1. Introduction
Onions (
Allium cepa L.), ubiquitous in kitchens worldwide, stand as a culinary cornerstone, adding depth and flavor to many dishes. Studies have highlighted their richness in phytochemicals, particularly medicinal flavonoids [
1]. Onions hold substantial economic importance globally and are paramount in Egypt for both local consumption and export. However, recent years have witnessed a significant decline in onion production, primarily attributed to the devastating white rot disease caused by
S. cepivorum [
2]. This continual threat poses a persistent risk to
Allium spp. cultivation, triggering disease outbreaks wherever such crops are grown.
The disease displays noticeable symptoms above ground, including wilting, yellowing of older leaves, and die-back of leaf tips. Eventually, collapse and decay occur as the symptoms advance along the leaf blades. Concurrently, below the ground, root infection leads to watery decay and the appearance of fluffy, white mycelial growth at the bulb base. Numerous sclerotia form within these mycelial mats, serving as the main source of inoculum for the next cultivated crops [
3]. This pathogen severely threatens onion cultivation, leading to significant yield losses and economic setbacks. It has become widely prevalent in Egypt, leading to severe damage and, at times, complete crop loss of up to 100%, as noted by Ahmed and Ahmed [
4]. Notably, this pathogenic menace is a major constraint on onion cultivation, particularly in Upper Egypt [
5]. Researchers exploring strategies to manage white rot disease consider both efficacy and environmental implications, with traditional fungicides serving as a reliable but not always eco-friendly solution [
6]. Among the alternatives are nanoparticles and chemical inducers, such as salicylic acid, which have demonstrated promising results in managing plant diseases [
7,
8].
Nanoparticles, with their minute size and unique properties, offer a novel avenue for disease management. Recent studies have highlighted the potential of nanoparticles in inhibiting the growth and spread of pathogens, including
S. cepivorum [
3]. When strategically employed, these microscopic wonders can target the pathogen while minimizing the impact on non-target organisms and the environment. In addition, nanoparticles can stimulate morphological and physiological changes in plants, with their efficacy influenced by factors like the composition, size, surface properties, reactivity, and dosage [
9]. The concentration of NPs is crucial for effectiveness, and nanotechnology presents unique applications in biotechnology and agriculture [
10]. NPs can serve as targeted carriers, delivering substances like herbicides, pesticides, fertilizers, or genes to specific cellular organelles in plants [
11]. Despite the potential benefits, inadequate information exists on NP toxicity in plants, and few studies have explored the mechanisms underlying NPs’ influences on plant growth. Understanding NP action’s physiological, biochemical, and molecular mechanisms is essential for optimizing their impact on host growth [
12]. The World Health Organization (WHO) and the United States Environmental Protection Agency (USEPA) set limits for heavy metals in drinking water: 5 mg/L for Zn, 0.3 mg/L for Fe, and 2 mg/L for Cu by the WHO; 1.3 mg/L for Cu by the USEPA [
13].
Inducers, particularly salicylic acid, have become key players in plant defense mechanisms. Salicylic acid is a signaling molecule that activates the plant’s immune responses, fortifying its defenses against invading pathogens [
14]. Incorporating these chemical inducers into disease management strategies holds promise, providing a targeted and potentially sustainable approach to combating white rot in onions. The shift toward nanoparticle and chemical inducer-based approaches represents a conscious effort to balance disease control and ecological responsibility. By exploring these alternatives, we aim to safeguard onion crops and cultivate a more sustainable and environmentally conscious agricultural landscape.
