
Nanovesicular Mediation of the Gut–Brain Axis by Probiotics: Insights into Irritable Bowel Syndrome


 , , , , , ,
, , , , , ,  ,
,  ,
,  , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Participant Recruitment
2.3. Plasmatic EV Purification and Characterization
2.3.1. DLS
2.3.2. STEM
2.3.3. Western Blot Analysis
2.4. In Vitro Study
2.4.1. Bacterial Strains and Culture Conditions
2.4.2. Cell Culture
2.4.3. Treatments of Cells
2.4.4. Protein Extraction, Quantification, and Western Blot Analysis
2.4.5. Immunofluorescence Staining and Confocal Microscopy
2.4.6. Isolation of Vesicles from Culture Medium
2.5. Statistical Analyses
3. Results
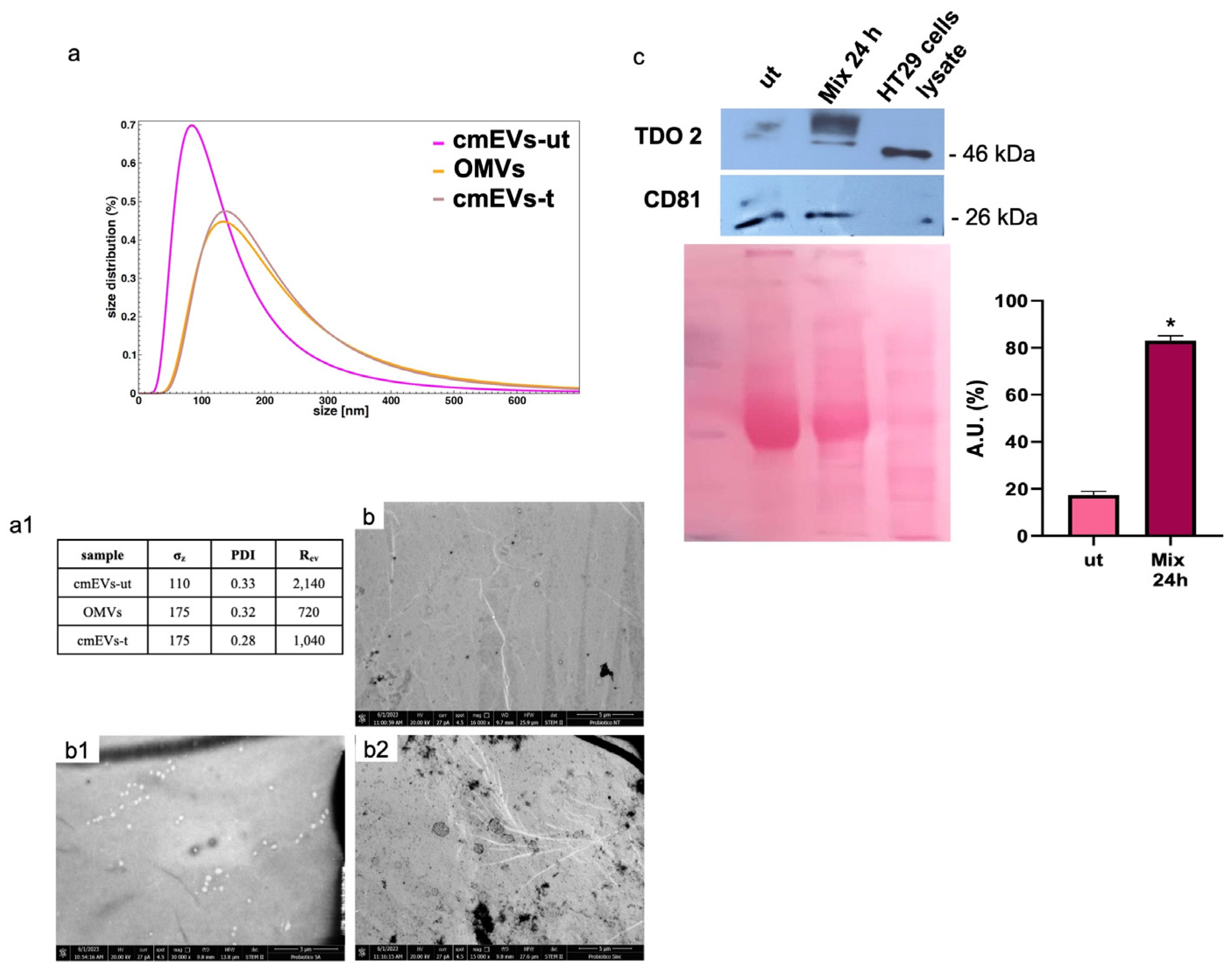
3.1. TDO 2 Is Secreted in Blood Circulation through EVs
3.2. The Probiotics Mix of LS33, LA14, and BGNA4 Strains Has Cytoprotective Properties on HT29 Cells
3.3. Anti-Stress Effect of Probiotics against Oxidative Stress Induced by H2O2 in HT29 Cells
3.4. Probiotics Treatment Increases TDO 2 and Hsp90 Protein Levels in HT29 Cells
3.5. The Probiotics Mix Modulates the Expression of Serotonin Receptors in HT29 Cells
3.6. Probiotics Treatment Increases TDO 2 and Hsp90 Protein Levels in EVs Isolated from HT29
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiegers, C.; van Beek, E.H.T.; Larsen, O.F.A. Clinical Research with Probiotics as an Indicator of Global Valorization since the Year 2000. Front. Microbiol. 2023, 14, 1323920. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.; Naveed, M.; Makhdoom, S.I.; Ali, U.; Mughal, M.S.; Sarwar, A.; Khan, A.A.; Zhennai, Y.; Sameeh, M.Y.; Dablool, A.S.; et al. Genome Investigation and Functional Annotation of Lactiplantibacillus Plantarum YW11 Revealing Streptin and Ruminococcin-A as Potent Nutritive Bacteriocins against Gut Symbiotic Pathogens. Molecules 2023, 28, 491. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.; Khan, A.A.; Tzora, A.; Voidarou, C.; Skoufos, I. Dietary Implications of the Bidirectional Relationship between the Gut Microflora and Inflammatory Diseases with Special Emphasis on Irritable Bowel Disease: Current and Future Perspective. Nutrients 2023, 15, 2956. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.; Hussain, N.; Hameed, Z.; Lin, L. Elucidating the Role of Diet in Maintaining Gut Health to Reduce the Risk of Obesity, Cardiovascular and Other Age-Related Inflammatory Diseases: Recent Challenges and Future Recommendations. Gut Microbes 2024, 16, 2297864. [Google Scholar] [CrossRef] [PubMed]
- Martyniak, A.; Medyńska-Przęczek, A.; Wędrychowicz, A.; Skoczeń, S.; Tomasik, P.J. Prebiotics, Probiotics, Synbiotics, Paraprobiotics and Postbiotic Compounds in IBD. Biomolecules 2021, 11, 1903. [Google Scholar] [CrossRef] [PubMed]
- Marasco, G.; Cremon, C.; Barbaro, M.R.; Stanghellini, V.; Barbara, G. Gut Microbiota Signatures and Modulation in Irritable Bowel Syndrome. Microbiome Res. Rep. 2022, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-Y.; Choi, S.-W. Dietary Modulation of Gut Microbiota for the Relief of Irritable Bowel Syndrome. Nutr. Res. Pract. 2021, 15, 411. [Google Scholar] [CrossRef] [PubMed]
