
Antiepileptic Effects of Acorus tatarinowii Schott in a Rat Model of Epilepsy: Regulation of Metabolic Axes and Gut Microbiota

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Drugs
2.2. Experimental Design
2.3. Sample Collections and Preparation
2.4. Metabolomics Analysis
2.5. 16S rRNA Gene Sequencing Analysis
2.6. Statistical Analysis
3. Results
3.1. Therapeutic Efect of ATS on PTZ-Induced Epilepsy
3.2. Brain Metabolomics Analysis
3.3. Screen and Identification of Potential Biomarkers
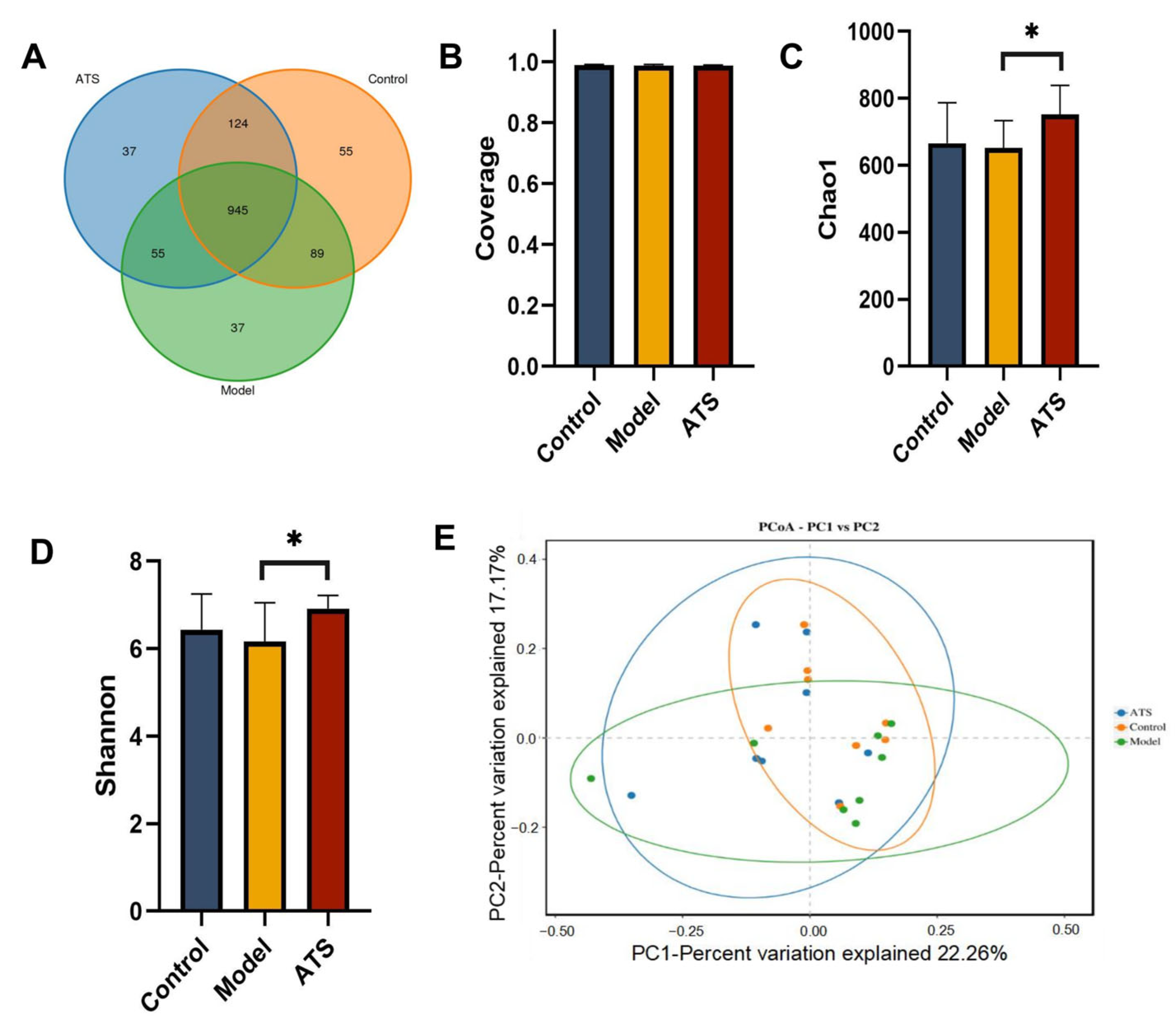
3.4. The Operating Unit (OTU) of Gut Microbiota Classification and Diversity Analysis
3.5. Composition and Structure Analysis of Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SV | Sodium valproate |
| PTZ | Pentylenetetrazol |
| ATS | Acorus tatarinowii Schott |
| PCA | Principal component analysis |
| OPLS-DA | Orthogonal projections to latent structures-discriminant analysis |
References
- Lu, Y.; Zhan, R.; Song, B.; Zhou, Y.; Zhu, L.; Chen, H.; Chen, X. The optimized biocatalytic synthesis of (S)-methyl 2-chlorobutanoate by Acinetobacter sp. lipase. Chirality 2022, 34, 1228–1238. [Google Scholar] [CrossRef]
- Manole, A.M.; Sirbu, C.A.; Mititelu, M.R.; Vasiliu, O.; Lorusso, L.; Sirbu, O.M.; Ionita Radu, F. State of the Art and Challenges in Epilepsy-A Narrative Review. J. Pers. Med. 2023, 13, 623. [Google Scholar] [CrossRef]
- Zhou, L.; Gao, Y.; Lu, H.; Liu, W.; Xu, X.; Xing, B.; Liang, X.; Wang, N.; Jiang, X.; Zhao, Q. Notopterygium incisum Root Extract (NRE) Alleviates Epileptiform Symptoms in PTZ-Induced Acute Seizure Mice. CNS Neurol. Disord. Drug Targets 2023, 22, 774–783. [Google Scholar]
- Pong, A.W.; Xu, K.J.; Klein, P. Recent advances in pharmacotherapy for epilepsy. Curr. Opin. Neurol. 2023, 36, 77–85. [Google Scholar] [CrossRef]
- Pan, I.; LoPresti, M.A.; Clarke, D.F.; Lam, S. The effectiveness of medical and surgical treatment for children with refractory epilepsy. Neurosurgery 2020, 88, 73–82. [Google Scholar] [CrossRef]
- López González, F.J.; Rodríguez Osorio, X.; Gil-Nagel Rein, A.; Carreño Martínez, M.; Serratosa Fernández, J.; Villanueva Haba, V.; Donaire Pedraza, A.J.; Mercadé Cerdá, J.M. Drug-resistant epilepsy: Definition and treatment alternatives. Neurologia 2015, 30, 439–446. [Google Scholar] [CrossRef]
- Harward, S.C.; Rolston, J.D.; Englot, D.J. Seizures in meningioma. Handb. Clin. Neurol. 2020, 170, 187–200. [Google Scholar]
- Maguire, M.; Singh, J.; Marson, A. Epilepsy and psychosis: A practical approach. Pract. Neurol. 2018, 18, 106–114. [Google Scholar] [CrossRef]
