
Influence of Mutations and N-Glycosylation Sites in the Receptor-Binding Domain (RBD) and the Membrane Protein of SARS-CoV-2 Variants of Concern on Antibody Binding in ELISA

 , , , , , ,
, , , , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum Collection
2.2. Recombinant Proteins Expressed in E. coli
2.3. Recombinant Proteins Expressed in HEK Cells
2.4. Glycostaining
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. FASP Digestion
2.7. Lectin Enrichment
2.8. Mass Spectrometry
2.9. Database Search
2.10. Statistical Analysis
3. Results
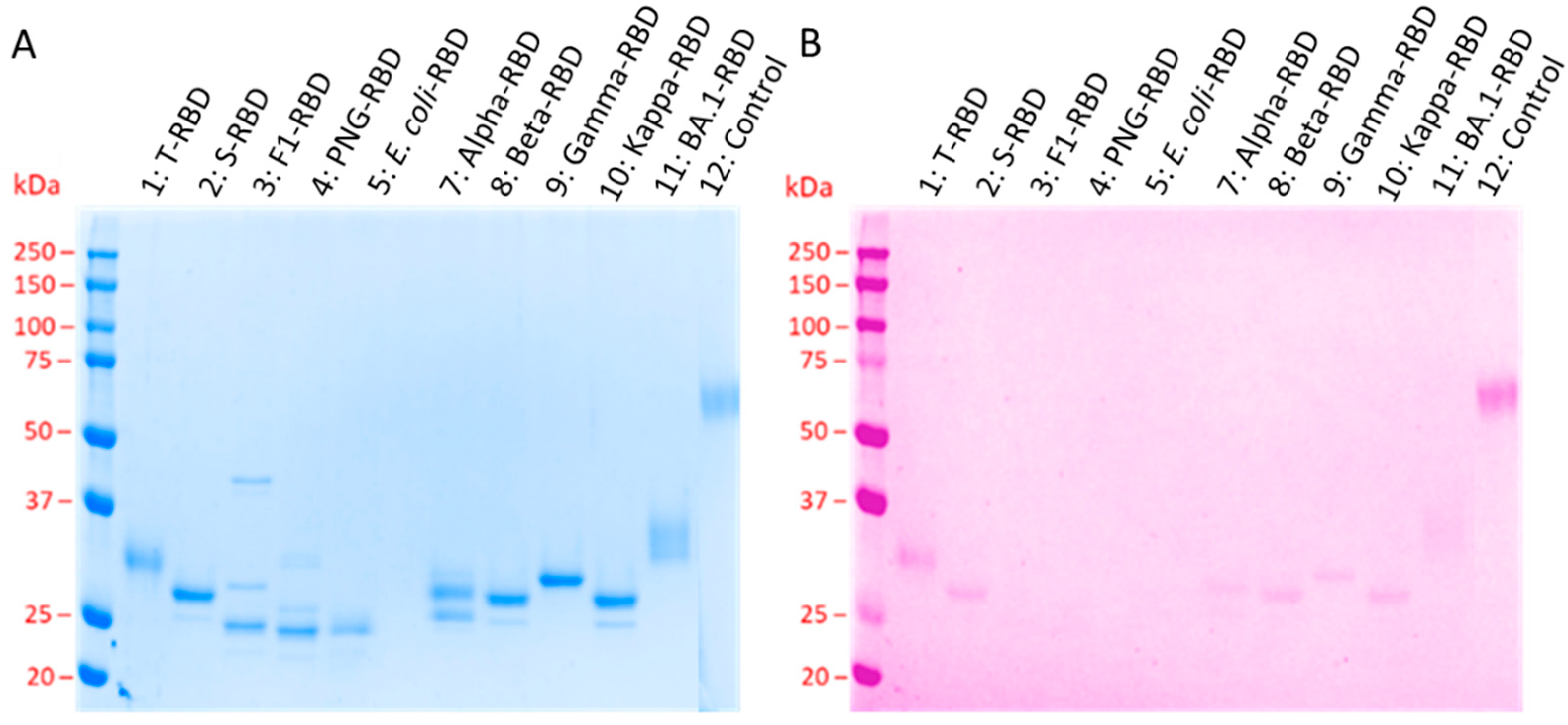
