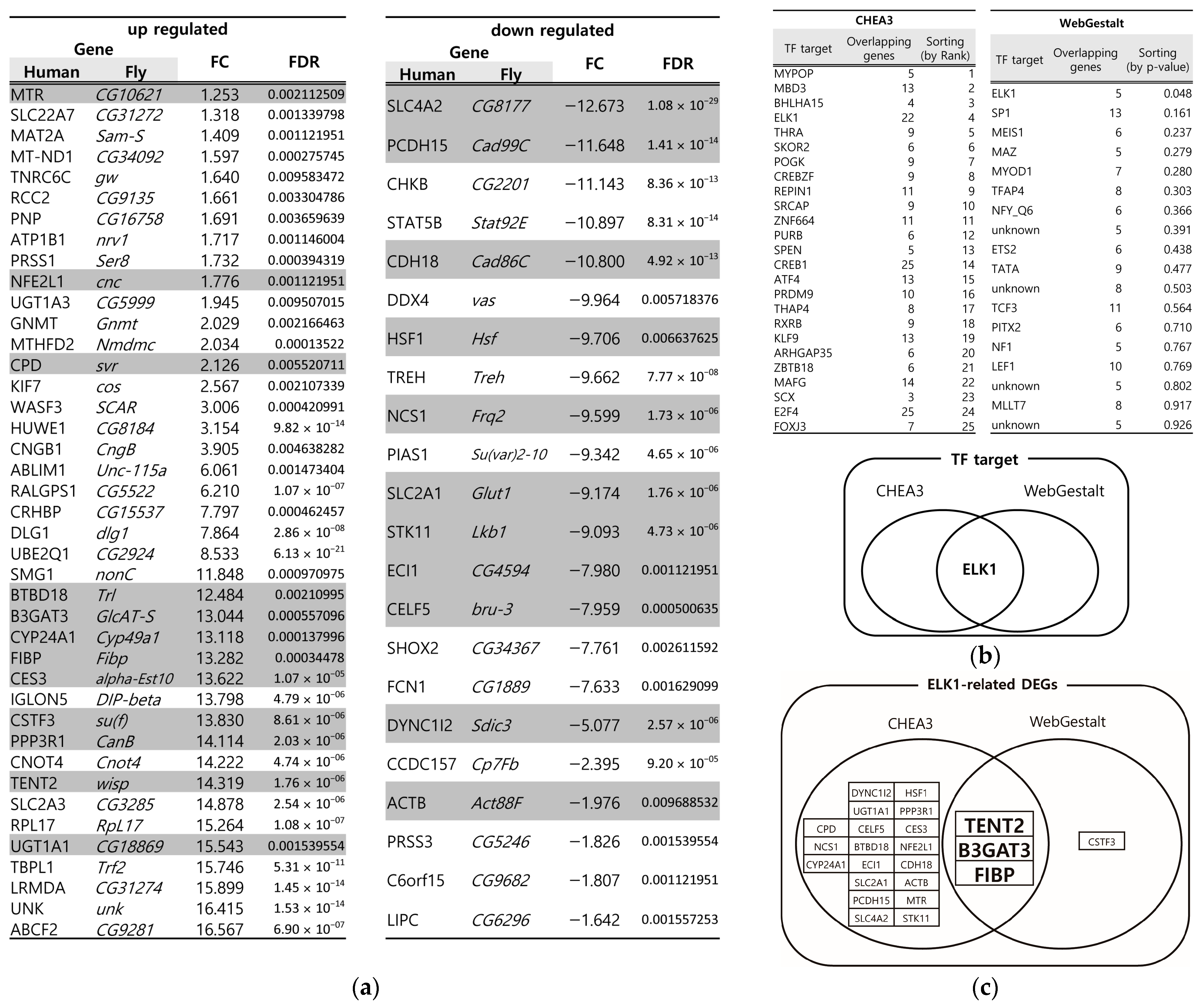
3.1. Identification of Key DEGs from the DSS-Induced Drosophila IBD Model Through Multiple Bioinformatics Tools
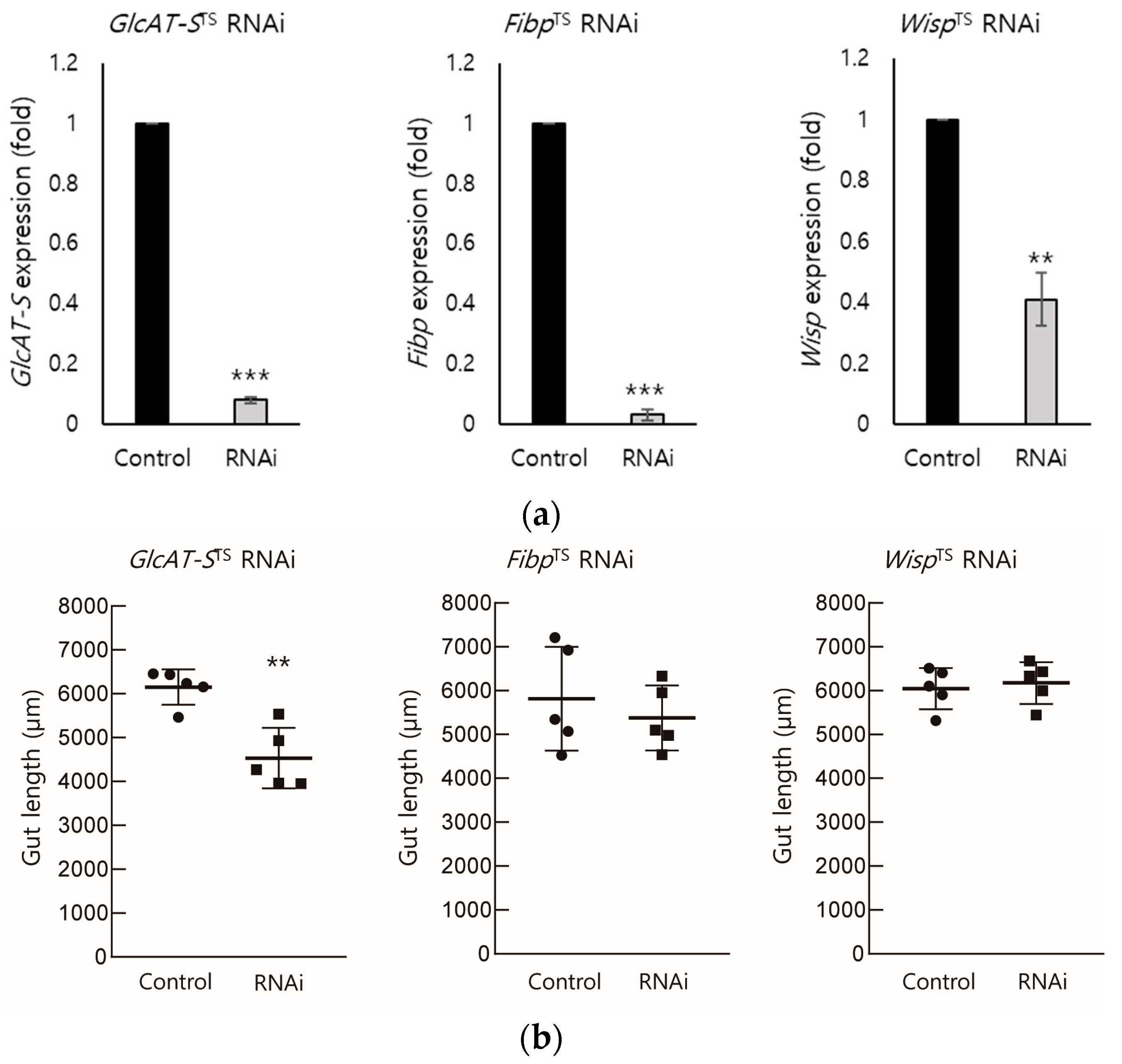
3.2. EC-Specific Knockdown of Screened DEGs
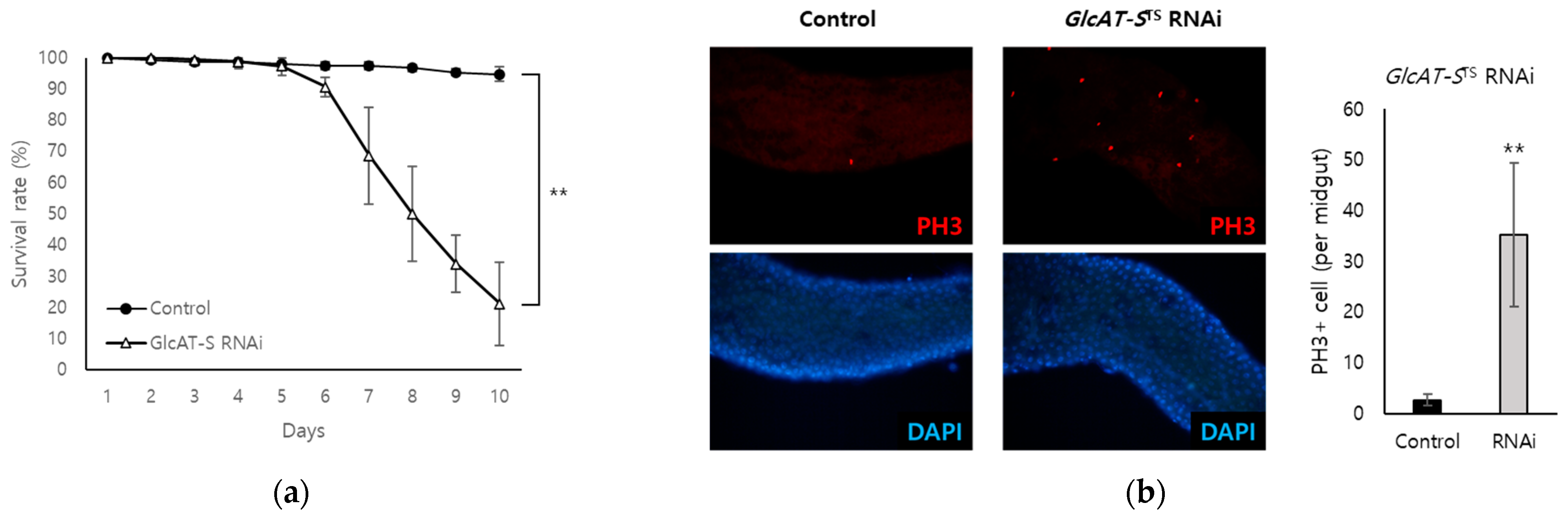
3.3. Confirmation of the Relationship Between GlcAT-S and Gut Tissue Damage Through EC-Specific Knockdown
3.4. Analysis of the Relationship Among GlcAT-S, Mucus Loss, and Inflammation in the Drosophila Gut-Damage Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neurath, M.F. Cytokines in Inflammatory Bowel Disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal Mucosal Barrier Function in Health and Disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Grondin, J.A.; Kwon, Y.H.; Far, P.M.; Haq, S.; Khan, W.I. Mucins in Intestinal Mucosal Defense and Inflammation: Learning from Clinical and Experimental Studies. Front. Immunol. 2020, 11, 2054. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N. Environmental Risk Factors for Inflammatory Bowel Disease. Gastroenterol. Hepatol. 2013, 9, 367–374. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C.D. Human Disease Models in Drosophila melanogaster and the Role of the Fly in Therapeutic Drug Discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef]
- Apidianakis, Y.; Rahme, L.G. Drosophila melanogaster as a Model for Human Intestinal Infection and Pathology. Dis. Models Mech. 2011, 4, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Galeone, A.; Han, S.Y.; Story, B.A.; Consonni, G.; Mueller, W.F.; Steinmetz, L.M.; Vaccari, T.; Jafar-Nejad, H. Gut Barrier Defects, Intestinal Immune Hyperactivation and Enhanced Lipid Catabolism Drive Lethality in NGLY1-Deficient Drosophila. Nat. Commun. 2023, 14, 5667. [Google Scholar] [CrossRef] [PubMed]
- Roditi, I.; Lehane, M.J. Interactions between Trypanosomes and Tsetse Flies. Curr. Opin. Microbiol. 2008, 11, 345–351. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. CP Immunol. 2014, 104, 15–25. [Google Scholar] [CrossRef]