- Lin, C.H.; Hsieh, C.L. Chinese herbal medicine for treating epilepsy. Front. Neurosci. 2021, 15, 682821. [Google Scholar] [CrossRef]
- Wang, M.; Tang, H.P.; Wang, S.; Hu, W.J.; Li, J.Y.; Yu, A.Q.; Bai, Q.X.; Yang, B.Y.; Kuang, H.X. Acorus tatarinowii Schott: A review of its botany, traditional uses, phytochemistry, and pharmacology. Molecules 2023, 28, 4525. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, X.; Hong, X.; Wang, S.; Wei, J.; Huang, J.; Ji, L.; Yang, Y.; Efferth, T.; Hong, C.; et al. Essential oil of Acorus tatarinowii Schott inhibits neuroinflammation by suppressing NLRP3 inflammasome activation in 3×Tg-AD transgenic mice. Phytomedicine 2023, 112, 154695. [Google Scholar] [CrossRef]
- Lee, Y.C.; Kao, S.T.; Cheng, C.Y. Acorus tatarinowii Schott extract reduces cerebral edema caused by ischemia-reperfusion injury in rats: Involvement in regulation of astrocytic NKCC1/AQP4 and JNK/iNOS-mediated signaling. BMC Complement. Med. Ther. 2020, 20, 374. [Google Scholar] [CrossRef]
- Umeda, T.; Sakai, A.; Shigemori, K.; Nakata, K.; Nakajima, R.; Yamana, K.; Tomiyama, T. New value of Acorus tatarinowii/gramineus leaves as a dietary source for dementia prevention. Nutrients 2024, 16, 1589. [Google Scholar] [CrossRef]
- Kim, C.J.; Kwak, T.Y.; Bae, M.H.; Shin, H.K.; Choi, B.T. Therapeutic potential of active components from Acorus gramineus and Acorus tatarinowii in neurological disorders and their application in Korean medicine. J. Pharmacopunct. 2022, 25, 326–343. [Google Scholar] [CrossRef]
- Cheng, G.; Wang, X.; Wang, C.; Zhang, Q.; Zhang, Y. Understanding the molecular mechanisms of Acori Tatarinowii Rhizoma: Nardostahyos Radix et Rhizoma in epilepsy treatment using network pharmacology and molecular docking. Medicine 2024, 103, e37224. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Lu, T.; Chen, W.; Yan, W.; Yuan, K.; Shi, L.; Liu, X.; Zhou, X.; Shi, J.; et al. The microbiota-gut-brain axis in sleep disorders. Sleep. Med. Rev. 2022, 65, 101691. [Google Scholar] [CrossRef]
- Yue, Q.; Cai, M.; Xiao, B.; Zhan, Q.; Zeng, C. The Microbiota-gut-brain axis and epilepsy. Cell. Mol. Neurobiol. 2022, 42, 439–453. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef]
- Wang, Y.; Zhuo, Z.; Wang, H. Epilepsy, gut microbiota, and circadian rhythm. Front. Neurol. 2023, 14, 1157358. [Google Scholar] [CrossRef]
- Russo, E. The gut microbiota as a biomarker in epilepsy. Neurobiol. Dis. 2022, 163, 105598. [Google Scholar] [CrossRef]