- Vijay, A.; Valdes, A.M. Role of the Gut Microbiome in Chronic Diseases: A Narrative Review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Fucarino, A.; Burgio, S.; Paladino, L.; Caruso Bavisotto, C.; Pitruzzella, A.; Bucchieri, F.; Cappello, F. The Microbiota Is Not an Organ: Introducing the Muco-Microbiotic Layer as a Novel Morphofunctional Structure. Anatomia 2022, 1, 186–203. [Google Scholar] [CrossRef]
- Cappello, F.; Saguto, D.; Burgio, S.; Paladino, L.; Bucchieri, F. Does Intestine Morphology Still Have Secrets to Reveal? A Proposal about the “Ghost” Layer of the Bowel. Appl. Biosci. 2022, 1, 95–100. [Google Scholar] [CrossRef]
- Intili, G.; Paladino, L.; Rappa, F.; Alberti, G.; Plicato, A.; Calabrò, F.; Fucarino, A.; Cappello, F.; Bucchieri, F.; Tomasello, G.; et al. From Dysbiosis to Neurodegenerative Diseases through Different Communication Pathways: An Overview. Biology 2023, 12, 195. [Google Scholar] [CrossRef] [PubMed]
- Min, Z.; Xiaona, H.; Aziz, T.; Jian, Z.; Zhennai, Y. Exopolysaccharides from Lactobacillus Plantarum YW11 Improve Immune Response and Ameliorate Inflammatory Bowel Disease Symptoms. Acta Biochim. Pol. 2020, 67, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-L.; Chu, H.-F.; Wu, C.-C.; Deng, F.-S.; Wen, P.-J.; Chien, S.-P.; Chao, C.-H.; Chen, Y.-T.; Lu, M.-K.; Tsai, Y.-C. Exopolysaccharide Is the Potential Effector of Lactobacillus Fermentum PS150, a Hypnotic Psychobiotic Strain. Front. Microbiol. 2023, 14, 1209067. [Google Scholar] [CrossRef] [PubMed]
- Usuda, H.; Okamoto, T.; Wada, K. Leaky Gut: Effect of Dietary Fiber and Fats on Microbiome and Intestinal Barrier. Int. J. Mol. Sci. 2021, 22, 7613. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M. Leaky Gut: Mechanisms, Measurement and Clinical Implications in Humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Dargenio, V.N.; Dargenio, C.; Castellaneta, S.; De Giacomo, A.; Laguardia, M.; Schettini, F.; Francavilla, R.; Cristofori, F. Intestinal Barrier Dysfunction and Microbiota–Gut–Brain Axis: Possible Implications in the Pathogenesis and Treatment of Autism Spectrum Disorder. Nutrients 2023, 15, 1620. [Google Scholar] [CrossRef] [PubMed]
- Kinashi, Y.; Hase, K. Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front. Immunol. 2021, 12, 673708. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Whitley, C.S.; Haribabu, B.; Jala, V.R. Regulation of Intestinal Barrier Function by Microbial Metabolites. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1463–1482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, R.; Zhang, D.; Qi, S.; Liu, Y. Metabolite Interactions between Host and Microbiota during Health and Disease: Which Feeds the Other? Biomed. Pharmacother. 2023, 160, 114295. [Google Scholar] [CrossRef]