- Katsandegwaza, B.; Horsnell, W.; Smith, K. Inflammatory Bowel Disease: A Review of Pre-Clinical Murine Models of Human Disease. Int. J. Mol. Sci. 2022, 23, 9344. [Google Scholar] [CrossRef]
- Zeng, C.; Liu, F.; Huang, Y.; Liang, Q.; He, X.; Li, L.; Xie, Y. Drosophila: An Important Model for Exploring the Pathways of Inflammatory Bowel Disease (IBD) in the Intestinal Tract. Int. J. Mol. Sci. 2024, 25, 12742. [Google Scholar] [CrossRef] [PubMed]
- Capo, F.; Wilson, A.; Di Cara, F. The Intestine of Drosophila Melanogaster: An Emerging Versatile Model System to Study Intestinal Epithelial Homeostasis and Host-Microbial Interactions in Humans. Microorganisms 2019, 7, 336. [Google Scholar] [CrossRef]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut Homeostasis in a Microbial World: Insights from Drosophila Melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef]
- Dietzl, G.; Chen, D.; Schnorrer, F.; Su, K.-C.; Barinova, Y.; Fellner, M.; Gasser, B.; Kinsey, K.; Oppel, S.; Scheiblauer, S.; et al. A Genome-Wide Transgenic RNAi Library for Conditional Gene Inactivation in Drosophila. Nature 2007, 448, 151–156. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A Revolutionary Tool for Transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Tarca, A.L.; Carey, V.J.; Chen, X.; Romero, R.; Drăghici, S. Machine Learning and Its Applications to Biology. PLoS Comput. Biol. 2007, 3, e116. [Google Scholar] [CrossRef]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, Transcriptome and Proteome: The Rise of Omics Data and Their Integration in Biomedical Sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Palmer, W.H.; Medd, N.C.; Beard, P.M.; Obbard, D.J. Isolation of a Natural DNA Virus of Drosophila Melanogaster, and Characterisation of Host Resistance and Immune Responses. PLoS Pathog. 2018, 14, e1007050. [Google Scholar] [CrossRef]