- Xu, S.; Li, X.; Zhang, S.; Qi, C.; Zhang, Z.; Ma, R.; Xiang, L.; Chen, L.; Zhu, Y.; Tang, C.; et al. Oxidative stress gene expression, DNA methylation, and gut microbiota interaction trigger Crohn’s disease: A multi-omics Mendelian randomization study. BMC Med. 2023, 21, 179. [Google Scholar] [CrossRef] [PubMed]
- Mossad, O.; Batut, B.; Yilmaz, B.; Dokalis, N.; Mezö, C.; Nent, E.; Nabavi, L.S.; Mayer, M.; Maron, F.J.M.; Buescher, J.M.; et al. Gut microbiota drives age-related oxidative stress and mitochondrial damage in microglia via the metabolite N6-carboxymethyllysine. Nat. Neurosci. 2022, 25, 295–305. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Q.; Liu, X. The microbiota-gut-brain axis and neurodevelopmental disorders. Protein Cell 2023, 14, 762–775. [Google Scholar] [CrossRef]
- Deng, Y.; Zhou, M.; Wang, J.; Yao, J.; Yu, J.; Liu, W.; Wu, L.; Wang, J.; Gao, R. Involvement of the microbiota-gut-brain axis in chronic restraint stress: Disturbances of the kynurenine metabolic pathway in both the gut and brain. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Liu, L.; Wu, Q.; Chen, Y.; Ren, H.; Zhang, Q.; Yang, H.; Zhang, W.; Ding, T.; Wang, S.; Zhang, Y.; et al. Gut microbiota in chronic pain: Novel insights into mechanisms and promising therapeutic strategies. Int. Immunopharmacol. 2023, 115, 109685. [Google Scholar] [CrossRef]
- Zhao, Z.; Ning, J.; Bao, X.Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef]
- Shearer, J.; Scantlebury, M.H.; Rho, J.M.; Tompkins, T.A.; Mu, C. Intermittent vs continuous ketogenic diet: Impact on seizures, gut microbiota, and mitochondrial metabolism. Epilepsia 2023, 64, 177–183. [Google Scholar] [CrossRef]
- Mu, C.; Nikpoor, N.; Tompkins, T.A.; Choudhary, A.; Wang, M.; Marks, W.N.; Rho, J.M.; Scantlebury, M.H.; Shearer, J. Targeted gut microbiota manipulation attenuates seizures in a model of infantile spasms syndrome. JCI Insight 2022, 7, e158521. [Google Scholar] [CrossRef]
- Xu, L.; Chang, C.; Jiang, P.; Wei, K.; Zhang, R.; Jin, Y.; Zhao, J.; Xu, L.; Shi, Y.; Guo, S.; et al. Metabolomics in rheumatoid arthritis: Advances and review. Front. Immunol. 2022, 13, 961708. [Google Scholar] [CrossRef]
- Lista, S.; González-Domínguez, R.; López-Ortiz, S.; González-Domínguez, Á.; Menéndez, H.; Martín-Hernández, J.; Lucia, A.; Emanuele, E.; Centonze, D.; Imbimbo, B.P.; et al. Integrative metabolomics science in Alzheimer’s disease: Relevance and future perspectives. Ageing Res. Rev. 2023, 89, 101987. [Google Scholar] [CrossRef]
