The Protection and Management of Wapiti in Desert Oases: Bare Land Poses a Limitation to Wapiti Conservation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Method
2.1. Study Area
2.2. GPS Tracking Records
2.3. Construction of a Multi-Scale Environmental Covariate Set
2.4. Model Selection
2.5. Spatial Filtering
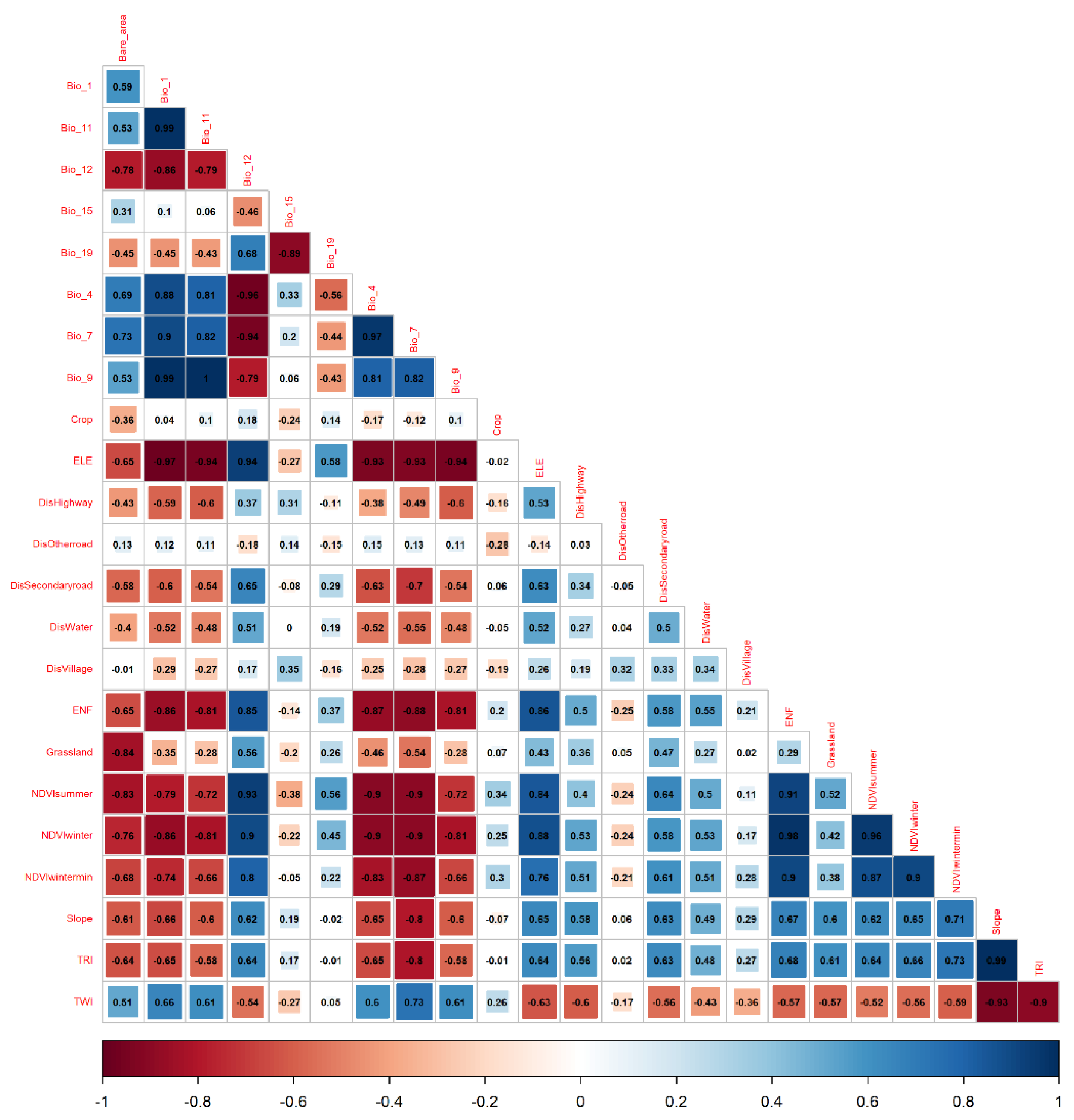
2.6. Variable Selection
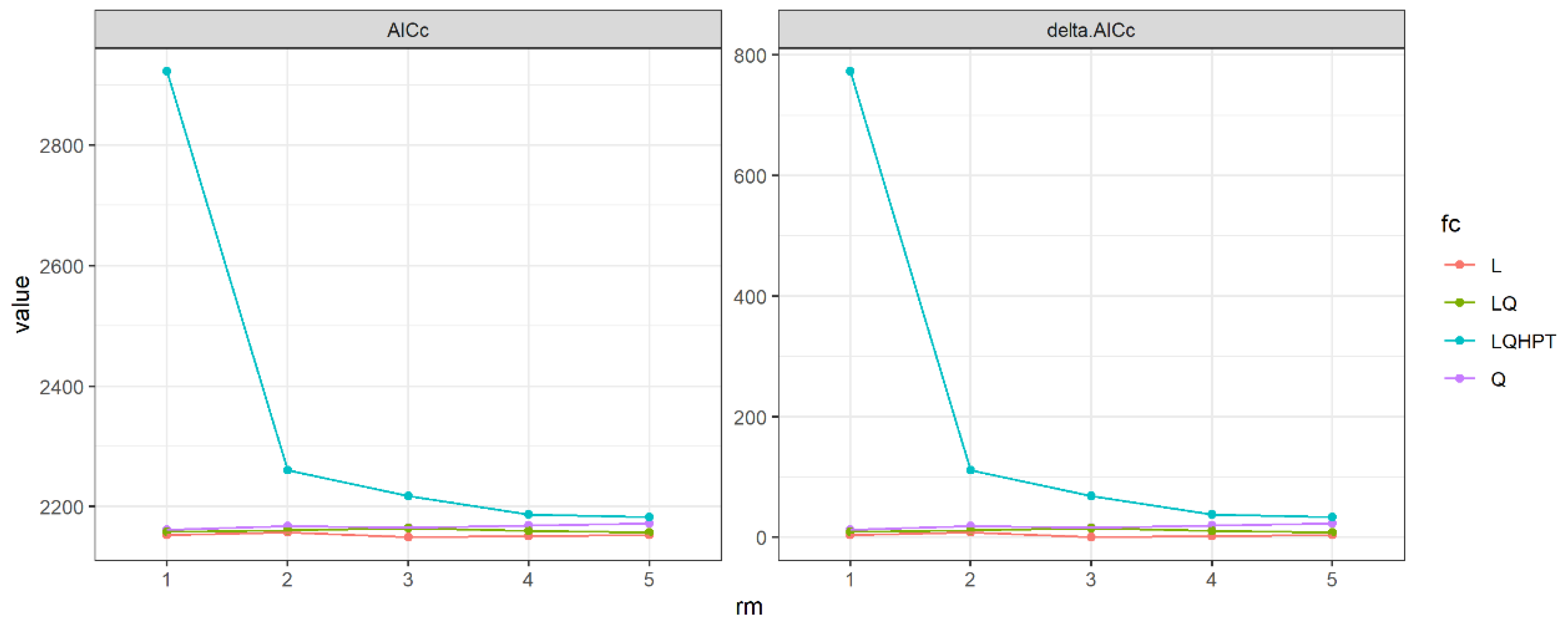
2.7. Model Optimization and Selection
2.8. Model Validation
3. Results
3.1. Determination of the Optimal Scales of Environmental Covariates
3.2. Reslts of Model Optimization and Selection
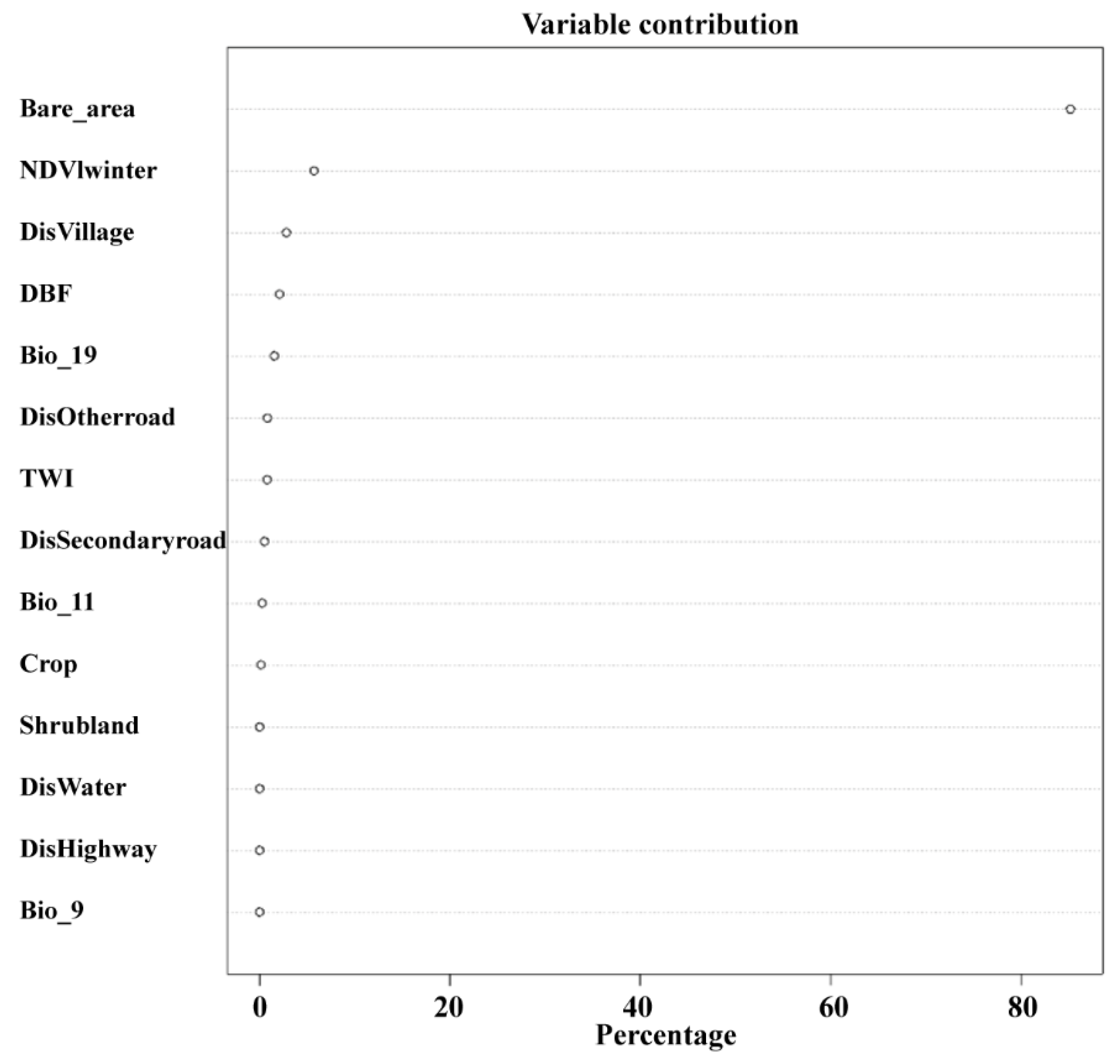
3.3. Habitat Selection Modeling Based on Environmental Covariates with Optimal Scales
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pringle, R.M.; Abraham, J.O.; Anderson, T.M.; Coverdale, T.C.; Davies, A.B.; Dutton, C.L.; Gaylard, A.; Goheen, J.R.; Holdo, R.M.; Hutchinson, M.C.; et al. Impacts of Large Herbivores on Terrestrial Ecosystems. Curr. Biol. 2023, 33, R584–R610. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Guinness, F.E.; Albon, S.D. Red Deer: Behavior and Ecology of Two Sexes; University of Chicago Press: Chicago, IL, USA, 1982; ISBN 978-0-226-11057-8. [Google Scholar]
- Rivrud, I.M.; Loe, L.E.; Mysterud, A. How Does Local Weather Predict Red Deer Home Range Size at Different Temporal Scales? J. Anim. Ecol. 2010, 79, 1280–1295. [Google Scholar] [CrossRef]
- Jerina, K. Roads and Supplemental Feeding Affect Home-Range Size of Slovenian Red Deer More than Natural Factors. J. Mammal. 2012, 93, 1139–1148. [Google Scholar] [CrossRef]
- Reinecke, H.; Leinen, L.; Thißen, I.; Meißner, M.; Herzog, S.; Schütz, S.; Kiffner, C. Home Range Size Estimates of Red Deer in Germany: Environmental, Individual and Methodological Correlates. Eur. J. Wildl. Res. 2014, 60, 237–247. [Google Scholar] [CrossRef]