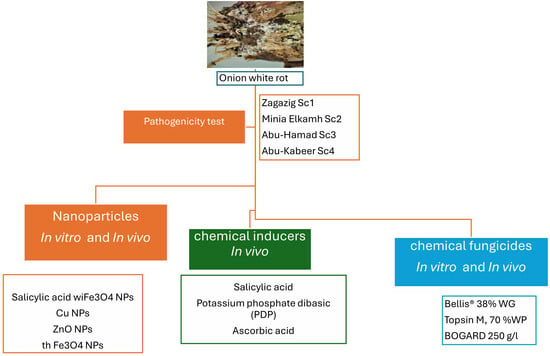
This study aims to compare various strategies for managing onion white rot disease caused by S. cepivorum. These strategies include the application of nanoparticles (Fe3O4 NPs, Cu NPs, and ZnO NPs), chemical inducers (salicylic acid, PDP, ascorbic acid), and traditional fungicides. Specifically, our objectives encompass evaluating the effectiveness of these alternative methods in inhibiting the mycelial growth of S. cepivorum in vitro, assessing their ability to reduce the disease incidence and severity in infected onions under greenhouse conditions, and investigating their impact on key plant health parameters, including the chlorophyll content (a, b, and carotenoids) and phenolic compounds (total, free and bound), in infected onions under various management strategies. Furthermore, we aim to compare the efficacy of nanoparticle-based treatments, chemical inducers, and traditional fungicides to identify the most promising eco-friendly management strategies for onion white rot. Through this comprehensive analysis, we seek to provide insights into sustainable agriculture by offering recommendations for adopting effective and environmentally friendly management practices for onion white rot disease.
4. Discussion
Nanomaterials have potent antimicrobial effects due to the increased surface contact with cells, triggering reactions that generate reactive oxygen species (ROS), damaging proteins, and disrupting cell membranes. These interactions make nanomaterial-based antimicrobials resistant to pathogen resistance [
32,
33,
34,
35].
In addition, nanoparticles can stimulate morphological and physiological changes in plants, with their efficacy influenced by factors like the composition, size, surface properties, reactivity, and dosage [
9]. The concentration of NPs is crucial for their effectiveness, and nanotechnology presents unique applications in biotechnology and agriculture [
10]. NPs can serve as targeted carriers, delivering substances like herbicides, pesticides, fertilizers, or genes to specific cellular organelles in plants [
11]. Despite the potential benefits, inadequate information exists on NP toxicity in plants, and few studies have explored the mechanisms underlying NP influences on plant growth. Understanding NP action’s physiological, biochemical, and molecular mechanisms is essential for optimizing the impact on host growth [
12].
It is evident that the pathogenicity of
S. cepivorum isolates varies significantly, impacting the Giza 20 onion cultivar under controlled greenhouse conditions. The maximum disease incidence and severity were observed in the third isolate from Abu-Hamad, suggesting a potent pathogenic strain in that region. This aligns with previous research by Tyson et al. [
36], who found regional variations in the virulence of
S. cepivorum isolates. The diverse responses of isolates one and two, from Zagazig and Minia Elkamh, may be due to environmental factors or genetic differences in the isolates themselves [
37].
The reduced plant weights, both fresh and dry, further underscore the detrimental effects of the pathogen on the Giza 20 onion cultivar. Similar findings were reported by Elshahawy et al. [
38], who demonstrated a correlation between the disease severity and diminished plant growth in onion crops infected with
S. cepivorum. Isolate four from Abu-Kabeer exhibited intermediate effects, indicating a moderate pathogenicity compared to the other isolates. This aligns with the concept of strain-specific variations in virulence, as discussed by Garcia-Rubio et al. [
39]. Several factors may contribute to the observed trends, such as the genetic diversity of
S. cepivorum isolates [
40,
41] and environmental conditions influencing the pathogen–host interaction [
42]. Understanding these factors is crucial for developing effective disease management strategies.
The effect of diverse NPs on the linear growth and reduction percentage of
S. cepivorum provides insights into potential antifungal strategies [
43]. The findings align with previous studies on the antifungal properties of nanoparticles. For instance, research by Wu et al. [
44] on Fe
3O
4 NPs corroborates the dose-dependent inhibition observed in this study. The substantial reduction in linear growth with increasing concentrations of Fe
3O
4 NPs suggests a potential mechanism of action, possibly involving interference with fungal cell membranes or vital cellular processes, as discussed by Rawat et al. [
45]. The Cu NPs and ZnO NPs also exhibited dose-dependent inhibitory effects, consistent with studies on their antifungal activities against various pathogens [
46]. The high reduction percentages at 100 μg/L indicate their potential as effective agents against
S. cepivorum, which aligns with Derbalah et al. [
47]. These nanoparticles’ mode of action could involve ROS generation, disrupting cellular functions [
48].