- Solch, R.J.; Aigbogun, J.O.; Voyiadjis, A.G.; Talkington, G.M.; Darensbourg, R.M.; O’Connell, S.; Pickett, K.M.; Perez, S.R.; Maraganore, D.M. Mediterranean diet adherence, gut microbiota, and Alzheimer’s or Parkinson’s disease risk: A systematic review. J. Neurol. Sci. 2022, 434, 120166. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, E.; Trivedi, D.K.; Sarkar, D.; Walton-Doyle, C.; Milne, J.; Kunath, T.; Rijs, A.M.; de Bie, R.M.A.; Goodacre, R.; Silverdale, M.; et al. Metabolomics of sebum reveals lipid dysregulation in Parkinson’s disease. Nat. Commun. 2021, 12, 1592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, X.; Mo, X.; Xiao, R.; Cheng, Q.; Wang, H.; Liu, L.; Xie, P. Enterogenic metabolomics signatures of depression: What are the possibilities for the future. Expert. Rev. Proteom. 2023, 20, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Bastos de Araújo, D.; Gurgel do Amaral, A.L.; Maia da Fonseca, S.; Rodrigues de Souza, K.; Santos da Paz, A.P.; Jóia de Mello, V.; Barbosa, G.B.; Otake Hamoy, M.K.; Hamoy, M. Lippia origanoides essential oil possesses anticonvulsant effect in pentylenetetrazol-induced seizures in rats: A behavioral, electroencephalographic, and electromyographic study. Front. Pharmacol. 2023, 14, 1289336. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Fang, C.; Wang, J. Metabolomics-Based Study on the Anticonvulsant Mechanism of Acorus tatarinowii: GABA Transaminase Inhibition Alleviates PTZ-Induced Epilepsy in Rats. Metabolites 2025, 15, 175. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Li, Q.; Sun, Q. Hepatotoxicity Induced by Methyl Eugenol: Insights from Toxicokinetics, Metabolomics, and Gut Microbiota. Curr. Issues Mol. Biol. 2024, 46, 11314–11325. [Google Scholar] [CrossRef]
- Ma, C.; Bi, K.; Zhang, M.; Su, D.; Fan, X.; Ji, W.; Wang, C.; Chen, X. Toxicology effects of morning glory seed in rat: A metabonomic method for profiling of urine metabolic changes. J. Ethnopharmacol. 2010, 130, 134–142. [Google Scholar] [CrossRef]
- How, M.S.; Hamid, N.; Liu, Y.; Kantono, K.; Oey, I.; Wang, M. Using OPLS-DA to Fingerprint Key Free Amino and Fatty Acids in Understanding the Influence of High Pressure Processing in New Zealand Clams. Foods 2023, 12, 1162. [Google Scholar] [CrossRef]
- Belmonte-Sánchez, J.R.; Romero-González, R.; Arrebola, F.J.; Vidal, J.L.M.; Garrido Frenich, A. An Innovative Metabolomic Approach for Golden Rum Classification Combining Ultrahigh-Performance Liquid Chromatography-Orbitrap Mass Spectrometry and Chemometric Strategies. J. Agric. Food Chem. 2019, 67, 1302–1311. [Google Scholar] [CrossRef]