- Sun, P.; Huang, S.; Su, Y.; Meng, D.; Zhang, Z.; Teng, L.; Liu, Z. Red deer population size and structure on Helan Mountain, Inner Mongolia, China. Chin. J. Wildl. 2021, 42, 341–347. [Google Scholar]
- Gaur, M.K.; Squires, V.R. Geographic Extent and Characteristics of the World’s Arid Zones and Their Peoples. In Climate Variability Impacts on Land Use and Livelihoods in Dylands; Gaur, M.K., Squires, V.R., Eds.; Springer: Cham, Switzerland, 2018; pp. 3–20. ISBN 978-3-319-56681-8. [Google Scholar]
- Luo, Y. Comparing the Diet and Habitat Selection of Sympatric Blue Sheep (Pseudois nayaur) and Reddeer (Cervus elaphus alxaicus) in Helan Mountains, China; Northeast Forestry University: Harbin, China, 2011. [Google Scholar]
- Zecherle, L.J.; Bar-David, S.; Nichols, H.J.; Templeton, A.R.; Hipperson, H.; Horsburgh, G.J.; Brown, R.P. Landscape Resistance Affects Individual Habitat Selection but Not Genetic Relatedness in a Reintroduced Desert Ungulate. Biol. Conserv. 2020, 252, 108845. [Google Scholar] [CrossRef]
- Gao, H.; Qiao, F.; Teng, L.; Junle, L.; Yu, M.; Liu, Z. Genetic Diversity and Structure of the Alashan Red Deer (Cervus elaphus alashanicus) in Helan Mountains, China. Acta Theriol. Sin. 2020, 40, 458–466. [Google Scholar]
- Templeton, A.R. Measuring Biodiversity and Monitoring Ecological and Evolutionary Processes with Genetic and Genomic Tools. In The Routledge Handbook of Philosophy of Biodiversity; Routledge: Oxfordshire, UK, 2016; ISBN 978-1-315-53021-5. [Google Scholar]
- Meserve, P.L.; Kelt, D.A.; Milstead, W.B.; GutiÉrrez, J.R. Thirteen Years of Shifting Top-Down and Bottom-Up Control. BioScience 2003, 53, 633–646. [Google Scholar] [CrossRef]
- Gao, H.; Huang, Y.; Sun, Y.; Qiao, F.; Yu, M.; Li, J.; Li, N.; Teng, L.; Liu, Z. Inferring Landscape Factors Driving Microgeographic Genetic Structure of Large-Sized Mountain Ungulates: A Case of Alashan Red Deer (Cervus elaphus alxaicus). Glob. Ecol. Conserv. 2023, 44, e02497. [Google Scholar] [CrossRef]