- Taleb, S. Tryptophan Dietary Impacts Gut Barrier and Metabolic Diseases. Front. Immunol. 2019, 10, 2113. [Google Scholar] [CrossRef]
- Ye, X.; Li, H.; Anjum, K.; Zhong, X.; Miao, S.; Zheng, G.; Liu, W.; Li, L. Dual Role of Indoles Derived From Intestinal Microbiota on Human Health. Front. Immunol. 2022, 13, 903526. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Rappa, F.; Fucarino, A.; Pitruzzella, A.; David, S.; Campanella, C. Exosomes: Can Doctors Still Ignore Their Existence? EuroMediterranean Biomed. J. 2013, 8, 136–139. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Maqueda, D.; Miralles, B.; Recio, I. HT29 Cell Line. In The Impact of Food Bioactives on Health; Springer International Publishing: Cham, Switzerland, 2015; pp. 113–124. [Google Scholar]
- Bock, H.-J.; Lee, N.-K.; Paik, H.-D. Neuroprotective Effects of Heat-Killed Levilactobacillus Brevis KU15152 on H2O2-Induced Oxidative Stress. J. Microbiol. Biotechnol. 2023, 33, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Guan, C.; Wang, T.; Mu, G.; Tuo, Y. Indole-3-Lactic Acid, a Tryptophan Metabolite of Lactiplantibacillus Plantarum DPUL-S164, Improved Intestinal Barrier Damage by Activating AhR and Nrf2 Signaling Pathways. J. Agric. Food Chem. 2023, 71, 18792–18801. [Google Scholar] [CrossRef]
- Rao, R.K.; Baker, R.D.; Baker, S.S.; Gupta, A.; Holycross, M. Oxidant-Induced Disruption of Intestinal Epithelial Barrier Function: Role of Protein Tyrosine Phosphorylation. Am. J. Physiol. Gastrointest. Liver Physiol. 1997, 273, G812–G823. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, C.; Li, Y.; Li, Z.; Li, C.; Luan, C. Antioxidant Effects and Probiotic Properties of Latilactobacillus Sakei MS103 Isolated from Sweet Pickled Garlic. Foods 2023, 12, 4276. [Google Scholar] [CrossRef] [PubMed]
- Graziano, F.; Iacopino, D.G.; Cammarata, G.; Scalia, G.; Campanella, C.; Giannone, A.G.; Porcasi, R.; Florena, A.M.; Conway de Macario, E.; Macario, A.J.L.; et al. The Triad Hsp60-MiRNAs-Extracellular Vesicles in Brain Tumors: Assessing Its Components for Understanding Tumorigenesis and Monitoring Patients. Appl. Sci. 2021, 11, 2867. [Google Scholar] [CrossRef]
- Paterna, A.; Rao, E.; Adamo, G.; Raccosta, S.; Picciotto, S.; Romancino, D.; Noto, R.; Touzet, N.; Bongiovanni, A.; Manno, M. Isolation of Extracellular Vesicles From Microalgae: A Renewable and Scalable Bioprocess. Front. Bioeng. Biotechnol. 2022, 10, 836747. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Lo Cascio, F.; Mocciaro, E.; Vitale, A.M.; Vergilio, G.; Pace, A.; Cappello, F.; Campanella, C.; Palumbo Piccionello, A. Curcumin Affects HSP60 Folding Activity and Levels in Neuroblastoma Cells. Int. J. Mol. Sci. 2020, 21, 661. [Google Scholar] [CrossRef]
- Mu, G.; Gao, Y.; Tuo, Y.; Li, H.; Zhang, Y.; Qian, F.; Jiang, S. Assessing and Comparing Antioxidant Activities of Lactobacilli Strains by Using Different Chemical and Cellular Antioxidant Methods. J. Dairy Sci. 2018, 101, 10792–10806. [Google Scholar] [CrossRef]