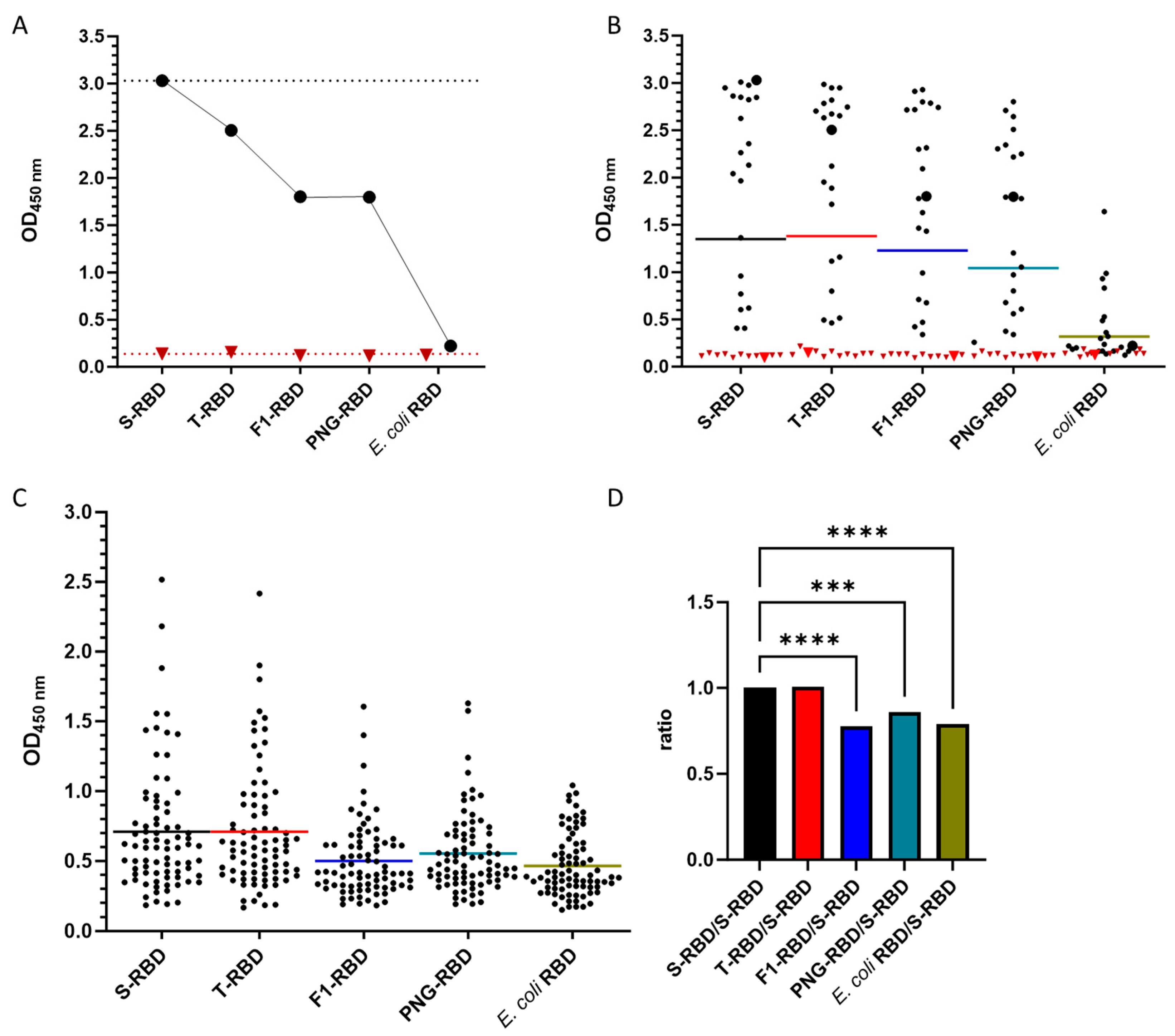
3.1. Confirmation of RBD Glycosylation
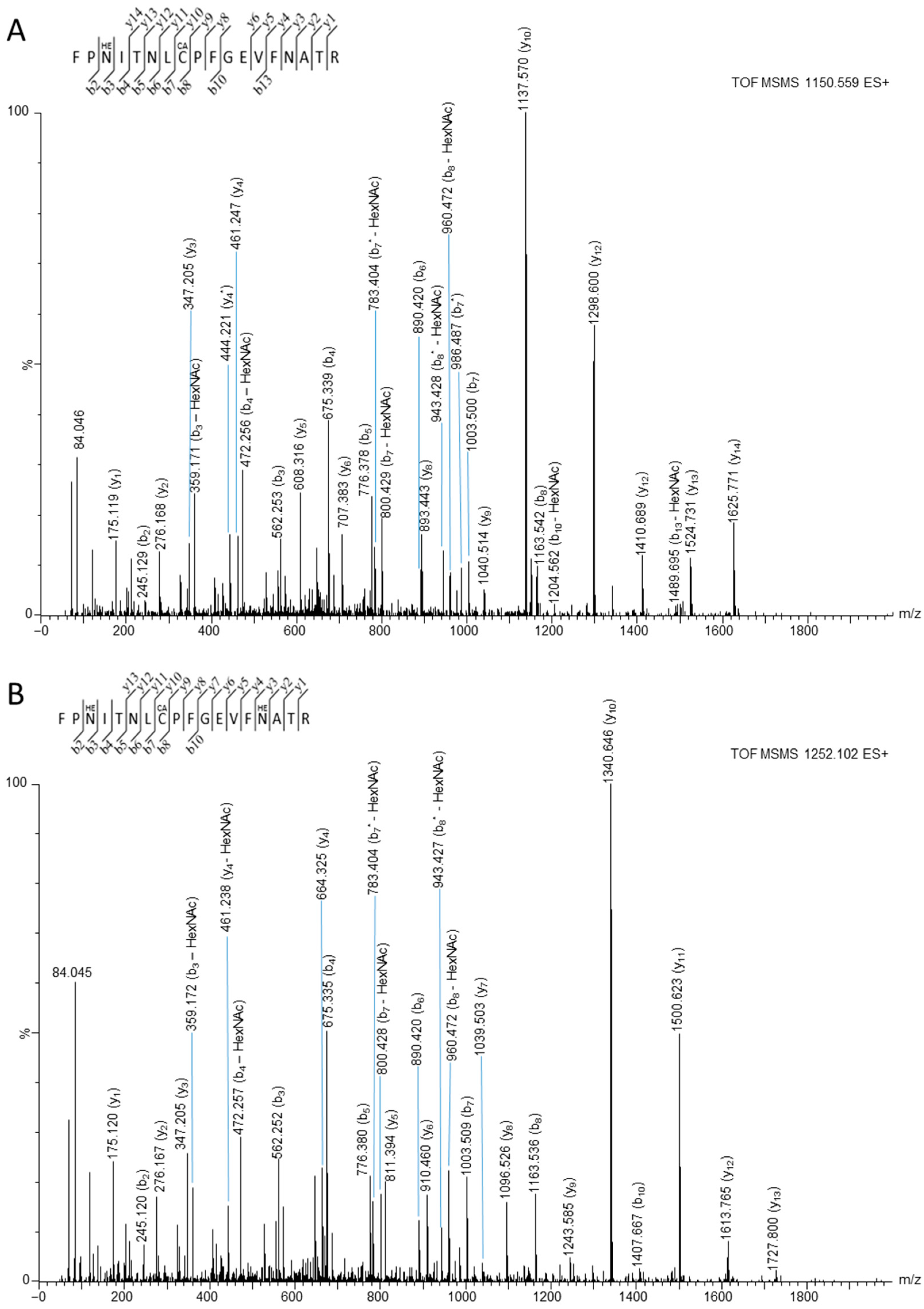