- Forbes, E.S.; Cushman, J.H.; Burkepile, D.E.; Young, T.P.; Klope, M.; Young, H.S. Synthesizing the Effects of Large, Wild Herbivore Exclusion on Ecosystem Function. Funct. Ecol. 2019, 33, 1597–1610. [Google Scholar] [CrossRef]
- Kerr, J.T.; Deguise, I. Habitat Loss and the Limits to Endangered Species Recovery. Ecol. Lett. 2004, 7, 1163–1169. [Google Scholar] [CrossRef]
- Kearney, M. Habitat, Environment and Niche: What Are We Modelling? Oikos 2006, 115, 186–191. [Google Scholar] [CrossRef]
- Orians, G.H.; Wittenberger, J.F. Spatial and Temporal Scales in Habitat Selection. Am. Nat. 1991, 137, S29–S49. [Google Scholar] [CrossRef]
- Coristine, L.E.; Kerr, J.T. Habitat Loss, Climate Change, and Emerging Conservation Challenges in Canada1This Review Is Part of the Virtual Symposium “Flagship Species – Flagship Problems” That Deals with Ecology, Biodiversity and Management Issues, and Climate Impacts on Species at Risk and of Canadian Importance, Including the Polar Bear (Ursus maritimus), Atlantic Cod (Gadus morhua), Piping Plover (Charadrius melodus), and Caribou (Rangifer tarandus). Can. J. Zool. 2011, 89, 435–451. [Google Scholar] [CrossRef]
- Manly, B.F.J.; McDonald, L.L.; Thomas, D.L.; McDonald, T.L.; Erickson, W.P. Introduction to Resource Selection Studies. In Resource Selection by Animals: Statistical Design and Analysis for Field Studies; Springer: Dordrecht, The Netherlands, 2002; pp. 1–15. ISBN 978-0-306-48151-2. [Google Scholar]
- McGarigal, K.; Zeller, K.A.; Cushman, S.A. Multi-Scale Habitat Selection Modeling: Introduction to the Special Issue. Landsc. Ecol. 2016, 31, 1157–1160. [Google Scholar] [CrossRef]
- Mayor, S.J.; Schneider, D.C.; Schaefer, J.A.; Mahoney, S.P. Habitat Selection at Multiple Scales. Écoscience 2009, 16, 238–247. [Google Scholar] [CrossRef]
- Lidicker, W.Z. Responses of Mammals to Habitat Edges: An Overview. Landsc. Ecol. 1999, 14, 333–343. [Google Scholar] [CrossRef]
- Uboni, A.; Smith, D.W.; Stahler, D.R.; Vucetich, J.A. Selecting Habitat to What Purpose? The Advantage of Exploring the Habitat–Fitness Relationship. Ecosphere 2017, 8, e01705. [Google Scholar] [CrossRef]