- Zheng, Q.; Bartow-McKenney, C.; Meisel, J.S.; Grice, E.A. HmmUFOtu: An HMM and phylogenetic placement based ultra-fast taxonomic assignment and OTU picking tool for microbiome amplicon sequencing studies. Genome Biol. 2018, 19, 82. [Google Scholar] [CrossRef]
- Wang, L.L.; Zhang, F.Y.; Dong, W.W.; Wang, C.-L.; Liang, X.-Y.; Suo, L.-L.; Cheng, J.; Zhang, M.; Guo, X.-S.; Jiang, P.-H.; et al. A novel approach for the forensic diagnosis of drowning by microbiological analysis with next-generation sequencing and unweighted UniFrac-based PCoA. Int. J. Leg. Med. 2020, 134, 2149–2159. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Sun, X.; Rajesh, K.; Chalasani, N.; Gelow, K.; Katz, B.; Shah, V.H.; Sanyal, A.J.; Smirnova, E. Effects of Rare Microbiome Taxa Filtering on Statistical Analysis. Front. Microbiol. 2021, 11, 607325. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Zhu, Y.Y.; Yi, X.; Zhou, Z.S.; He, Y.J.; Zhou, Y.; Qi, Z.H.; Jin, D.N.; Zhao, L.X.; Luo, X.D. Bioguided isolation, identification and activity evaluation of antifungal compounds from Acorus tatarinowii Schott. J. Ethnopharmacol. 2020, 261, 113119. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.H.; Wu, Y.W.; Hung, J.I.; Chen, M.C. Epigallocatechin gallate/L-ascorbic acid-loaded poly-γ-glutamate microneedles with antioxidant, anti-inflammatory, and immunomodulatory effects for the treatment of atopic dermatitis. Acta Biomater. 2021, 130, 223–233. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Zhao, T.; Gong, M.; Wang, X.; Zhang, Y.; Xu, L.; Li, W.; Li, Y.; Jia, J. The role of N-methyl-D-aspartate glutamate receptors in Alzheimer’s disease: From pathophysiology to therapeutic approaches. Prog. Neurobiol. 2023, 231, 102534. [Google Scholar] [CrossRef]
- Kovács, Z.; Skatchkov, S.N.; Szabó, Z.; Qahtan, S.; Méndez-González, M.P.; Malpica-Nieves, C.J.; Eaton, M.J.; Kardos, J.; Héja, L. Putrescine intensifies Glu/GABA exchange mechanism and promotes early termination of seizures. Int. J. Mol. Sci. 2022, 23, 8191. [Google Scholar] [CrossRef]
- Janicot, R.; Shao, L.R.; Stafstrom, C.E. 2-deoxyglucose and β-hydroxybutyrate fail to attenuate seizures in the betamethasone-NMDA model of infantile spasms. Epilepsia Open 2022, 7, 181–186. [Google Scholar] [CrossRef]
- Wanleenuwat, P.; Suntharampillai, N.; Iwanowski, P. Antibiotic-induced epileptic seizures: Mechanisms of action and clinical considerations. Seizure 2020, 81, 167–174. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, Y.; Song, J.; Zhu, M.; Zhu, G.; Cai, H.; Chen, C.; Jin, M.; Song, Y. Perturbation of arachidonic acid and glycerolipid metabolism promoted particulate matter-induced inflammatory responses in human bronchial epithelial cells. Ecotoxicol. Environ. Saf. 2023, 256, 114839. [Google Scholar] [CrossRef]
