
Analysis of the Interrelation and Seasonal Variation Characteristics of the Spatial Niche of Dominant Fishery Species—A Case Study of the East China Sea

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source
2.1.1. Resource Survey Data
2.1.2. Environment Data
2.2. Analysis Methods
2.2.1. Screening of Major Fish Species by Season
2.2.2. Index Calculation Method
Boosted Regression Trees
Suitability Index
Habitat Suitability Index
Overlap Index
2.3. The Software Used
3. Results
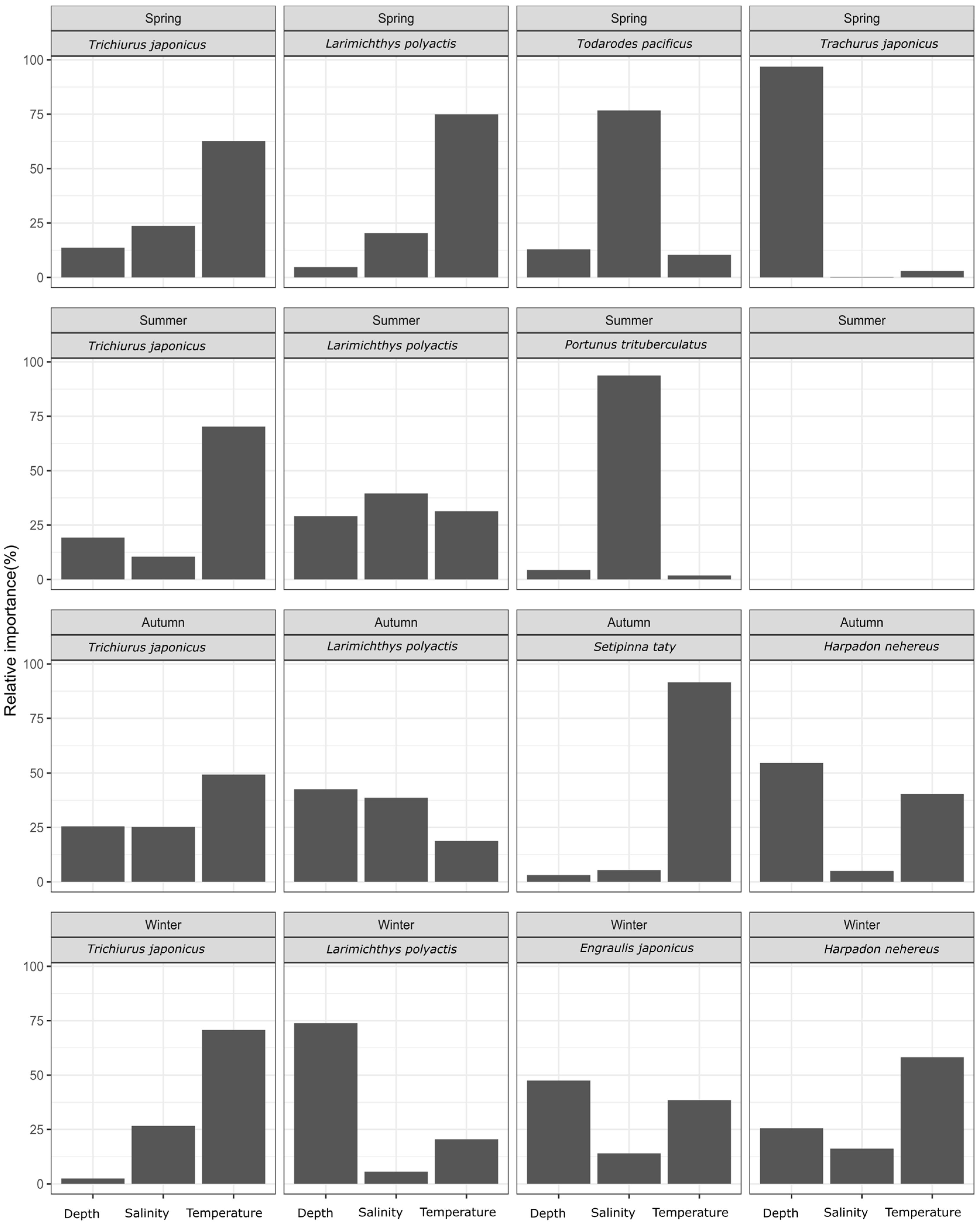
3.1. Relative Importance of Environmental Factors by Season for Each Fish Species