- Alves, N.J.; Turner, K.B.; Walper, S.A. Directed Protein Packaging within Outer Membrane Vesicles from Escherichia Coli: Design, Production and Purification. J. Vis. Exp. 2016, 117, e54458. [Google Scholar] [CrossRef] [PubMed]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G.; Pandol, S.J.; Grady, E.F.; Makharia, G.K. Brain-Gut-Microbe Communication in Health and Disease. Front. Physiol. 2011, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; Kennedy, P.J. Irritable Bowel Syndrome: A Microbiome-Gut-Brain Axis Disorder? World J. Gastroenterol. 2014, 20, 14105–14125. [Google Scholar] [CrossRef]
- Lamas, B.; Richard, M.L.; Leducq, V.; Pham, H.-P.; Michel, M.-L.; Da Costa, G.; Bridonneau, C.; Jegou, S.; Hoffmann, T.W.; Natividad, J.M.; et al. CARD9 Impacts Colitis by Altering Gut Microbiota Metabolism of Tryptophan into Aryl Hydrocarbon Receptor Ligands HHS Public Access Author Manuscript. Isabelle Nion-Larmurier 2017, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Pan, T.; Li, L.; Wang, H.; Zhu, J.; Zhang, H.; Zhao, J.; Chen, W.; Lu, W. Bifidobacterium Longum Mediated Tryptophan Metabolism to Improve Atopic Dermatitis via the Gut-Skin Axis. Gut Microbes 2022, 14, 2044723. [Google Scholar] [CrossRef] [PubMed]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.-Q. Kynurenines in the Mammalian Brain: When Physiology Meets Pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef]
- Black, C.J.; Drossman, D.A.; Talley, N.J.; Ruddy, J.; Ford, A.C. Functional Gastrointestinal Disorders: Advances in Understanding and Management. Lancet 2020, 396, 1664–1674. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Ghoshal, U.C. Gut Microbiota-Brain Axis Modulation by a Healthier Microbiological Microenvironment: Facts and Fictions. J. Neurogastroenterol. Motil. 2018, 24, 4–6. [Google Scholar] [CrossRef]
- McKenzie, Y.A.; Bowyer, R.K.; Leach, H.; Gulia, P.; Horobin, J.; O’Sullivan, N.A.; Pettitt, C.; Reeves, L.B.; Seamark, L.; Williams, M.; et al. British Dietetic Association Systematic Review and Evidence-based Practice Guidelines for the Dietary Management of Irritable Bowel Syndrome in Adults (2016 Update). J. Hum. Nutr. Diet. 2016, 29, 549–575. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Cremon, C.; Bellini, M.; Corsetti, M.; Di Nardo, G.; Falangone, F.; Fuccio, L.; Galeazzi, F.; Iovino, P.; Sarnelli, G.; et al. Italian Guidelines for the Management of Irritable Bowel Syndrome. Dig. Liver Dis. 2023, 55, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Heaton, K.W.; Ghosh, S.; Braddon, F.E. How Bad Are the Symptoms and Bowel Dysfunction of Patients with the Irritable Bowel Syndrome? A Prospective, Controlled Study with Emphasis on Stool Form. Gut 1991, 32, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, I.K.; Fullerton, S.; Hawkey, C.J.; Jones, R.H.; Longstreth, G.F.; Mayer, E.A.; Peacock, R.A.; Wilson, I.K.; Naesdal, J. An Irritable Bowel Syndrome-Specific Symptom Questionnaire: Development and Validation. Scand. J. Gastroenterol. 