Combining salicylic acid with Fe
3O
4 NPs demonstrated remarkable efficacy, achieving a substantial reduction percentage [
49]. This aligns with studies highlighting the synergistic effects of combining nanoparticles with other antifungal agents [
50]. Salicylic acid, known for its role in plant defense mechanisms, may enhance the antifungal activity of nanoparticles through various biochemical pathways [
49]. Factors such as the nanoparticle size, surface charge, and interactions with fungal cell components may influence their efficacy [
51]. Iron acts as a cofactor for various enzyme systems and plays a crucial role in plant cells, constituting a significant portion of metal microelements and supporting the activity of enzymes like peroxidases and catalases [
52].
The results showed that the Fe
3O
4 NPs at varying concentrations resulted in disease reduction and improved plant growth, while the Cu NPs also exhibited disease reduction and increased plant growth. Additionally, the ZnO NPs showed similar effects. The Fe
3O
4 NPs further reduced disease and improved plant growth when combined with salicylic acid. These results suggest the ability of these nanoparticles to manage
S. cepivorum infection in onions and growth enhancement, which is in line with Faizan et al. [
53]. A study by Khan et al. [
54] found that Fe
3O
4 NPs efficiently lowered the disease incidence and severity in tomato plants infected with a fungal pathogen. Similarly, the work of Gaba et al. [
55] demonstrated the disease-suppressive effects of Cu nanoparticles on Alternaria blight disease and other various plant pathogens, as reported by Elmer and White [
56]. Furthermore, the study by Zhang et al. [
57] highlighted the positive influence of ZnO NPs on rice growth and disease resistance.
The trends observed in our study could be because of the unique physicochemical properties of the nanoparticles. For example, Fe
3O
4 NPs were confirmed to induce systemic resistance in plants by modulating defense-related gene expression and activating signaling pathways [
58]. Similarly, Cu NPs have been shown to possess antifungal properties, disrupting the cell membranes of pathogens and inhibiting their growth [
59]. Moreover, ZnO NPs enhanced host growth by promoting nutrient uptake and modifying physiological processes [
60,
61]. Moreover, the combination of salicylic acid and Fe
3O
4 NPs resulted in further disease reduction and improved plant growth [
62,
63], demonstrating nanoparticles’ enhanced antifungal activity when combined with plant defense signaling compounds. ZnO NPs had the most significant positive impact, increasing the chlorophyll levels at higher concentrations [
64,
65].
This study demonstrated that different nanoparticles exhibit varying effects on the disease incidence and severity in
S. cepivorum-infected onions. The Cu NPs and the combination of salicylic acid and Fe
3O
4 NPs showed promising results in reducing the disease incidence and severity. This aligns with the findings of a study by Pariona et al. [
66], which reported the antifungal activity of copper nanoparticles against plant pathogens. The ability of the Cu NPs to suppress the growth and development of fungal pathogens could explain their effectiveness in reducing the disease incidence and severity in infected onions. Therefore, Cu NPs are an affordable solution for combating fungal diseases, showing strong antifungal activity. Studies reveal that Cu NPs can penetrate cell membranes, causing rupture, impeding colony growth, and inhibiting mycelium and spore growth [
67]. The spherical shape enhances their antifungal activity [
68]. Interestingly, Cu NPs completely inhibited phytopathogens at a concentration of 1000 ppm, indicating their promise in managing agricultural diseases [
69]. CuO-NPs possess versatile properties, making them valuable in drug delivery and anticancer, antifungal, antioxidant, and antibacterial applications [
70,
71].
Cu NPs can modify physiological and biochemical processes in plants, impacting germination and growth. Elevated concentrations in the solution culture have been detrimental to wheat plants, whereas lower concentrations in the growth medium improved the yield parameters [
72]. Despite copper being an essential micronutrient, excessive exposure can lead to phytotoxicity, inducing oxidative stress and DNA damage in plants [
73]. CuO-NPs and ZnO-NPs showed stronger antifungal effects, forming larger inhibition zones and causing the distortion of fungal hyphae, inhibiting conidia and conidiophores’ expansion, leading to cell death [
71,
74].