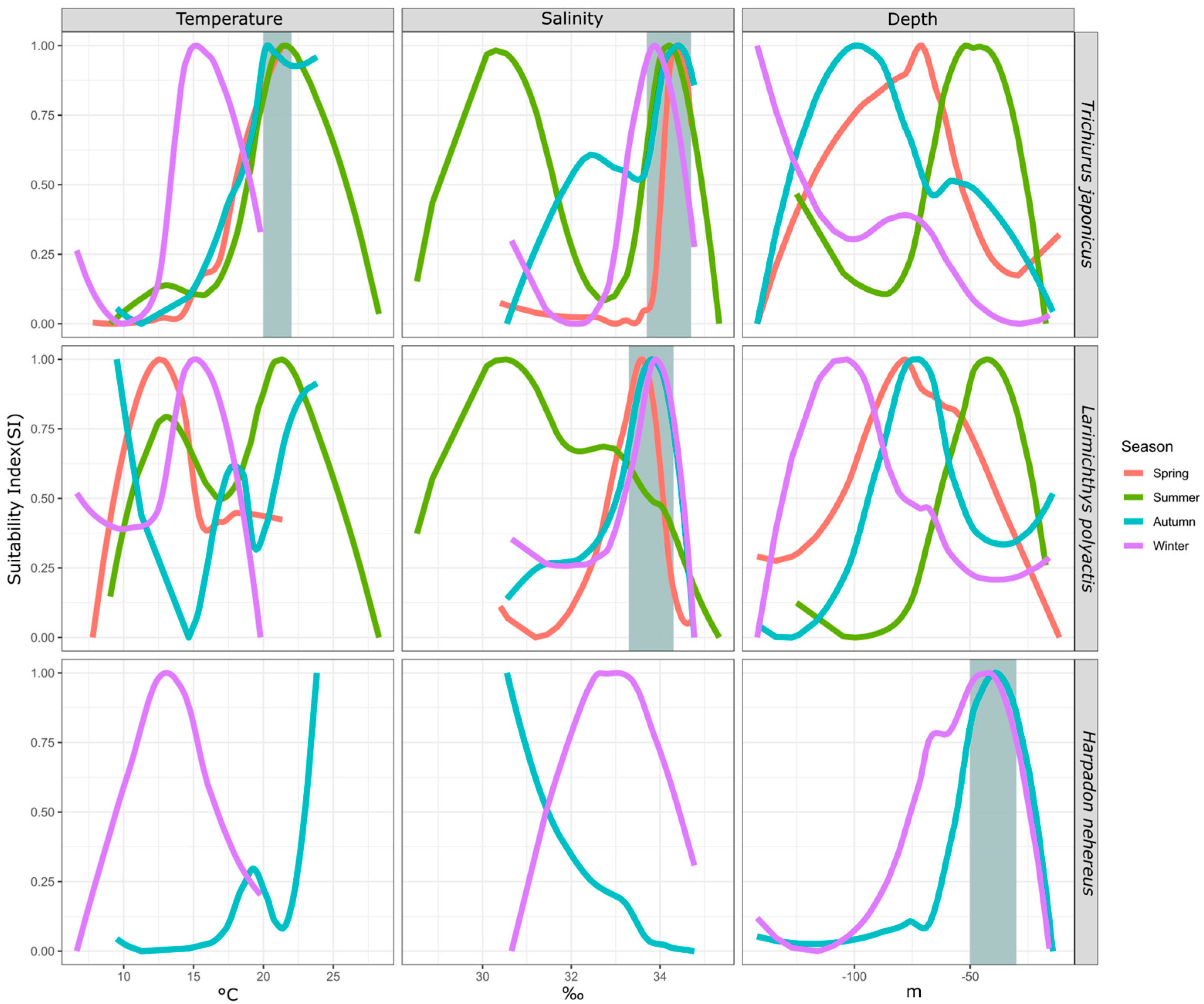
3.2. Smoothed Fitting Results of Environmental Factor SI
3.2.1. Differences in the Environmental Suitability Selection among Fish Species in Each Season
3.2.2. Seasonal Differences in the Environmental Suitability Selection for Each Fish Species
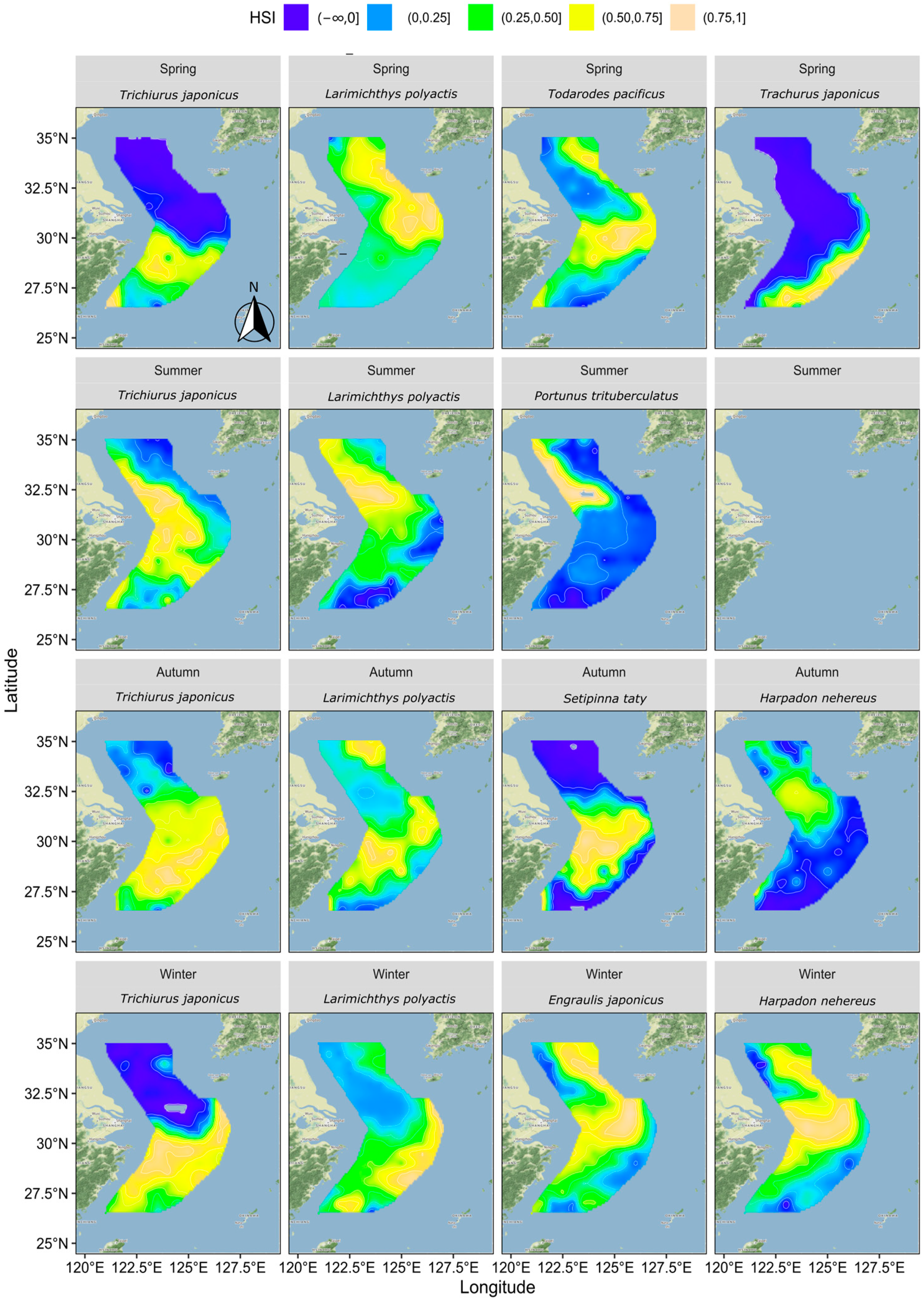
3.3. Distribution of HSI
3.4. Optimal Suitable Habitats Distribution Relationships
3.5. Comparison of Overlap Indices by Season
4. Discussion
4.1. Identification of Key Environmental Factors of the Species’ Habitats
4.2. Overlapping Ecological Niche Relationships between Species
4.3. A Comprehensive Inter-Seasonal Comparison of Overlapping Ecological Niche Relationships among Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pocheville, A. The ecological niche: History and recent controversies. In Handbook of Evolutionary Thinking in the Sciences; Springer: Dordrecht, The Netherlands, 2015; pp. 547–586. [Google Scholar]
- Encyclopedia B. Ecological Niche. 2021. Available online: https://baike.baidu.com/item/%E7%94%9F%E6%80%81%E4%BD%8D/1872121?fr=aladdin (accessed on 1 September 2024).
