
New Methods in Digital Wood Anatomy: The Use of Pixel-Contrast Densitometry with Example of Angiosperm Shrubs in Southern Siberia

 , , and
, , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection
2.3. Cell Structure Analysis
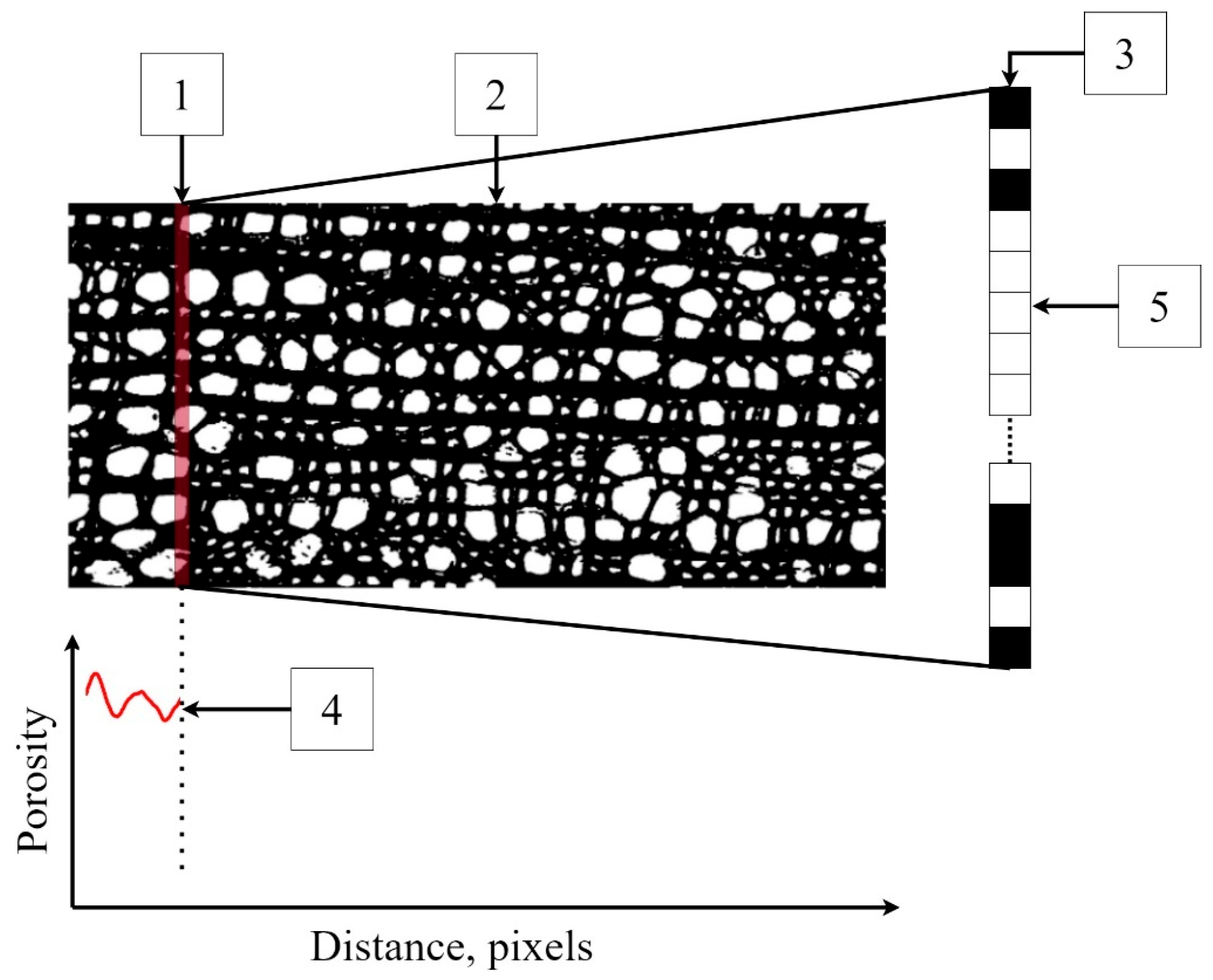
2.4. Implementation of PiC Densitometry and Software
3. Results and Discussion
3.1. Influence of the Selection of the Scanning Area on the Measured Porosity Profile
3.2. Comparison of Typical Porosity Profiles of the Studied Species
3.3. Comparative Analysis of Porosity Profile Indicators and Ring width Using Three Long-Living Species
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Arx, G.; Carrer, M. ROXAS. A new tool to build centuries-long tracheid-lumen chronologies in conifers. Dendrochronologia 2014, 32, 290–293. [Google Scholar] [CrossRef]
- Ziaco, E.; Biondi, F.; Heinrich, I. Wood cellular dendroclimatology: Testing new proxies in Great Basin bristlecone pine. Front. Plant Sci. 2016, 7, 1602. [Google Scholar] [CrossRef] [PubMed]
- Ziaco, E.; Liang, E. New perspectives on sub-seasonal xylem anatomical responses to climatic variability. Trees 2019, 33, 973–975. [Google Scholar] [CrossRef]
- Anchukaitis, K.J.; Evans, M.N.; Hughes, M.K.; Vaganov, E.A. An interpreted language implementation of the Vaganov–Shashkin tree-ring proxy system model. Dendrochronologia 2020, 60, 125677. [Google Scholar] [CrossRef]
- Tabakova, K.A.; Arzak, A.; Carrer, M.; Vaganov, E.A.; Kirdyanov, A.V. Contribution of Russian dendroanatomical studies to the dendrochronology since the mid-20th century. Dendrochronologia 2023, 81, 126128. [Google Scholar] [CrossRef]
- Fonti, P.; Eilmann, B.; Garcia-Gonsalez, I.; von Arx, G. Expeditious building of ring-porous earlywood vessel chronologies without loosing signal information. Trees 2009, 23, 665–671. [Google Scholar] [CrossRef]
- Noyer, E.; Stojanovic, M.; Horacek, P.; Perez-de-Lis, G. Toward a better understanding of angiosperm xylogenesis: A new method for a cellular approach. N. Phytol. 2023, 239, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Silkin, P.P.; Kirdyanov, A.V.; Krusic, P.J.; Ekimov, M.V.; Barinov, V.V.; Büntgen, U. A new approach to measuring tree-ring density parameters. J. Sib. Fed. Univ. Biol. 2022, 15, 441–455. [Google Scholar]
- Menezes-Silva, P.E.; Loram-Lourenço, L.; Alves, R.D.F.B.; Sousa, L.F.; Almeida, S.E.S.; Farnese, F.S. Different ways to die in a changing world: Consequences of climate change for tree species performance and survival through an ecophysiological perspective. Ecol. Evol. 2019, 9, 11979–11999. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S. Climatic influence on tree wood anatomy: A review. J. Wood Sci. 2021, 67, 24. [Google Scholar] [CrossRef]
- Alisov, B.P. Climate of the USSR; Moscow State University: Moscow, Russia, 1956. (In Russian) [Google Scholar]
- Polikarpov, N.P.; Nazimova, D.I. The dark coniferous forests of the northern part of the west Siberian mountains. For. Res. For. Sib. 1963, 57, 103–147. [Google Scholar]