The application of zinc oxide (ZnO) and copper (Cu) nanoparticles has demonstrated improvements in seed quality [
75]. This treatment led to increased growth parameters, relative water content, and biochemical factors like total phenolic and proline contents, catalase (CAT), peroxidase (POD), and glutathione S-transferase [
76]. Similarly, treating chickpea plants with ZnO NPs below 30 nm at a concentration of 1.5 ppm significantly increased the total biomass compared to 10 ppm. Higher concentrations (10 ppm) were correlated with decreased relative water levels in plant leaves [
77]. Nanoparticles were used as nano-priming agents for antibacterial and antifungal activities against phytopathogens, besides enhancing plant growth [
78]. Using Fe
2O
3 NPs as co-fertilizers at lower concentrations can effectively improve chickpea growth [
79]. Studies by George et al. [
80] on the anti-fungal activity of ZnO and TiO
2 nanoparticles against various fungal pathogens demonstrated their efficacy compared to bulk particles. Both nanoparticles exhibited high antimicrobial activities against bacteria and phytopathogenic fungi. ZnO NPs displayed better anti-fungal properties than TiO
2. The most effective nanoparticles for antimicrobial properties typically have diameters in the range of 10–100 nm. Both ZnO NPs and CuO NPs have shown broad-spectrum antimicrobial activities, making them promising candidates for various applications, including antiviral, anticancer, and UV-protection [
81,
82]. ZnO-NPs have diverse applications, including sensors, drug delivery, wastewater treatment, tissue engineering, catalysts, and antimicrobials. Sustainable synthesis methods are crucial for widespread use [
83].
In terms of the phenolic compounds, the Fe
3O
4 NPs caused variable effects on the phenolic compounds in
S. cepivorum-infected onions, with concentrations of 1000 ppm, 1200 ppm, and 1400 ppm leading to fluctuations in the total, free, and related phenols. Cai et al. [
84] found that Fe
3O
4 NPs, when foliar exposed to
Nicotiana benthamiana, demonstrate evidence of nanoparticle uptake, acting as both plant growth promoters and elicitors of defense responses against plant viruses. Also, Fe
3O
4 NPs have increased phenolic compound levels [
63]. The Cu NPs and ZnO NPs minimally impacted the phenolic compound levels in infected onions, aligning with studies on
Hypericum perforatum L. using various metal oxide nanoparticles [
85]. The combination of salicylic acid and Fe
3O
4 NPs yielded concentration-dependent effects on the phenolic compounds, reflecting the complex interactions between salicylic acid, nanoparticles, and the plant–pathogen system. The control group, representing infected and uninfected onions without nanoparticle treatment, exhibited the lowest phenolic compound levels, indicating that
S. cepivorum infection negatively influences their synthesis or accumulation due to plant–pathogen interactions [
86].
The three tested inducers, salicylic acid, PDP, and ascorbic acid, exhibited potential impacts on disease management and plant growth. Salicylic acid, at concentrations of 250 ppm, 500 ppm, and 1000 ppm, demonstrated promising results in decreasing both the disease incidence and severity. These findings align with previous studies highlighting salicylic acid’s role in stimulating plant defensive mechanisms versus various pathogens [
87,
88]. Salicylic acid is known to activate systemic acquired resistance (SAR) in plants, which enhances their ability to ward off infections [
88]. The increase in the salicylic acid concentration resulted in a decline in the disease severity, indicating a concentration-dependent response, in line with Thaler et al. [
89]. Salicylic acid is fundamental in regulating plant defense responses [
90]. The positive impact of salicylic acid on plant growth, as indicated by the increased fresh and dry weights, is consistent with studies demonstrating its involvement in promoting overall plant health and development [
91].