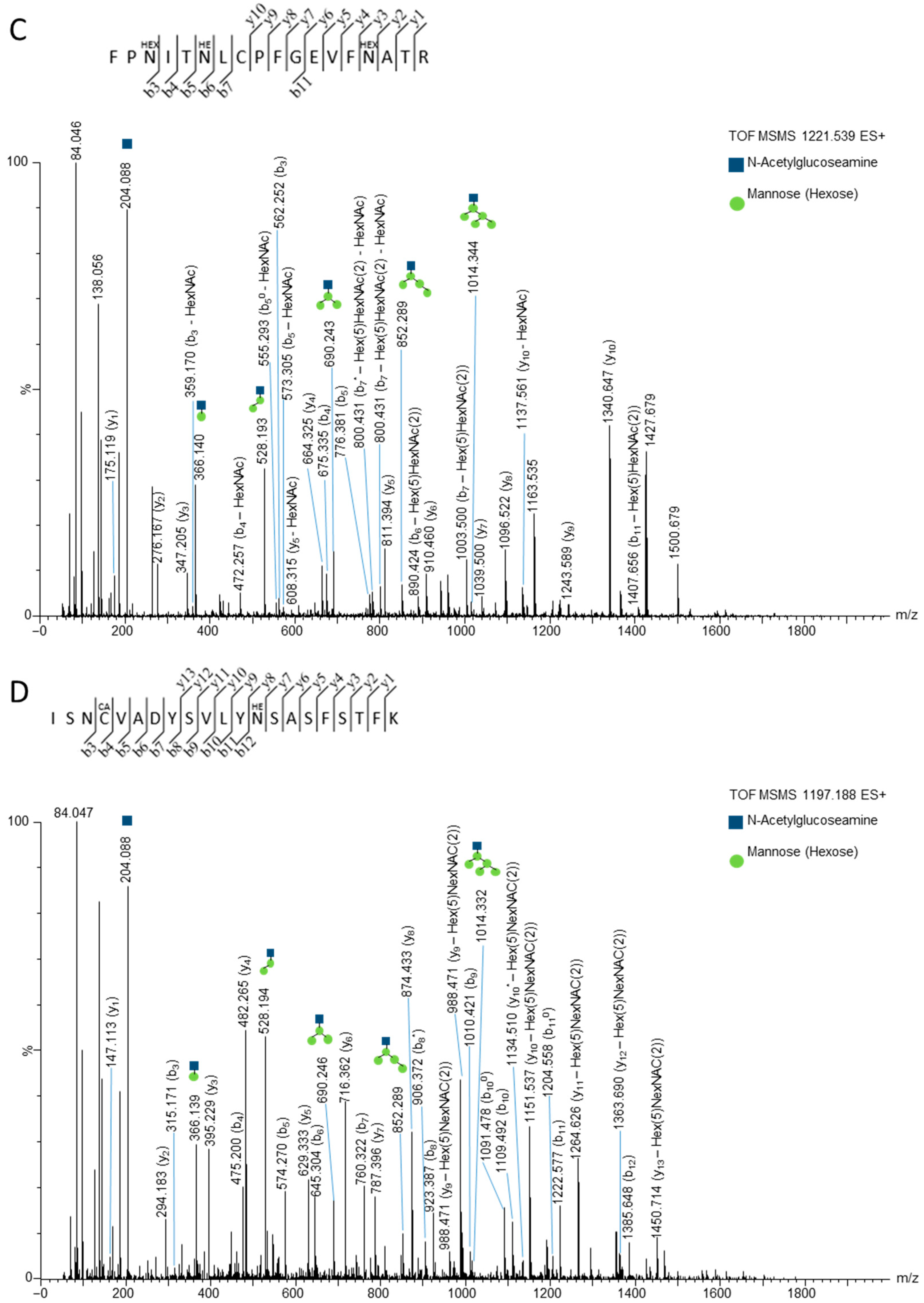
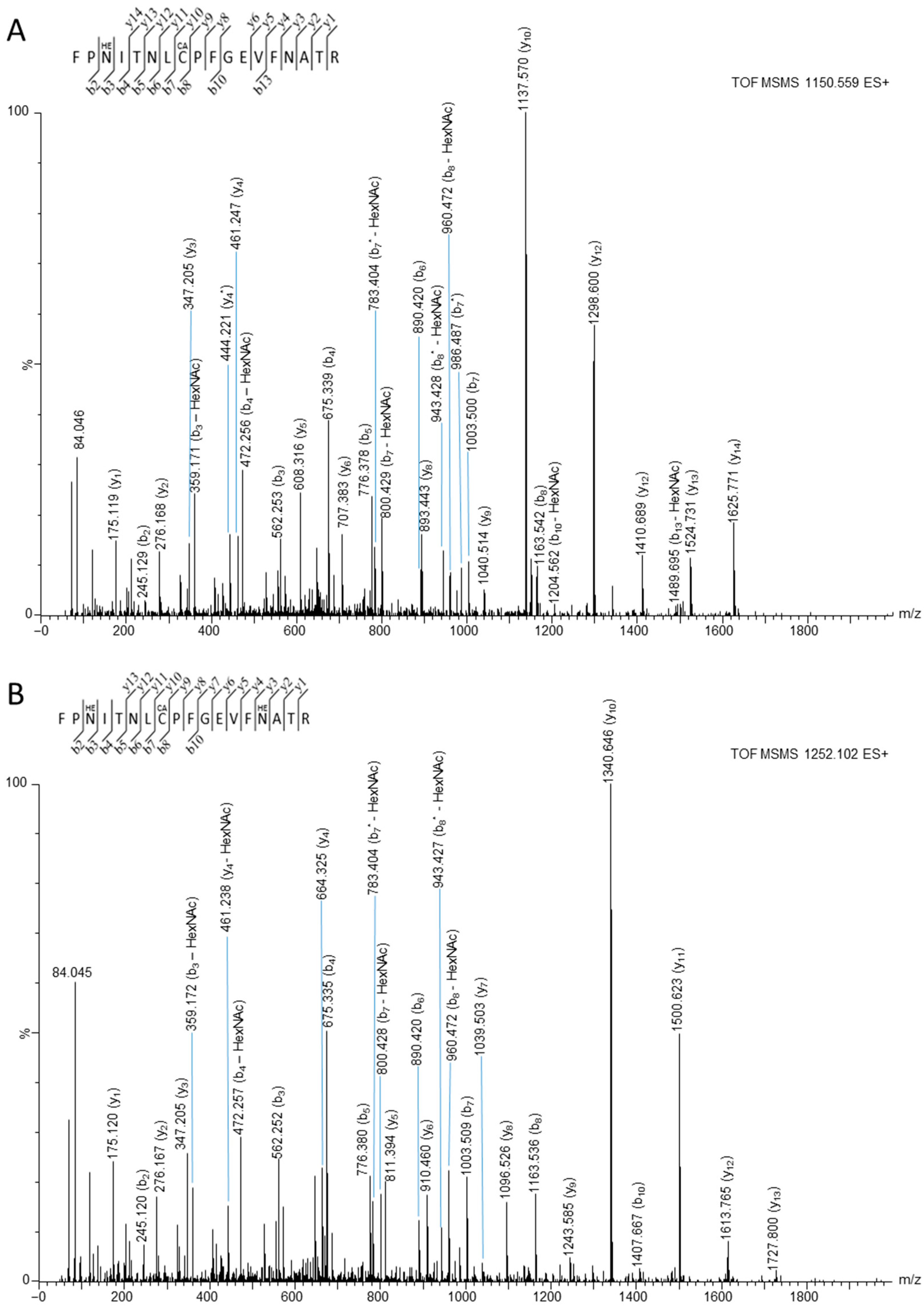
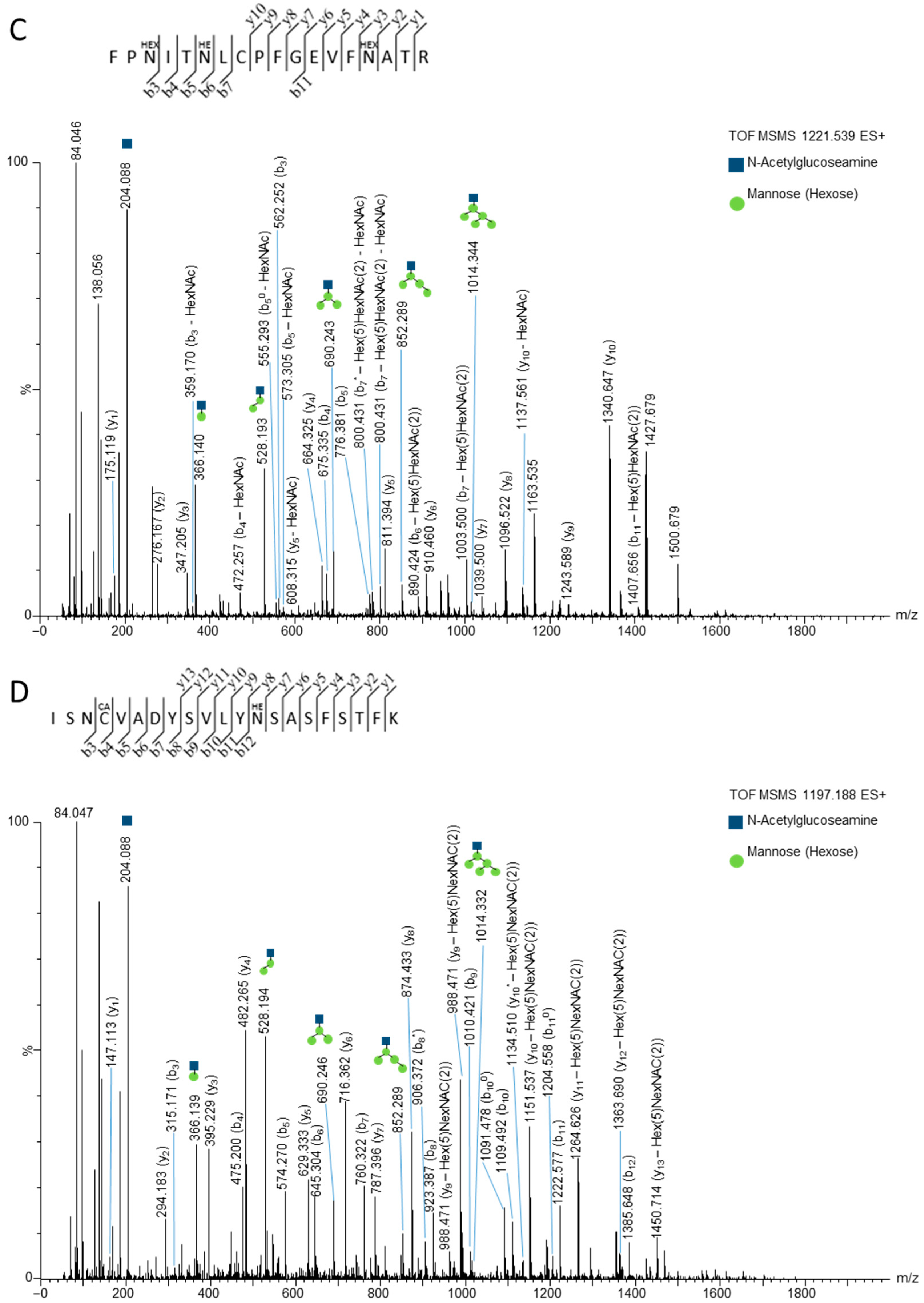
3.2. Mass Spectrometry Studies