2003, 38, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Fang, L.; Dong, Y.; Yang, J.; Chen, X.; Zhang, N.; Zhu, Y.; Huang, T. Mitochondrial Quality Control Protects Photoreceptors against Oxidative Stress in the H2O2-Induced Models of Retinal Degeneration Diseases. Cell Death Dis. 2021, 12, 413. [Google Scholar] [CrossRef]
- Basuroy, S.; Sheth, P.; Kuppuswamy, D.; Balasubramanian, S.; Ray, R.M.; Rao, R.K. Expression of Kinase-Inactive c-Src Delays Oxidative Stress-Induced Disassembly and Accelerates Calcium-Mediated Reassembly of Tight Junctions in the Caco-2 Cell Monolayer. J. Biol. Chem. 2003, 278, 11916–11924. [Google Scholar] [CrossRef] [PubMed]
- Alexeev, E.E.; Lanis, J.M.; Kao, D.J.; Campbell, E.L.; Kelly, C.J.; Battista, K.D.; Gerich, M.E.; Jenkins, B.R.; Walk, S.T.; Kominsky, D.J.; et al. Microbiota-Derived Indole Metabolites Promote Human and Murine Intestinal Homeostasis through Regulation of Interleukin-10 Receptor. Am. J. Pathol. 2018, 188, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Grifka-Walk, H.M.; Jenkins, B.R.; Kominsky, D.J. Amino Acid Trp: The Far Out Impacts of Host and Commensal Tryptophan Metabolism. Front. Immunol. 2021, 12, 653208. [Google Scholar] [CrossRef] [PubMed]
- Israelyan, N.; Del Colle, A.; Li, Z.; Park, Y.; Xing, A.; Jacobsen, J.P.R.; Luna, R.A.; Jensen, D.D.; Madra, M.; Saurman, V.; et al. Effects of Serotonin and Slow-Release 5-HTP on Gastrointestinal Motility in a Mouse Model of Depression. Gastroenterology, 2019; 157, 507–521.e4. [Google Scholar] [CrossRef]
- Correia, A.S.; Silva, I.; Oliveira, J.C.; Reguengo, H.; Vale, N. Serotonin Type 3 Receptor Is Potentially Involved in Cellular Stress Induced by Hydrogen Peroxide. Life 2022, 12, 1645. [Google Scholar] [CrossRef]
- Foster, J.A. Gut Microbiome and Behavior: Focus on Neuroimmune Interactions. Int. Rev. Neurobiol. 2016, 131, 49–65. [Google Scholar]
- Pinto, R.; Belsky, J.; Baptista, J.; Carvalho, A.; Cunha, C.; Soares, I.; Mesquita, A.R. Mothers’ Distress Exposure and Children’s Withdrawn Behavior—A Moderating Role for the Interferon Gamma Gene (IFNG). Dev. Psychobiol. 2020, 62, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Merchak, A.R.; Wachamo, S.; Brown, L.C.; Thakur, A.; Moreau, B.; Brown, R.M.; Rivet-Noor, C.R.; Raghavan, T.; Gaultier, A. Lactobacillus from the Altered Schaedler Flora Maintain IFNγ Homeostasis to Promote Behavioral Stress Resilience. Brain Behav. Immun. 2024, 115, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Heng, B.; Guillemin, G.J. The Gut Microbiota, Kynurenine Pathway, and Immune System Interaction in the Development of Brain Cancer. Front. Cell Dev. Biol. 2020, 8, 562812. [Google Scholar] [CrossRef] [PubMed]