The application of SA has shown positive effects on the growth vigor of both root and shoot [
92]. This improvement is attributed to hormonal regulation, increased leaf turgidity through stomatal closure, reduced transpiration rates, and enhanced relative water content [
93]. As a phenolic phytohormone, SA acts as a systemic signal that triggers the local systemic resistance response. It serves as a key regulator in the signaling network for plants facing abiotic and biotic stresses, leading to the increased activity of the PAL, PO, and PPO enzymes, along with elevated phenolic contents. The exogenous supplementation of SA stimulates systemic resistance and enhances plant performance under both abiotic and biotic stress [
94,
95,
96].
PDP exhibited mixed effects on the disease severity and plant growth. This variation in response might be due to the complex interaction between phosphorus, potassium, and disease resistance in plants [
97]. Potassium is known to enhance plant resistance toward diverse phytopathogens [
98,
99]. The positive impact of higher PDP concentrations on the plant growth parameters, despite the presence of the disease, is consistent with studies indicating the role of phosphorus and potassium in enhancing plant vigor and tolerance to stress [
100]. However, the mixed effects on the disease assessment suggest that PDP may have a more nuanced impact on the specific pathogen–host interaction under consideration.
Ascorbic acid (AsA) demonstrated limited efficacy in reducing the disease incidence and severity. The consistently high disease incidence and severity, even at higher concentrations of ascorbic acid, indicate that it may not be as effective in managing onion white rot disease. However, ascorbic acid has demonstrated effectiveness in minimizing conidia numbers’ impact and powdery mildew severity on flower clusters and fruit set [
101]. Similar findings have been reported in studies on other plant–pathogen systems, where the effectiveness of ascorbic acid as a disease control agent varies [
102,
103]. The slight improvements in the plant growth parameters with increasing ascorbic acid concentrations may suggest a potential role in promoting plant health. Still, the limited impact on disease control raises questions about its efficacy as a standalone control measure for this specific disease [
104,
105,
106,
107].
Ascorbic acid is a crucial plant antioxidant, acting as a frontline defense against ROS induced by environmental factors like wounding, ozone, high salinity, and pathogens [
105,
107]. Its role extends to plant–pathogen interactions, contributing to the overall plant growth, development, and stress tolerance. As part of a complex defense network, AsA, along with glutathione and enzymatic antioxidants, maintains a redox environment essential for regulating defense pathways. These pathways involve the activation of the NPR1 regulatory transcription factor, expression of defense genes, cell wall reinforcement, and modulation of defense–hormonal signaling networks. AsA also acts as an inducer or part of the induced resistance processes triggered by elicitors like β-aminobutyric acid, jasmonic acid, methyljasmonate, and extracellular polysaccharides [
104,
105,
106,
107].
The varying effects of PDP on the phenolic compounds at different concentrations highlight the complex interplay between nutrient availability and plant defense responses. The decrease in the total phenols and related phenols at 1000 ppm may be attributed to the intricate balance between phosphorus and potassium, where an excess of one nutrient may affect the plant’s ability to mount an effective defense [
108]. Moreover, the consistent increase in the phenolic compounds with ascorbic acid supplementation aligns with the antioxidant properties of ascorbic acid, which can stimulate the synthesis of secondary metabolites, including phenolics [
109]. The elevated levels of the phenolic compounds, particularly at 1000 ppm, suggest that ascorbic acid positively influences the induction of plant defense mechanisms against