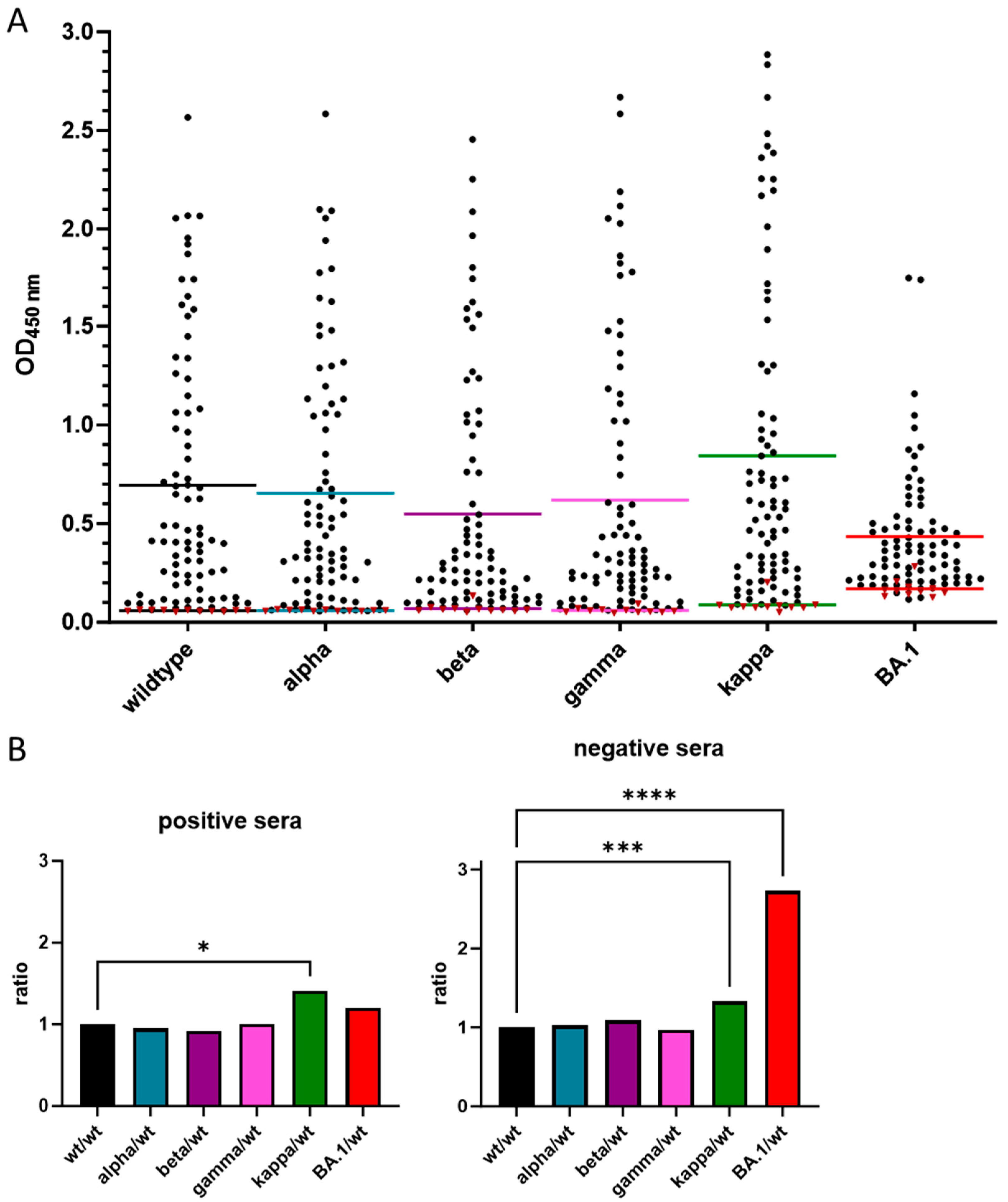
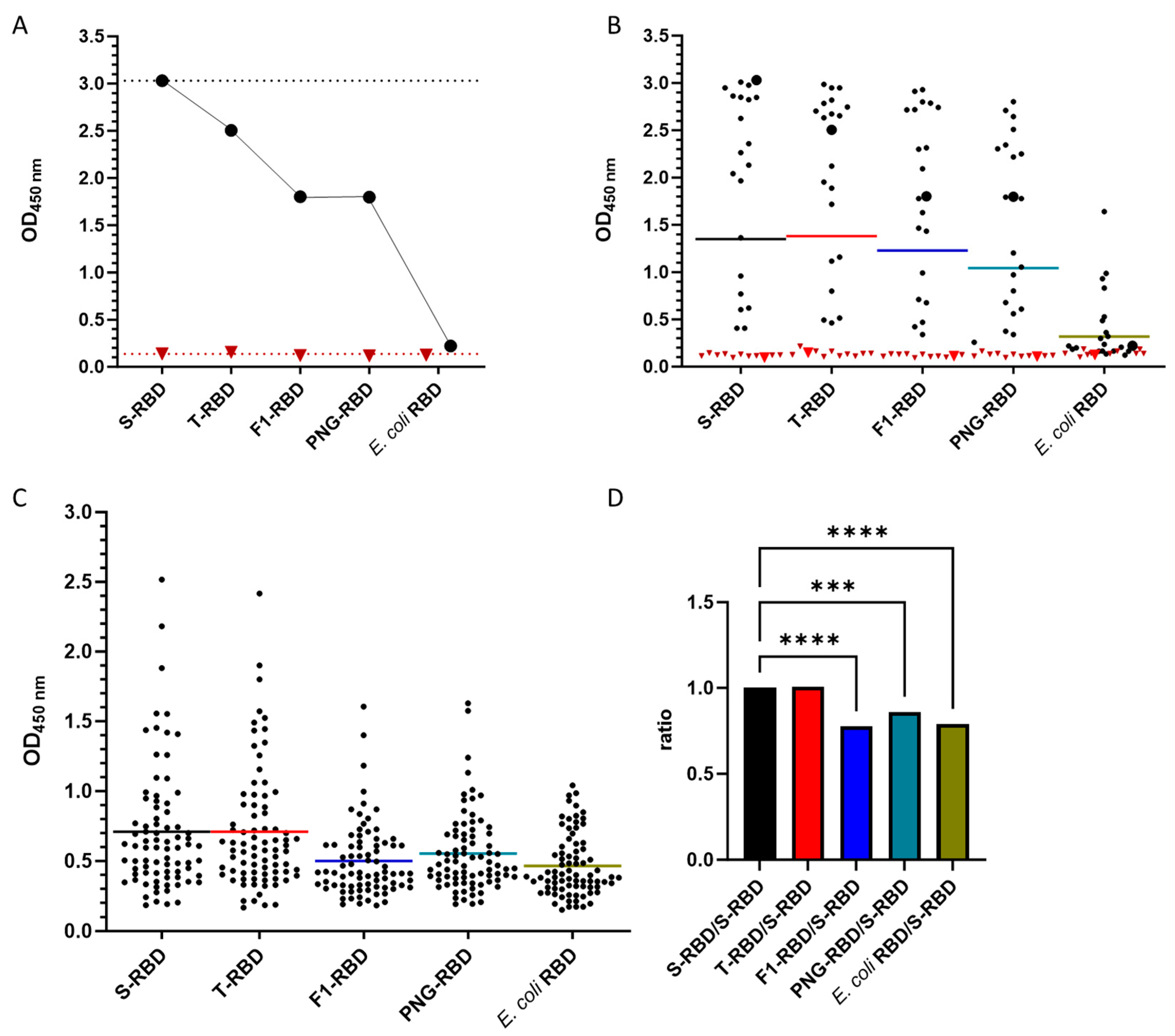
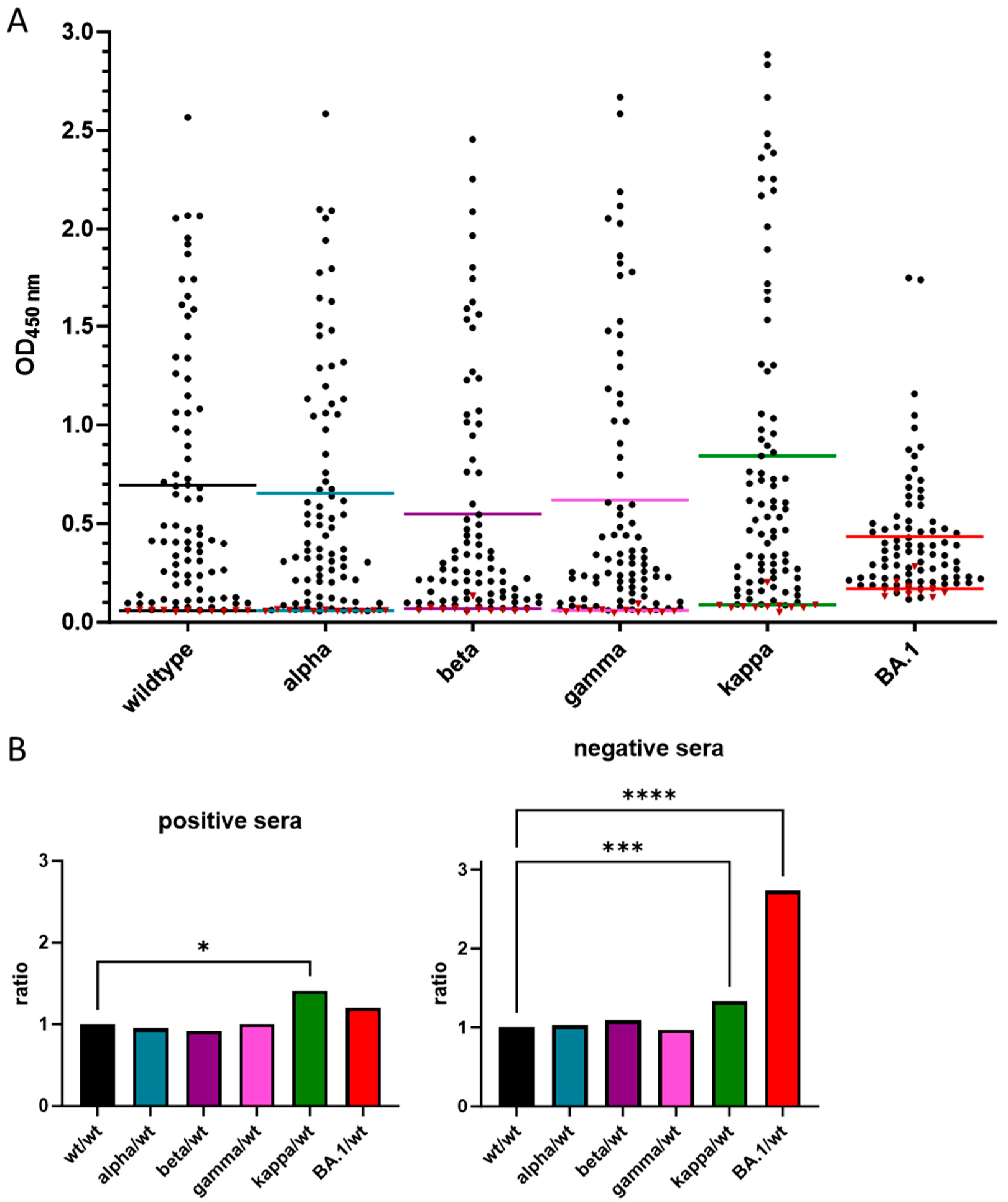
3.3. Cross-Reactivity of