- Furst, G.G. Methods of Anatomical-Histochemical Research of Plant Tissues; Nauka: Moscow, Russia, 1979. (In Russian) [Google Scholar]
- Kellogg, R.M.; Wangaard, F.F. Variation in the cell-wall density of wood. Wood Fiber Sci. 1969, 1, 180–204. [Google Scholar]
- Kellogg, R.M.; Sastry, C.B.R.; Wellwood, R.W. Relationships between cell-wall composition and cell-wall density. Wood Fiber Sci. 1975, 7, 170–177. [Google Scholar]
- Janusz, S.; Danilov, D. Density of wood of pine and spruce in the postagrogenic soil of the boreal zone. In Proceedings of the Annual 24th International Scientific Conference Proceedings, Research for Rural Development 2018, Jelgava, Latvia, 16–18 May 2018; Latvia University of Life Sciences and Technologies: Jelgava, Latvia, 2018; Volume 1, pp. 92–96. [Google Scholar] [CrossRef]
- Park, W.K.; Telewski, F.W. Measuring maximum latewood density by image analysis at the cellular level. Wood Fiber Sci. 1993, 25, 326–332. [Google Scholar]
- Decoux, V.; Varcin, É.; Leban, J.M. Relationships between the intra-ring wood density assessed by X-ray densitometry and optical anatomical measurements in conifers. Consequences for the cell wall apparent density determination. Ann. For. Sci. 2004, 61, 251–262. [Google Scholar] [CrossRef]
- Wassenberg, M.; Montwé, D.; Kahle, H.P.; Spiecker, H. Exploring high frequency densitometry calibration functions for different tree species. Dendrochronologia 2014, 32, 273–281. [Google Scholar] [CrossRef]
- Terskov, I.A.; Vaganov, E.A.; Spirov, V.V. New methods of studying the within-ring distribution of porosity and wood density. News Sib. Branch USSR Acad. Sci. 1972, 5, 115–120. [Google Scholar]
- Vaganov, E.A.; Smirnov, V.V.; Terskov, I.A. On the possibility of determining the rate of seasonal growth of a stem in thickness and changes in the water regime of a tree using a photometric curve. Ecology 1975, 2, 45–53. (In Russian) [Google Scholar]
- Gedraite, E.S.; Hadad, M. Investigation on the effect of a Gaussian Blur in image filtering and segmentation. In Proceedings of the ELMAR-2011, Zadar, Croatia, 14–16 September 2011; IEEE: Piscataway, NJ, USA, 2011; pp. 393–396. [Google Scholar]
- Vahadane, A.; Sethi, A. Towards generalized nuclear segmentation in histological images. In Proceedings of the 13th IEEE International Conference on BioInformatics and BioEngineering, Chania, Greece, 10–13 November 2013; IEEE: Piscataway, NJ, USA, 2013; pp. 1–4. [Google Scholar]
- McCann, M.T.; Majumdar, J.; Peng, C.; Castro, C.A.; Kovačević, J. Algorithm and benchmark dataset for stain separation in histology images. In Proceedings of the 2014 IEEE International Conference on Image Processing (ICIP), Paris, France, 27–30 October 2014; IEEE: Piscataway, NJ, USA, 2014; pp. 3953–3957. [Google Scholar]