S. cepivorum.
In turn, the results highlight the effectiveness of Bellis, Topsin M, and BOGARD in inhibiting the linear growth of
S. cepivorum. This aligns with the existing literature highlighting the efficacy of Bellis
® in controlling fungal pathogens [
110,
111,
112]. Similarly, Topsin M demonstrated potent antifungal activity against various pathogens [
113,
114]. Moreover, other findings have been reported, emphasizing BOGARD’s strong antifungal properties [
115]. Fungicides, such as those tested in this study, are known to interfere with crucial cellular processes in fungi, including cell wall synthesis and energy production [
112,
114]. The observed effectiveness of the tested fungicides in inhibiting the linear growth of
S. cepivorum can be attributed to their distinct active ingredients and modes of action [
116]. Bellis
® contains a combination of pyraclostrobin and boscalid. Pyraclostrobin is a strobilurin fungicide that disrupts fungal respiration by inhibiting the electron transport chain in mitochondria, decreasing ATP production [
117,
118]. Conversely, boscalid acts on the succinate dehydrogenase enzyme, disrupting the tricarboxylic acid cycle in fungi [
118,
119]. The dual action of pyraclostrobin and boscalid disrupts multiple cellular processes in the fungus, resulting in a potent antifungal effect [
120]. This combination likely contributes to the high reduction percentages observed in the linear growth of
S. cepivorum. Topsin M contains thiophanate methyl, which belongs to the benzimidazole class of fungicides. Thiophanate methyl inhibits microtubule assembly in fungi, disrupting cell division and growth [
121,
122]. The observed reduction in the linear growth of
S. cepivorum can be attributed to the disruption of cell division processes by thiophanate methyl [
123]. The fungicidal activity of this benzimidazole likely hampers the pathogen’s ability to proliferate and spread [
124]. As for the BOGARD, it contains tebuconazole, a triazole fungicide. Triazoles interfere with the process of producing ergosterol, which is necessary for the membranes of fungi, leading to membrane destabilization and cell death [
125]. The inhibition of ergosterol biosynthesis disrupts the integrity of the fungal cell membrane, compromising its structural integrity and function [
126]. This likely contributes to the complete growth inhibition observed at lower concentrations of BOGARD. Understanding the specific modes of action of these fungicides provides insights into their effectiveness against
S. cepivorum. The combination of multiple active ingredients in Bellis
®, the microtubule disruption by the thiophanate methyl in Topsin M, and the membrane-targeting action of the tebuconazole in BOGARD collectively contribute to the potent antifungal effects observed in the study.
This study offers insightful information on the complex reactions of onion plants to different fungicides and concentrations. While Bellis
® demonstrated stability in the chlorophyll content, Topsin M and BOGARD showed concentration-dependent trends. These findings highlight the importance of optimizing the fungicide concentrations for effective disease management while minimizing potential physiological stress in plants. The chlorophyll levels in Bellis
® treatments suggest that it exerts minimal direct stress on the photosynthetic apparatus. Similar findings have been reported in studies assessing the impact of fungicides on the chlorophyll content of various crops [
112,
127]. The absence of a clear concentration-dependent response might indicate that it primarily functions as a disease control agent without causing significant physiological disruptions. Also, some studies indicated that certain fungicides, including those containing thiophanate methyl, as in Topsin M, can induce stress in the photosynthetic apparatus at higher concentrations [
128,
129]. The decrease in the chlorophyll levels may be attributed to the fungicide’s impact on cellular processes involved in photosynthesis, emphasizing the importance of optimizing fungicide concentrations to avoid adverse effects on plant health [
129]. The observed variability suggests that while lower concentrations may have a minimal impact on chlorophyll, higher concentrations can lead to physiological stress. This emphasizes the importance of carefully calibrating the fungicide doses for optimal disease control and minimal impact on plant physiology [
130].
In addition, the consistent increase in phenols across all the fungicide treatments suggests a systemic induction of phenolic compounds, indicative of an enhanced defense response against the pathogen [
131]. Notably, the tested fungicides demonstrated a dose-dependent increase in the phenolic compounds, emphasizing their potential to induce the synthesis of secondary metabolites involved in plant defense mechanisms. The control groups exhibited significantly lower phenolic levels, with the positive control (+) indicating the detrimental impact of the pathogen and the negative control (−) highlighting the inherent baseline stress response induced by the plant immune system [
132]. This consistent increase in the phenolic compounds aligns with the current understanding of plant defense responses, where the activation of secondary metabolites, including phenolics, is a common strategy to combat pathogenic attacks [
133]. The findings suggest that the tested fungicides contribute to disease management and induce a systemic defense response in the form of increased phenolic compounds, potentially fortifying the plants against further pathogenic infection.






{kind=link}
{kind=link}
{kind=link}