- Tomberlin, J.K.; Crippen, T.L.; Wu, G.; Griffin, A.S.; Wood, T.K.; Kilner, R.M. Indole: An Evolutionarily Conserved Influencer of Behavior across Kingdoms. BioEssays 2017, 39, 1600203. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Coloma, C.; Santaballa, A.; Sanmartín, E.; Calvo, D.; García, A.; Hervás, D.; Cordón, L.; Quintas, G.; Ripoll, F.; Panadero, J.; et al. Immunosuppressive Profiles in Liquid Biopsy at Diagnosis Predict Response to Neoadjuvant Chemotherapy in Triple-Negative Breast Cancer. Eur. J. Cancer 2020, 139, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Bianco, N.R.; Kim, S.H.; Ruffner, M.A.; Robbins, P.D. Therapeutic Effect of Exosomes from Indoleamine 2,3-dioxygenase–Positive Dendritic Cells in Collagen-induced Arthritis and Delayed-type Hypersensitivity Disease Models. Arthritis Rheum. 2009, 60, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Peck, K.A.; Ciullo, A.; Li, L.; Li, C.; Morris, A.; Marbán, E.; Ibrahim, A.G. Extracellular Vesicles Secreted by TDO2-Augmented Fibroblasts Regulate Pro-Inflammatory Response in Macrophages. Front. Cell Dev. Biol. 2021, 9, 733354. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Wang, Y.; Lu, L.; Wei, R.; Xu, B.; Hu, X.; Jiang, X.; Du, L. Effects of Combined Live Bifidobacterium, Lactobacillus, Enterococcus and Bacillus Cereus Tablets on Post-Stroke Depression and Serum Inflammatory Factorse. Discov. Med. 2023, 35, 312. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, Stability and Resilience of the Human Gut Microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Park, S.C.; Lim, J.-Y.; Jeen, Y.T.; Keum, B.; Seo, Y.S.; Kim, Y.S.; Lee, S.J.; Lee, H.S.; Chun, H.J.; Um, S.H.; et al. Ethanol-Induced DNA Damage and Repair-Related Molecules in Human Intestinal Epithelial Caco-2 Cells. Mol. Med. Rep. 2012, 5, 1027–1032. [Google Scholar] [CrossRef]
- Rao, R.K.; Basuroy, S.; Rao, V.U.; Karnaky, K.J.; Gupta, A. Tyrosine Phosphorylation and Dissociation of Occludin–ZO-1 and E-Cadherin–β-Catenin Complexes from the Cytoskeleton by Oxidative Stress. Biochem. J. 2002, 368, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, G.; Rodolico, V.; Zerilli, M.; Martorana, A.; Bucchieri, F.; Pitruzzella, A.; Gammazza, A.M.; David, S.; Rappa, F.; Zummo, G.; et al. Changes in Immunohistochemical Levels and Subcellular Localization After Therapy and Correlation and Colocalization With CD68 Suggest a Pathogenetic Role of Hsp60 in Ulcerative Colitis. Appl. Immunohistochem. Mol. Morphol. 2011, 19, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Paladino, L.; Rappa, F.; Barone, R.; Macaluso, F.; Zummo, F.P.; David, S.; Szychlinska, M.A.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; et al. NF-KB Regulation and the Chaperone System Mediate Restorative Effects of the Probiotic Lactobacillus Fermentum LF31 in the Small Intestine and Cerebellum of Mice with Ethanol-Induced Damage. Biology 2023, 12, 1394. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of Human Epithelial Tight Junction Proteins by Lactobacillus Plantarum in Vivo and Protective Effects on the Epithelial Barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed]