- Ewen-Campen, B.; Luan, H.; Xu, J.; Singh, R.; Joshi, N.; Thakkar, T.; Berger, B.; White, B.H.; Perrimon, N. Split-Intein Gal4 Provides Intersectional Genetic Labeling That Is Repressible by Gal80. Proc. Natl. Acad. Sci. USA 2023, 120, e2304730120. [Google Scholar] [CrossRef]
- Lee, S.H.; Hwang, D.; Goo, T.-W.; Yun, E.-Y. Prediction of Intestinal Stem Cell Regulatory Genes from Drosophila Gut Damage Model Created Using Multiple Inducers: Differential Gene Expression-Based Protein-Protein Interaction Network Analysis. Dev. Comp. Immunol. 2023, 138, 104539. [Google Scholar] [CrossRef]
- Park, D.; Singh, R.; Baym, M.; Liao, C.-S.; Berger, B. IsoBase: A Database of Functionally Related Proteins across PPI Networks. Nucleic Acids Res. 2011, 39, D295–D300. [Google Scholar] [CrossRef] [PubMed]
- Keenan, A.B.; Torre, D.; Lachmann, A.; Leong, A.K.; Wojciechowicz, M.L.; Utti, V.; Jagodnik, K.M.; Kropiwnicki, E.; Wang, Z.; Ma’ayan, A. ChEA3: Transcription Factor Enrichment Analysis by Orthogonal Omics Integration. Nucleic Acids Res. 2019, 47, W212–W224. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene Set Analysis Toolkit with Revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed]
- Osumi, R.; Sugihara, K.; Yoshimoto, M.; Tokumura, K.; Tanaka, Y.; Hinoi, E. Role of Proteoglycan Synthesis Genes in Osteosarcoma Stem Cells. Front. Oncol. 2024, 14, 1325794. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Niu, W.-B.; Hu, R.; Wang, L.-J.; Huang, Z.-Y.; Ni, S.-H.; Wang, M.-Q.; Yang, Y.; Huang, Y.-S.; Feng, W.-J.; et al. FIBP Knockdown Attenuates Growth and Enhances Chemotherapy in Colorectal Cancer via Regulating GSK3β-Related Pathways. Oncogenesis 2018, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Bofill-De Ros, X.; Stanton, R.; Shao, T.-J.; Villanueva, P.; Gu, S. TENT2, TUT4, and TUT7 Selectively Regulate miRNA Sequence and Abundance. Nat. Commun. 2022, 13, 5260. [Google Scholar] [CrossRef]
- Kong, S.; Zhang, Y.H.; Zhang, W. Regulation of Intestinal Epithelial Cells Properties and Functions by Amino Acids. BioMed Res. Int. 2018, 2018, 2819154. [Google Scholar] [CrossRef]