- Sun, R. General Ecology; Higher Education Press: Beijing, China, 1993. [Google Scholar]
- Odum, E.; Barrett, G. Fundermentals of Ecology, 5th ed.; Lu, J., Wang, W., Wang, T., He, W., Li, X., Eds.; Higher Education Press: Beijing, China, 2009. [Google Scholar]
- Liu, Y.; Cheng, J. Spato-temporal relationships among three groups of fishes caught in bottom trawl survey in the East China Sea. Acta Ecol. Sin. 2015, 35, 3403–3410. [Google Scholar]
- Lomolino, M.V.; Riddle, B.R.; Brown, J.H. Biogeography, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2009; p. 579. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bond, W.J. Alternative biome states challenge the modelling of species’ niche shifts under climate change. J. Ecol. 2021, 109, 3962–3971. [Google Scholar] [CrossRef]
- Lin, H.-C.; Tsai, C.-J.; Wang, H.-Y. Variation in global distribution, population structures, and demographic history for four Trichiurus cutlassfishes. PeerJ 2021, 9, e12639. [Google Scholar] [CrossRef] [PubMed]
- Bosso, L.; Panzuto, R.; Balestrieri, R.; Smeraldo, S.; Chiusano, M.L.; Raffini, F.; Canestrelli, D.; Musco, L.; Gili, C. Integrating citizen science and spatial ecology to inform management and conservation of the Italian seahorses. Ecol. Inform. 2024, 79, 102402. [Google Scholar] [CrossRef]
- Ge, X.; Griswold, C.K.; Newman, J.A. Robust species distribution predictions of predator and prey responses to climate change. J. Biogeogr. 2024, 51, 2047–2061. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Habitat Suitability Index (HSI). (2006–07). Available online: https://archive.epa.gov/aed/html/research/scallop/web/html/hsi.html (accessed on 1 September 2024).
- Di Febbraro, M.; Bosso, L.; Fasola, M.; Santicchia, F.; Aloise, G.; Lioy, S.; Tricarico, E.; Ruggieri, L.; Bovero, S.; Mori, E.; et al. Different facets of the same niche: Integrating citizen science and scientific survey data to predict biological invasion risk under multiple global change drivers. Glob. Chang. Biol. 2023, 29, 5509–5523. [Google Scholar] [CrossRef]
- Waheed, M.; Arshad, F.; Majeed, M.; Haq, S.M.; Aziz, R.; Bussmann, R.W.; Ali, K.; Subhan, F.; Jones, D.A.; Zaitouny, A. Potential distribution of a noxious weed (Solanum viarum Du-nal), current status, and future invasion risk based on MaxEnt modeling. Geol. Ecol. Landsc. 2023, 1–16. [Google Scholar] [CrossRef]
- Melo-Merino, S.M.; Reyes-Bonilla, H.; Lira-Noriega, A. Ecological niche models and species distribution models in marine environments: A literature review and spatial analysis of evidence. Ecol. Model. 2020, 415, 108837. [Google Scholar] [CrossRef]
- Xing, Q.; Yu, H.; Liu, Y.; Li, J.; Tian, Y.; Bakun, A.; Cao, C.; Tian, H.; Li, W. Application of a fish habitat model considering mesoscale oceanographic features in evaluating climatic impact on distribution and abundance of Pacific saury (Cololabis saira). Prog. Oceanogr. 2022, 201, 102743. [Google Scholar] [CrossRef]
- Feng, Z.; Yu, W.; Zhang, Y.; Li, Y.; Chen, X. Habitat variations of two commercially valuable species along the Chilean waters under different-intensity El Niño events. Front. Mar. Sci. 2022, 9, 946. [Google Scholar] [CrossRef]
- Fisheries Bureau of the Ministry of Agriculture. Animal Husbandry and Fisheries, East China Sea Fisheries Headquarters of Ministry of Agriculture, Animal Husbandry and Fisheries. Investigation and Division of Fishery Resources in the East China Sea; East China Normal University Press: Shanghai, China, 1987. [Google Scholar]
- Chen, D. Fisheries Resource Biology; China Agriculture Press: Beijing, China, 1997. [Google Scholar]
- Copernicus Marine Service. Global Ocean Physics Reanalysis, Sea Water Potential Temperature at Sea Floor & Sea Water Salinity, 0.083° × 0.083°, Daily. Available online: https://doi.org/10.48670/moi-00021 (accessed on 1 September 2024).