- Shinde, R.; McGaha, T.L. The Aryl Hydrocarbon Receptor: Connecting Immunity to the Microenvironment. Trends Immunol. 2018, 39, 1005–1020. [Google Scholar] [CrossRef]
- Bi, Z.; Zhang, S.; Meng, Y.; Feng, Y.; Wang, Y.; Wang, E.; Pan, X.; Zhu, R.; Fan, H.; Pang, S.; et al. Female Serotonin Transporter-knockout Rat: A Potential Model of Irritable Bowel Syndrome. FASEB J. 2021, 35, e21701. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Overview | Western Blotting (WB) | Immunofluorescence (IF) |
|---|---|---|---|
| Tryptophan 2,3-dioxygenase 2 (TDO 2) | SAB2105644 Rabbit Polyclonal Sigma-Aldrich, Darmstadt, Germany | 1:500 | - |
| Heat Shock Protein 60 (Hsp60) | Sc-59567 Mouse Monoclonal Santa Cruz Biotechnology, Dallas, TX, USA | 1:1000 | 1:50 |
| Heat Shock Protein 90 α/β (Hsp90) | Sc-13119 Mouse Monoclonal Santa Cruz Biotechnology, Dallas, TX, USA | 1:1000 | 1:50 |
| CD81 | Sc-70803 Mouse Monoclonal Santa Cruz Biotechnology, Dallas, TX, USA | 1:1000 | - |
| Tight Junction Protein 1 (TJP1) | C82740 Rabbit Polyclonal Sigma-Aldrich, Darmstadt, Germany | - | 1:50 |
| 5-HT2C | Ab-133570 Rabbit Monoclonal Abcam, Cambridge, UK | 1:1000 | - |
| Anti-GAPDH | ABS16 Rabbit Polyclonal Sigma-Aldrich, Darmstadt, Germany | 1:1000 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santonocito, R.; Paladino, L.; Vitale, A.M.; D'Amico, G.; Zummo, F.P.; Pirrotta, P.; Raccosta, S.; Manno, M.; Accomando, S.; D’Arpa, F.; et al. Nanovesicular Mediation of the Gut–Brain Axis by Probiotics: Insights into Irritable Bowel Syndrome. Biology 2024, 13, 296. https://doi.org/10.3390/biology13050296
Santonocito R, Paladino L, Vitale AM, D'Amico G, Zummo FP, Pirrotta P, Raccosta S, Manno M, Accomando S, D’Arpa F, et al. Nanovesicular Mediation of the Gut–Brain Axis by Probiotics: Insights into Irritable Bowel Syndrome. Biology. 2024; 13(5):296. https://doi.org/10.3390/biology13050296
Chicago/Turabian StyleSantonocito, Radha, Letizia Paladino, Alessandra Maria Vitale, Giuseppa D'Amico, Francesco Paolo Zummo, Paolo Pirrotta, Samuele Raccosta, Mauro Manno, Salvatore Accomando, Francesco D’Arpa, and et al. 2024. "Nanovesicular Mediation of the Gut–Brain Axis by Probiotics: Insights into Irritable Bowel Syndrome" Biology 13, no. 5: 296. https://doi.org/10.3390/biology13050296
APA StyleSantonocito, R., Paladino, L., Vitale, A. M., D'Amico, G., Zummo, F. P., Pirrotta, P., Raccosta, S., Manno, M., Accomando, S., D’Arpa, F., Carini, F., Barone, R., Rappa, F., Marino Gammazza, A., Bucchieri, F., Cappello, F., & Caruso Bavisotto, C. (2024). Nanovesicular Mediation of the Gut–Brain Axis by Probiotics: Insights into Irritable Bowel Syndrome. Biology, 13(5), 296. https://doi.org/10.3390/biology13050296