- McMahon, L.A.; Rachlow, J.L.; Shipley, L.A.; Forbey, J.S.; Johnson, T.R. Habitat Selection Differs across Hierarchical Behaviors: Selection of Patches and Intensity of Patch Use. Ecosphere 2017, 8, e01993. [Google Scholar] [CrossRef]
- Ciarniello, L.M.; Boyce, M.S.; Seip, D.R.; Heard, D.C. Grizzly Bear Habitat Selection Is Scale Dependent. Ecol. Appl. 2007, 17, 1424–1440. [Google Scholar] [CrossRef]
- Bauder, J.M.; Breininger, D.R.; Bolt, M.R.; Legare, M.L.; Jenkins, C.L.; Rothermel, B.B.; McGarigal, K. Multi-Level, Multi-Scale Habitat Selection by a Wide-Ranging, Federally Threatened Snake. Landsc. Ecol. 2018, 33, 743–763. [Google Scholar] [CrossRef]
- Bozek, C.K.; Prange, S.; Gehrt, S.D. The Influence of Anthropogenic Resources on Multi-Scale Habitat Selection by Raccoons. Urban Ecosyst. 2007, 10, 413–425. [Google Scholar] [CrossRef]
- Cushman, S.A.; McGarigal, K. Patterns in the Species–Environment Relationship Depend on Both Scale and Choice of Response Variables. Oikos 2004, 105, 117–124. [Google Scholar] [CrossRef]
- Martínez-Miranzo, B.; Banda, E.I.; Aguirre, J.I. Multiscale Analysis of Habitat Selection by Bonelli’s Eagle (Aquila fasciata) in NE Spain. Eur. J. Wildl. Res. 2016, 62, 673–679. [Google Scholar] [CrossRef]
- Salo, P.; Banks, P.B.; Dickman, C.R.; Korpimäki, E. Predator Manipulation Experiments: Impacts on Populations of Terrestrial Vertebrate Prey. Ecol. Monogr. 2010, 80, 531–546. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Z.; Gao, H.; Zhao, C.; Sun, Y.; Teng, L. Comparative Analysis of Winter Diets and Habitat Use by the Sympatric Blue Sheep (Pseudois nayaur) and Alashan Red Deer (Cervus alashanicus) in the Helan Mountains, China. Folia Zool. 2018, 67, 43–53. [Google Scholar] [CrossRef]