- Xu, X.; Xu, S.; Jin, L.; Song, E. Characteristic analysis of Otsu threshold and its applications. Pattern Recognit. Lett. 2011, 32, 956–961. [Google Scholar] [CrossRef]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization (Dendrochronology, Forestry, Dendroclimatology, Autoregressive Process). Ph.D. Thesis, The University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Wheeler, E.A.; Baas, P.; Gasson, P.E. IAWA list of microscopic features for hardwood identification. IAWA Bull. 1989, 10, 219–332. [Google Scholar] [CrossRef]
- Benkova, V.E.; Schweingruber, F.H. Anatomy of Russian Woods: An Atlas for the Identification of Trees, Shrubs, Dwarf Shrubs and Woody Lianas from Russia; Swiss Federal Institute for Forest, Snow and Landscape Research: Birmensdorf, Switzerland; Haupt Verlag: Bern, Switzerland, 2004; (In English and Russian). [Google Scholar]
- Garcia-Gonzalez, I.; Souto-Herrero, M.; Campelo, F. Ring-porosity and earlywood vessels: A review on extracting environmental information through time. IAWA J. 2016, 37, 295–314. [Google Scholar] [CrossRef]
- Venkat, A.; Muneer, S. Role of circadian rhythms in major plant metabolic and signaling pathways. Front. Plant Sci. 2022, 13, 836244. [Google Scholar] [CrossRef] [PubMed]
- Mork, E. Die Qualitat des Fichtenhozes unter besonder Rucksichtnahme auf Schleif und Papierholz. Pap. Fabr. 1928, 26, 741–747. (In German) [Google Scholar]
- Denne, M.P. Definition of latewood according to Mork (1928). IAWA Bull. 1989, 10, 59–62. [Google Scholar] [CrossRef]
- Creber, G.T.; Chaloner, W.G. Influence of environmental factors on the wood structure of living and fossil trees. Bot. Rev. 1984, 50, 357–448. [Google Scholar] [CrossRef]
- Koubaa, A.; Zhang, S.T.; Makni, S. Defining the transition from earlywood to latewood in black spruce based on intra-ring wood density profiles from X-ray densitometry. Ann. For. Sci. 2002, 59, 511–518. [Google Scholar] [CrossRef]
- Kumar, S. Earlywood–latewood demarcation criteria and their effect on genetic parameters of growth ring density components and efficiency of selection for end-of-rotation density of radiata pine. Silv. Genet. 2002, 51, 241–246. [Google Scholar]
- Tardif, J.C.; Kames, S.; Nolin, A.F.; Bergeron, Y. Earlywood vessels in black ash (Fraxinus nigra Marsh.) trees show contrasting sensitivity to hydroclimate variables according to flood exposure. Front. Plant Sci. 2021, 12, 754596. [Google Scholar] [CrossRef]
- Belokopytova, L.V.; Zhirnova, D.F.; Yang, B.; Babushkina, E.A.; Vaganov, E.A. Modeling of the statistical distribution of tracheids in conifer rings: Finding universal criterion for earlywood–latewood distinction. Plants 2023, 12, 3454. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.E.; Anfodillo, T.; Gleason, S.M.; McCulloh, K.A. Tip-to-base xylem conduit widening as an adaptation: Causes, consequences, and empirical priorities. N. Phytol. 2021, 229, 1877–1893. [Google Scholar] [CrossRef] [PubMed]