- Kuo, Y.M.; Lee, Y.H. Epoxyeicosatrienoic acids and soluble epoxide hydrolase in physiology and diseases of the central nervous system. Chin. J. Physiol. 2022, 65, 1–11. [Google Scholar] [CrossRef]
- Chen, Y.; Nagib, M.M.; Yasmen, N.; Sluter, M.N.; Littlejohn, T.L.; Yu, Y.; Jiang, J. Neuroinflammatory mediators in acquired epilepsy: An update. Inflamm. Res. 2023, 72, 683–701. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Q.; Wu, D.; Zhang, D.W.; Li, S.C.; Zhang, S.W.; Chen, X.; Li, W. The effect of a novel anticonvulsant chemical Q808 on gut microbiota and hippocampus neurotransmitters in pentylenetetrazole-induced seizures in rats. BMC Neurosci. 2022, 23, 7. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Mai, Y.; Hu, L.; Zheng, R.; Zheng, D.; Chen, W.; Cai, Y.; Wang, J. Altered gut microbiota in temporal lobe epilepsy with anxiety disorders. Front. Microbiol. 2023, 14, 1165787. [Google Scholar] [CrossRef]
- Chakraborty, P.; Gamage, H.K.A.H.; Laird, A.S. Butyrate as a potential therapeutic agent for neurodegenerative disorders. Neurochem. Int. 2024, 176, 105745. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | Metabolite | m/z | RT (min) | VIP | p-Value | Trend (Model vs. Control) | Trend (ATS vs. Model) |
|---|---|---|---|---|---|---|---|
| ESI− | Hydroxymethyl cimetidine | 289.0869 | 2.3422 | 1.6938 | 0.0452 | ↓# | ↑* |
| 6-Keto-prostaglandin F1alpha | 369.2295 | 4.7634 | 2.2995 | 0.0025 | ↑# | ↓* | |
| Glycerophospho-N-Palmitoyl Ethanolamine | 452.2784 | 8.0968 | 1.6705 | 0.0409 | ↓# | ↑* | |
| PE (18:1(11Z)/18:2 (9Z,12Z)) | 762.5077 | 9.6214 | 1.7988 | 0.0435 | ↑# | ↓* | |
| Tylosin | 950.4912 | 9.6714 | 2.0651 | 0.0065 | ↑# | ↓* | |
| Threonylglutamic acid | 247.0937 | 0.6684 | 1.8601 | 0.0222 | ↑# | ↓* | |
| Biochanin A | 319.0345 | 0.7034 | 1.7813 | 0.0326 | ↑# | ↓* | |
| 7,8-Dihydro-2′-deoxyguanosine | 290.0868 | 0.8105 | 1.7534 | 0.0388 | ↑# | ↓* | |
| ESI+ | Succinoadenosine | 384.1176 | 1.8647 | 1.7709 | 0.0375 | ↑# | ↓* |
| 5-Ethynyl-2′-deoxyuridine | 252.0749 | 1.8718 | 1.8585 | 0.0266 | ↑# | ↓* | |
| Morinidazole | 271.1426 | 1.9575 | 1.7562 | 0.0429 | ↑# | ↓* | |
| PC (20:0/18:3 (10,12,15)-OH(9)) | 810.6015 | 10.0276 | 1.8457 | 0.0337 | ↑# | ↓* | |
| PC (P-18:1 (11Z)/PGE1) | 824.5823 | 10.0347 | 1.7213 | 0.0315 | ↓# | ↑* | |
| PS (22:0/18:1 (12Z)-O(9S,10R)) | 898.5612 | 11.1239 | 2.0329 | 0.0085 | ↑# | ↓* | |