- Li, Z.; Wang, J.; Khattak, R.H.; Han, X.; Liu, P.; Liu, Z.; Teng, L. Coexistence Mechanisms of Sympatric Ungulates: Behavioral and Physiological Adaptations of Blue Sheep (Pseudois nayaur) and Red Deer (Cervus elaphus alxaicus) in Helan Mountains, China. Front. Ecol. Evol. 2022, 10, 925465. [Google Scholar] [CrossRef]
- Liu, F.; Huang, S.; Xie, J.; Liang, J.; Su, Y. Population Status and Protection of Blue Sheep in Helan Mountains of the Inner Mongolia, China. Acta Ecol. Sin. 2023, 43, 5829–5839. [Google Scholar]
- Ma, T.; Wang, G.; Guo, R.; Chen, H.; Jia, N.; Ma, J.; Cheng, H.; Zhang, Y. Multi-Scale Habitat Selection Modeling Using Combinatorial Optimization of Environmental Covariates: A Case Study on Nature Reserve of Red-Crowned Cranes. Ecol. Indic. 2023, 154, 110488. [Google Scholar] [CrossRef]
- Bojarska, K.; Kurek, K.; Śnieżko, S.; Wierzbowska, I.; Król, W.; Zyśk-Gorczyńska, E.; Baś, G.; Widera, E.; Okarma, H. Winter Severity and Anthropogenic Factors Affect Spatial Behaviour of Red Deer in the Carpathians. Mammal Res. 2020, 65, 815–823. [Google Scholar] [CrossRef]
- Jarnemo, A.; Nilsson, L.; Wikenros, C. Home Range Sizes of Red Deer in Relation to Habitat Composition: A Review and Implications for Management in Sweden. Eur. J. Wildl. Res. 2023, 69, 92. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive Habitat Distribution Models in Ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Austin, M.P. Spatial Prediction of Species Distribution: An Interface between Ecological Theory and Statistical Modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Chivers, L.S.; Lundy, M.G.; Colhoun, K.; Newton, S.F.; Houghton, J.D.R.; Reid, N. Identifying Optimal Feeding Habitat and Proposed Marine Protected Areas (pMPAs) for the Black-Legged Kittiwake (Rissa tridactyla) Suggests a Need for Complementary Management Approaches. Biol. Conserv. 2013, 164, 73–81. [Google Scholar] [CrossRef]
- Na, X.; Zhou, H.; Zang, S.; Wu, C.; Li, W.; Li, M. Maximum Entropy Modeling for Habitat Suitability Assessment of Red-Crowned Crane. Ecol. Indic. 2018, 91, 439–446. [Google Scholar] [CrossRef]