- Skomarkova, M.V.; Vaganov, E.A.; Mund, M.; Knohl, A.; Linke, P.; Boerner, A.; Schulze, E.D. Inter-annual and seasonal variability of radial growth, wood density and carbon isotope ratios in tree rings of beech (Fagus sylvatica) growing in Germany and Italy. Trees 2006, 20, 571–586. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Fonti, P.; Cherubini, P.; Martín-Benito, D.; Chaar, H.; Cañellas, I. Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability. Tree Physiol. 2012, 32, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Carrer, M.; Brunetti, M.; Castagneri, D. The imprint of extreme climate events in century-long time series of wood anatomical traits in high-elevation conifers. Front. Plant Sci. 2016, 7, 187577. [Google Scholar] [CrossRef] [PubMed]
- Castagneri, D.; Fonti, P.; von Arx, G.; Carrer, M. How does climate influence xylem morphogenesis over the growing season? Insights from long-term intra-ring anatomy in Picea abies. Ann. Bot. 2017, 119, 1011–1020. [Google Scholar] [CrossRef]
- Rita, A.; Camarero, J.J.; Colangelo, M.; de Andrés, E.G.; Pompa-García, M. Wood anatomical traits respond to climate but more individualistically as compared to radial growth: Analyze trees, not means. Forests 2022, 13, 956. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Scanning Area | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Cornus alba | 0.87 | 0.94 | 0.94 | 0.91 | 0.82 |
| Alnus alnobetula | 0.82 | 0.89 | 0.93 | 0.93 | 0.85 |
| Prunus padus | 0.86 | 0.94 | 0.97 | 0.95 | 0.98 |
| Caragana arborescens | 0.79 | 0.94 | 0.97 | 0.92 | 0.82 |
| Caragana altaica | 0.96 | 0.98 | 0.98 | 0.88 | 0.95 |
| Rhododendron ledebourii | 0.95 | 0.96 | 0.96 | 0.79 | 0.89 |
| Ribes nigrum | 0.91 | 0.95 | 0.92 | 0.91 | 0.79 |
| Spiraea hypericifolia | 0.60 | 0.87 | 0.99 | 0.97 | 0.95 |
| Characteristics | Species | ||
|---|---|---|---|
| Caragana arborescens | Prunus padus | Alnus alnobetula | |
| RW | 0.41 | 0.17 | 0.28 |
| MEAN Por | 0.09 | 0.53 | 0.17 |
| MAX Por | 0.19 | 0.30 | 0.14 |
| MIN Por | 0.03 | 0.47 | 0.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khudykh, T.A.; Belokopytova, L.V.; Yang, B.; Kholdaenko, Y.A.; Babushkina, E.A.; Vaganov, E.A. New Methods in Digital Wood Anatomy: The Use of Pixel-Contrast Densitometry with Example of Angiosperm Shrubs in Southern Siberia. Biology 2024, 13, 223. https://doi.org/10.3390/biology13040223
Khudykh TA, Belokopytova LV, Yang B, Kholdaenko YA, Babushkina EA, Vaganov EA. New Methods in Digital Wood Anatomy: The Use of Pixel-Contrast Densitometry with Example of Angiosperm Shrubs in Southern Siberia. Biology. 2024; 13(4):223. https://doi.org/10.3390/biology13040223
Chicago/Turabian StyleKhudykh, Timofey A., Liliana V. Belokopytova, Bao Yang, Yulia A. Kholdaenko, Elena A. Babushkina, and Eugene A. Vaganov. 2024. "New Methods in Digital Wood Anatomy: The Use of Pixel-Contrast Densitometry with Example of Angiosperm Shrubs in Southern Siberia" Biology 13, no. 4: 223. https://doi.org/10.3390/biology13040223
APA StyleKhudykh, T. A., Belokopytova, L. V., Yang, B., Kholdaenko, Y. A., Babushkina, E. A., & Vaganov, E. A. (2024). New Methods in Digital Wood Anatomy: The Use of Pixel-Contrast Densitometry with Example of Angiosperm Shrubs in Southern Siberia. Biology, 13(4), 223. https://doi.org/10.3390/biology13040223