| (Carbamoylamino) (2R)-2,5-diaminopentanoate | 155.0936 | 2.2209 | 2.3355 | 0.0050 | ↑# | ↓* | |
| 2-((3-Aminopyridin-2-yl) methylene) hydrazinecarbothioamide | 413.1061 | 2.2209 | 1.7978 | 0.0473 | ↑# | ↓* | |
| Aminomethyl fluorescein | 362.1020 | 2.2637 | 1.9851 | 0.0222 | ↑# | ↓* | |
| Hyaluronan biosynthesis, precursor 1 | 344.0949 | 3.0539 | 1.8754 | 0.0358 | ↑# | ↓* | |
| N6-Cyclopentyladenosine | 336.1668 | 3.6527 | 1.8747 | 0.0184 | ↑# | ↓* | |
| cis-trans-Nepetalactol | 191.1087 | 4.2366 | 1.7594 | 0.0332 | ↓# | ↑* | |
| 3′-Keto-3′-deoxy-AMP | 387.0799 | 4.6784 | 1.7929 | 0.0311 | ↓# | ↑* | |
| Guanidinosuccinic acid | 389.0785 | 4.6784 | 1.8230 | 0.0264 | ↓# | ↑* | |
| N-Acetylbialaphos | 404.0991 | 6.5657 | 1.7645 | 0.0436 | ↑# | ↓* | |
| 1-acyl-PAF | 538.3528 | 8.5529 | 1.8027 | 0.0274 | ↓# | ↑* | |
| PC (20:1 (11Z)/20:4 (5Z,8Z,11Z,14Z)-OH (20)) | 816.5843 | 9.5429 | 1.8898 | 0.0271 | ↑# | ↓* | |
| PC (P-18:1 (11Z)/PGJ2) | 844.5398 | 9.9419 | 1.6700 | 0.0433 | ↑# | ↓* | |
| Homocysteine thiolactone | 100.0218 | 0.7177 | 1.9255 | 0.0240 | ↓# | ↑* | |
| Mercaptopropionylglycine | 146.0278 | 0.7177 | 1.9153 | 0.0210 | ↓# | ↑* | |
| Phosphodimethylethanolamine | 192.0340 | 0.7177 | 1.9254 | 0.0230 | ↓# | ↑* | |
| L-xylo-Hexulonolactone | 141.0195 | 0.7248 | 2.0583 | 0.0107 | ↓# | ↑* | |
| Allantoic acid | 199.0448 | 0.8533 | 1.8406 | 0.0236 | ↓# | ↑* | |
| bemegride | 155.0939 | 1.4086 | 2.1221 | 0.0120 | ↑# | ↓* | |
| 7-Mercaptoheptanoylthreonine | 264.1257 | 1.7648 | 2.0653 | 0.0069 | ↑# | ↓* | |
| Topotecan | 444.1588 | 1.7933 | 1.8863 | 0.0171 | ↑# | ↓* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Li, J.; Zhang, W.; Wang, J. Antiepileptic Effects of Acorus tatarinowii Schott in a Rat Model of Epilepsy: Regulation of Metabolic Axes and Gut Microbiota. Biology 2025, 14, 488. https://doi.org/10.3390/biology14050488
Chen L, Li J, Zhang W, Wang J. Antiepileptic Effects of Acorus tatarinowii Schott in a Rat Model of Epilepsy: Regulation of Metabolic Axes and Gut Microbiota. Biology. 2025; 14(5):488. https://doi.org/10.3390/biology14050488
Chicago/Turabian StyleChen, Liang, Jiaxin Li, Wenhui Zhang, and Jiepeng Wang. 2025. "Antiepileptic Effects of Acorus tatarinowii Schott in a Rat Model of Epilepsy: Regulation of Metabolic Axes and Gut Microbiota" Biology 14, no. 5: 488. https://doi.org/10.3390/biology14050488
APA StyleChen, L., Li, J., Zhang, W., & Wang, J. (2025). Antiepileptic Effects of Acorus tatarinowii Schott in a Rat Model of Epilepsy: Regulation of Metabolic Axes and Gut Microbiota. Biology, 14(5), 488. https://doi.org/10.3390/biology14050488