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. dismo: Species Distribution Modeling[M/OL]. R Package Version 1.3-3. 2020. Available online: https://CRAN.R-project.org/package=dismo (accessed on 1 September 2020).
- Chang, Y.J.; Sun, C.L.; Chen, Y.; Yeh, S.Z.; Dinardo, G. Habitat suitability analysis and identification of potential fishing grounds for swordfish, Xiphias gladius, in the South Atlantic Ocean. Int. J. Remote Sens. 2012, 33, 7523–7541. [Google Scholar] [CrossRef]
- Wakeley, J.S. A method to create simplified versions of existing habitat suitability index (HSI) models. Environ. Manag. 1988, 12, 79–83. [Google Scholar] [CrossRef]
- Vincenzi, S.; Caramori, G.; Rossi, R.; De Leo, G.A. A GIS-based habitat suitability model for commercial yield estimation of Tapes philippinarum in a Mediterranean coastal lagoon (Sacca di Goro, Italy). Ecol. Model. 2006, 193, 90–104. [Google Scholar] [CrossRef]
- Burman, P. A comparative study of ordinary cross-validation, v-fold cross-validation and the repeated learning-testing methods. Biometrika 1989, 76, 503–514. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 July 2021).
- Ripley, B.; Lapsley, M. RODBC: ODBC Database Access. Available online: https://CRAN.R-project.org/package=RODBC (accessed on 1 July 2021).
- Wickham, H.; Bryan, J. readxl: Read Excel Files. Available online: https://CRAN.R-project.org/package=readxl (accessed on 1 July 2021).
- Richard, A. BECKER O S Code by, RAY BROWNRIGG. A R W R Version by. Mapdata: Extra Map Databases. Available online: https://doi.org/10.32614/CRAN.package.mapdata (accessed on 1 July 2021).
- Richard, A. BECKER O S Code by, RAY BROWNRIGG. Enhancements by Thomas P minka A R W R Version by, DECKMYN. A. Maps: Draw Geographical Maps. Available online: https://CRAN.R-project.org/package=maps (accessed on 1 July 2021).
- Gerritsen, H. mapplots: Data Visualisation on Maps. Available online: https://CRAN.R-project.org/package=mapplots (accessed on 1 July 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 1 September 2024).
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, J.; Jia, S. Characteristics of submarine water temperature distribution of Trichiurus haumela in the East China Sea and Southern Yellow Sea with the improvement of the analysis methods. J. Fish. China 2019, 1749, 8. [Google Scholar]
- Lin, J. Trichiurus Haumela; Agriculture Press: Beijing, China, 1985. [Google Scholar]
- Mokhtari, M.; Ghaffar, M.A.; Usup, G.; Cob, Z.C. Determination of key environmental factors responsible for distribution patterns of fiddler crabs in a tropical mangrove ecosystem. PLoS ONE 2015, 10, e0117467. [Google Scholar] [CrossRef]
- Lins, R.C.; Martinez, J.M.; Motta Marques, D.D.; Cirilo, J.A.; Medeiros PR, P.; Fragoso Júnior, C.R. A Multivariate analysis framework to detect key environmental factors affecting spatiotemporal variability of chlorophyll-a in a tropical productive estuarine-lagoon system. Remote Sens. 2018, 10, 853. [Google Scholar] [CrossRef]
- Diankha, O.; Ba, A.; Brehmer, P.; Brochier, T.; Sow, B.A.; Thiaw, M.; Gaye, A.T.; Ngom, F.; Demarcq, H. Contrasted optimal environmental windows for both sardinella species in Senegalese waters. Fish. Oceanogr. 2018, 27, 351–365. [Google Scholar] [CrossRef]
- Cresci, A.; Larsen, T.; Halvorsen, K.T.; Durif, C.M.; Bjelland, R.; Browman, H.I.; Skiftesvik, A.B. Goldsinny Wrasse (Ctenolabrus Rupestris) Have a Sex-Dependent Magnetic Compass for Maintaining Site Fidelity. Fish. Oceanogr. 2022, 31, 164–171. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, J. Analyzing the Combined Effect of Multiple Environmental Factors on Fish Distribution, by Means of the Mixed Distribution–Decomposition Approach, as Illustrated by the East China Sea Hairtail. Biology 2023, 12, 1009. [Google Scholar] [CrossRef] [PubMed]