- Frazer, L.C.; Good, M. Intestinal Epithelium in Early Life. Mucosal Immunol. 2022, 15, 1181–1187. [Google Scholar] [CrossRef]
- Weaver, L.T.; Austin, S.; Cole, T.J. Small Intestinal Length: A Factor Essential for Gut Adaptation. Gut 1991, 32, 1321–1323. [Google Scholar] [CrossRef]
- Viragova, S.; Li, D.; Klein, O.D. Activation of Fetal-like Molecular Programs during Regeneration in the Intestine and Beyond. Cell Stem Cell 2024, 31, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Amcheslavsky, A.; Lindblad, J.L.; Bergmann, A. Transiently “Undead” Enterocytes Mediate Homeostatic Tissue Turnover in the Adult Drosophila Midgut. Cell Rep. 2020, 33, 108408. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Oz, H.S.; Yeh, S.-L.; Neuman, M.G. Gastrointestinal Inflammation and Repair: Role of Microbiome, Infection, and Nutrition. Gastroenterol. Res. Pract. 2016, 2016, 6516708. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and Regulation of the Mucus Barrier in the Gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef]
- Saez, A.; Herrero-Fernandez, B.; Gomez-Bris, R.; Sánchez-Martinez, H.; Gonzalez-Granado, J.M. Pathophysiology of Inflammatory Bowel Disease: Innate Immune System. Int. J. Mol. Sci. 2023, 24, 1526. [Google Scholar] [CrossRef]
- Sun, J.; Shen, X.; Li, Y.; Guo, Z.; Zhu, W.; Zuo, L.; Zhao, J.; Gu, L.; Gong, J.; Li, J. Therapeutic Potential to Modify the Mucus Barrier in Inflammatory Bowel Disease. Nutrients 2016, 8, 44. [Google Scholar] [CrossRef]
- Rawat, P.S.; Seyed Hameed, A.S.; Meng, X.; Liu, W. Utilization of Glycosaminoglycans by the Human Gut Microbiota: Participating Bacteria and Their Enzymatic Machineries. Gut Microbes 2022, 14, 2068367. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, S.-H.; Yang, J.-Y. Mechanobiological Approach for Intestinal Mucosal Immunology. Biology 2025, 14, 110. [Google Scholar] [CrossRef]