- Zhang, W.; Li, X.; Yu, L.; Si, Y. Multi-Scale Habitat Selection by Two Declining East Asian Waterfowl Species at Their Core Spring Stopover Area. Ecol. Indic. 2018, 87, 127–135. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial Filtering to Reduce Sampling Bias Can Improve the Performance of Ecological Niche Models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander Jr, J.A. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making Better Maxent Models of Species Distributions: Complexity, Overfitting and Evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM Eval: An R Package for Conducting Spatially Independent Evaluations and Estimating Optimal Model Complexity for Maxent Ecological Niche Models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Zeller, K.A.; Vickers, T.W.; Ernest, H.B.; Boyce, W.M. Multi-Level, Multi-Scale Resource Selection Functions and Resistance Surfaces for Conservation Planning: Pumas as a Case Study. PLoS ONE 2017, 12, e0179570. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, W.; Bleisch, W.V.; Li, Q.; Wang, H.; Lu, W.; Sun, J.; Zhang, F.; Ti, B.; Jiang, X. Functional Diversity Loss and Change in Nocturnal Behavior of Mammals under Anthropogenic Disturbance. Conserv. Biol. 2022, 36, e13839. [Google Scholar] [CrossRef] [PubMed]
- Nickel, B.A.; Suraci, J.P.; Allen, M.L.; Wilmers, C.C. Human Presence and Human Footprint Have Non-Equivalent Effects on Wildlife Spatiotemporal Habitat Use. Biol. Conserv. 2020, 241, 108383. [Google Scholar] [CrossRef]
- Cimatti, M.; Ranc, N.; Benítez-López, A.; Maiorano, L.; Boitani, L.; Cagnacci, F.; Čengić, M.; Ciucci, P.; Huijbregts, M.A.; Krofel, M. Large Carnivore Expansion in Europe Is Associated with Human Population Density and Land Cover Changes. Divers. Distrib. 2021, 27, 602–617. [Google Scholar] [CrossRef]
- Suel, H. Brown Bear (Ursus arctos) Habitat Suitability Modelling and Mapping. Appl. Ecol. Environ. Res. 2019, 17, 4245–4255. [Google Scholar] [CrossRef]