- von Schuckmann, K.; Le Traon, P.-Y.; Smith, N.; Pascual, A.; Djavidnia, S.; Brasseur, P.; Grégoire, M. Copernicus Ocean state report, issue 6. J. Oper. Oceanogr. 2022, 15 (Suppl. 1), 1–220. [Google Scholar]
- Li, D.Z.; Shi, Q.A.; Zang, R.G.; Wang, X.P.; Sheng, L.J.; Zhu, Z.L.; Wang, C.A. Models for Niche Breadth and Niche Overlap of Species or Populations. Sci. Silvae Sin. 2006, 42, 95–103. [Google Scholar]
- Wei, S. Food and feeding habits of hairtail (Trichiurus haumela Forskal) from the Yellow Sea. Mar. Fish. Res. 1980, 1, 49–57. [Google Scholar]
- Liu, Y.; Cheng, J.; Chen, Y. A spatial analysis of trophic composition: A case study of hairtail (Trichiurus japonicus) in the East China Sea. Hydrobiologia 2009, 632, 79–90. [Google Scholar] [CrossRef]
- Zhang, B. Feeding habits and ontogenetic diet shift of hairtail fish (Trichiurus lepturus) in East China Sea and Yellow Sea. Mar. Fish. Res. 2004, 25, 6–12. [Google Scholar]
- Lin, L.S.; Yan, L.P.; Ling, J.Z.; Liu, Y.; Zhou, R. Food habits of hairtail in the East China Sea region. Mar. Fish. 2005, 27, 187–192. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Species | Biomass (g) | Proportion | Cumulative Proportion |
|---|---|---|---|---|
| Spring | Hairtail (Trichiurus japonicus) | 1,620,338 | 0.36 | 0.36 |
| Small Yellow Croaker (Larimichthys polyactis) | 566,553 | 0.13 | 0.49 | |
| Pacific Squid (Todarodes pacificus) | 297,404 | 0.07 | 0.56 | |
| Japanese Jack Mackerel (Trachurus japonicus) | 290,772 | 0.07 | 0.62 | |
| Summer | Hairtail | 6,074,370 | 0.41 | 0.41 |
| Small Yellow Croaker | 3,494,318 | 0.24 | 0.65 | |
| Swimming Crab (Portunus trituberculatus) | 907,367 | 0.06 | 0.71 | |
| Autumn | Hairtail | 893,098 | 0.23 | 0.23 |
| Small yellow Croaker | 412,755 | 0.11 | 0.33 | |
| Half-fin Anchovy (Setipinna taty) | 321,371 | 0.08 | 0.41 | |
| Bombay Duck (Harpadon nehereus) | 314,403 | 0.08 | 0.50 | |
| Winter | Small Yellow Croaker | 699,973 | 0.23 | 0.23 |
| Hairtail | 356,924 | 0.12 | 0.34 | |
| Anchovy (Engraulis japonicus) | 301,428 | 0.10 | 0.44 | |
| Bombay Duck | 228,832 | 0.07 | 0.52 |
| Season | Count Area | Overlap Index |
|---|---|---|
| Spring | 2163 | −0.1583 |
| Summer | 1554 | 0.1287 |
| Autumn | 1920 | 0.4612 |
| Winter | 2564 | 0.5187 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Cheng, J. Analysis of the Interrelation and Seasonal Variation Characteristics of the Spatial Niche of Dominant Fishery Species—A Case Study of the East China Sea. Biology 2024, 13, 751. https://doi.org/10.3390/biology13090751
Liu Y, Cheng J. Analysis of the Interrelation and Seasonal Variation Characteristics of the Spatial Niche of Dominant Fishery Species—A Case Study of the East China Sea. Biology. 2024; 13(9):751. https://doi.org/10.3390/biology13090751
Chicago/Turabian StyleLiu, Yong, and Jiahua Cheng. 2024. "Analysis of the Interrelation and Seasonal Variation Characteristics of the Spatial Niche of Dominant Fishery Species—A Case Study of the East China Sea" Biology 13, no. 9: 751. https://doi.org/10.3390/biology13090751
APA StyleLiu, Y., & Cheng, J. (2024). Analysis of the Interrelation and Seasonal Variation Characteristics of the Spatial Niche of Dominant Fishery Species—A Case Study of the East China Sea. Biology, 13(9), 751. https://doi.org/10.3390/biology13090751