- Murch, S.H.; MacDonald, T.T.; Walker-Smith, J.A.; Lionetti, P.; Levin, M.; Klein, N.J. Disruption of Sulphated Glycosaminoglycans in Intestinal Inflammation. Lancet 1993, 341, 711–714. [Google Scholar] [CrossRef]
- Kim, K.; Lane, E.A.; Saftien, A.; Wang, H.; Xu, Y.; Wirtz-Peitz, F.; Perrimon, N. Drosophila as a Model for Studying Cystic Fibrosis Pathophysiology of the Gastrointestinal System. Proc. Natl. Acad. Sci. USA 2020, 117, 10357–10367. [Google Scholar] [CrossRef]
- Kitagawa, H.; Uyama, T.; Sugahara, K. Molecular Cloning and Expression of a Human Chondroitin Synthase. J. Biol. Chem. 2001, 276, 38721–38726. [Google Scholar] [CrossRef]
- Pompili, S.; Latella, G.; Gaudio, E.; Sferra, R.; Vetuschi, A. The Charming World of the Extracellular Matrix: A Dynamic and Protective Network of the Intestinal Wall. Front. Med. 2021, 8, 610189. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Hansson, G.C. Immunological Aspects of Intestinal Mucus and Mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Corfield, A.P. Mucins: A Biologically Relevant Glycan Barrier in Mucosal Protection. BBA Gen. Subj. 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Dasgupta, S.; Silva, J.; Wang, G.; Yu, R.K. Sulfoglucuronosyl Paragloboside Is a Ligand for T Cell Adhesion: Regulation of Sulfoglucuronosyl Paragloboside Expression via Nuclear Factor κB Signaling. J. Neurosci. Res. 2009, 87, 3591–3599. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Wang, B.; Yue, T.; Yun, E.-Y.; Ip, Y.T.; Jiang, J. Hippo Signaling Regulates Drosophila Intestine Stem Cell Proliferation through Multiple Pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 21064–21069. [Google Scholar] [CrossRef]
- Noah, T.K.; Donahue, B.; Shroyer, N.F. Intestinal Development and Differentiation. Exp. Cell Res. 2011, 317, 2702–2710. [Google Scholar] [CrossRef]