- Parsons, B.M.; Wilson, A.E.; Graham, K.; Stenhouse, G.B. Grizzly Bear (Ursus arctos) Movements and Habitat Use Predict Human-Caused Mortality across Temporal Scales. Can. J. Zool. 2022, 101, 81–94. [Google Scholar] [CrossRef]
- Hubbard, T.; Cove, M.V.; Lafferty, D.J. Human Recreation Impacts Seasonal Activity and Occupancy of American Black Bears (Ursus americanus) across the Anthropogenic-Wildland Interface. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Hoeks, S.; Huijbregts, M.A.; Busana, M.; Harfoot, M.B.; Svenning, J.-C.; Santini, L. Mechanistic Insights into the Role of Large Carnivores for Ecosystem Structure and Functioning. Ecography 2020, 43, 1752–1763. [Google Scholar] [CrossRef]
- Marshal, J.P.; Krausman, P.R.; Bleich, V.C. Rainfall, Temperature, and Forage Dynamics Affect Nutritional Quality of Desert Mule Deer Forage. Rangel. Ecol. Manag. 2005, 58, 360–365. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, F.; Du, H.; Zhang, X.; Feng, C.; Tan, Z.; Yu, Y.; Liu, Z. The Protection and Management of Wapiti in Desert Oases: Bare Land Poses a Limitation to Wapiti Conservation. Biology 2024, 13, 737. https://doi.org/10.3390/biology13090737
Qiao F, Du H, Zhang X, Feng C, Tan Z, Yu Y, Liu Z. The Protection and Management of Wapiti in Desert Oases: Bare Land Poses a Limitation to Wapiti Conservation. Biology. 2024; 13(9):737. https://doi.org/10.3390/biology13090737
Chicago/Turabian StyleQiao, Fujie, Hairong Du, Xia Zhang, Caiping Feng, Zhihua Tan, Yanqin Yu, and Zhensheng Liu. 2024. "The Protection and Management of Wapiti in Desert Oases: Bare Land Poses a Limitation to Wapiti Conservation" Biology 13, no. 9: 737. https://doi.org/10.3390/biology13090737
APA StyleQiao, F., Du, H., Zhang, X., Feng, C., Tan, Z., Yu, Y., & Liu, Z. (2024). The Protection and Management of Wapiti in Desert Oases: Bare Land Poses a Limitation to Wapiti Conservation. Biology, 13(9), 737. https://doi.org/10.3390/biology13090737