- Watnick, P.I.; Jugder, B.-E. Microbial Control of Intestinal Homeostasis via Enteroendocrine Cell Innate Immune Signaling. Trends Microbiol. 2020, 28, 141–149. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bergström, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.H.; Schütte, A.; van der Post, S.; Svensson, F.; Rodríguez-Piñeiro, A.M.; Nyström, E.E.L.; et al. The Mucus and Mucins of the Goblet Cells and Enterocytes Provide the First Defense Line of the Gastrointestinal Tract and Interact with the Immune System. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef]
- Hung, R.-J.; Hu, Y.; Kirchner, R.; Liu, Y.; Xu, C.; Comjean, A.; Tattikota, S.G.; Li, F.; Song, W.; Ho Sui, S.; et al. A Cell Atlas of the Adult Drosophila Midgut. Proc. Natl. Acad. Sci. USA 2020, 117, 1514–1523. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.H.; Hwang, D.; Lee, J.-W.; Goo, T.-W.; Yun, E.-Y. The Identification of a Glucuronyltransferase-Related Gene, GlcAT-S, with Putative Mucus Protection and Anti-Inflammatory Effects from Gut-Damaged Drosophila by Dextran Sulfate Sodium (DSS). Biology 2025, 14, 513. https://doi.org/10.3390/biology14050513
Lee SH, Hwang D, Lee J-W, Goo T-W, Yun E-Y. The Identification of a Glucuronyltransferase-Related Gene, GlcAT-S, with Putative Mucus Protection and Anti-Inflammatory Effects from Gut-Damaged Drosophila by Dextran Sulfate Sodium (DSS). Biology. 2025; 14(5):513. https://doi.org/10.3390/biology14050513
Chicago/Turabian StyleLee, Seung Hun, Dooseon Hwang, Jang-Won Lee, Tae-Won Goo, and Eun-Young Yun. 2025. "The Identification of a Glucuronyltransferase-Related Gene, GlcAT-S, with Putative Mucus Protection and Anti-Inflammatory Effects from Gut-Damaged Drosophila by Dextran Sulfate Sodium (DSS)" Biology 14, no. 5: 513. https://doi.org/10.3390/biology14050513
APA StyleLee, S. H., Hwang, D., Lee, J.-W., Goo, T.-W., & Yun, E.-Y. (2025). The Identification of a Glucuronyltransferase-Related Gene, GlcAT-S, with Putative Mucus Protection and Anti-Inflammatory Effects from Gut-Damaged Drosophila by Dextran Sulfate Sodium (DSS). Biology, 14(5), 513. https://doi.org/10.3390/biology14050513