Anti-RBD IgG Antibodies against Different VOCs
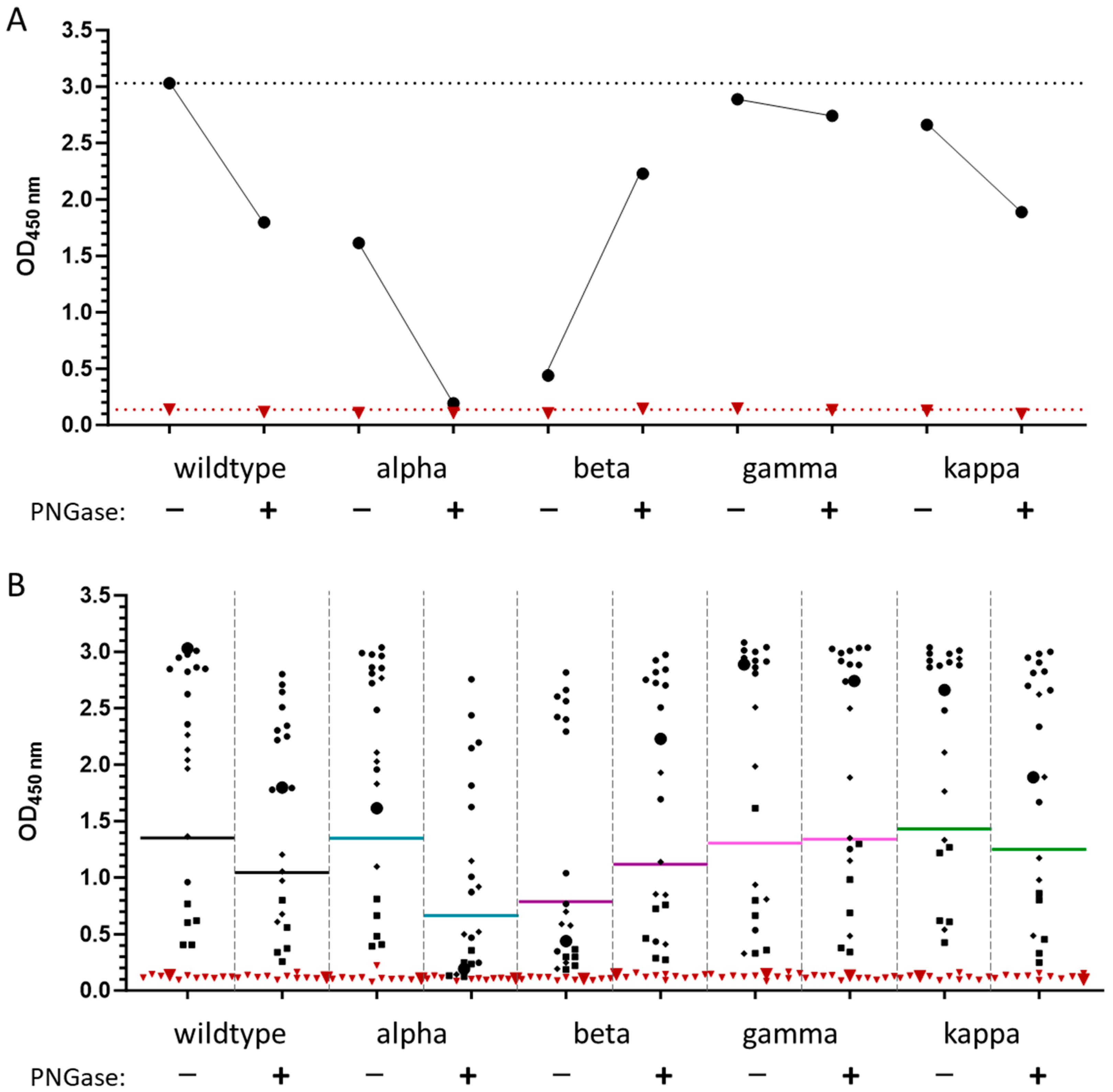
3.4. Effect of Glycosylation of wt-RBD on IgG Recognition
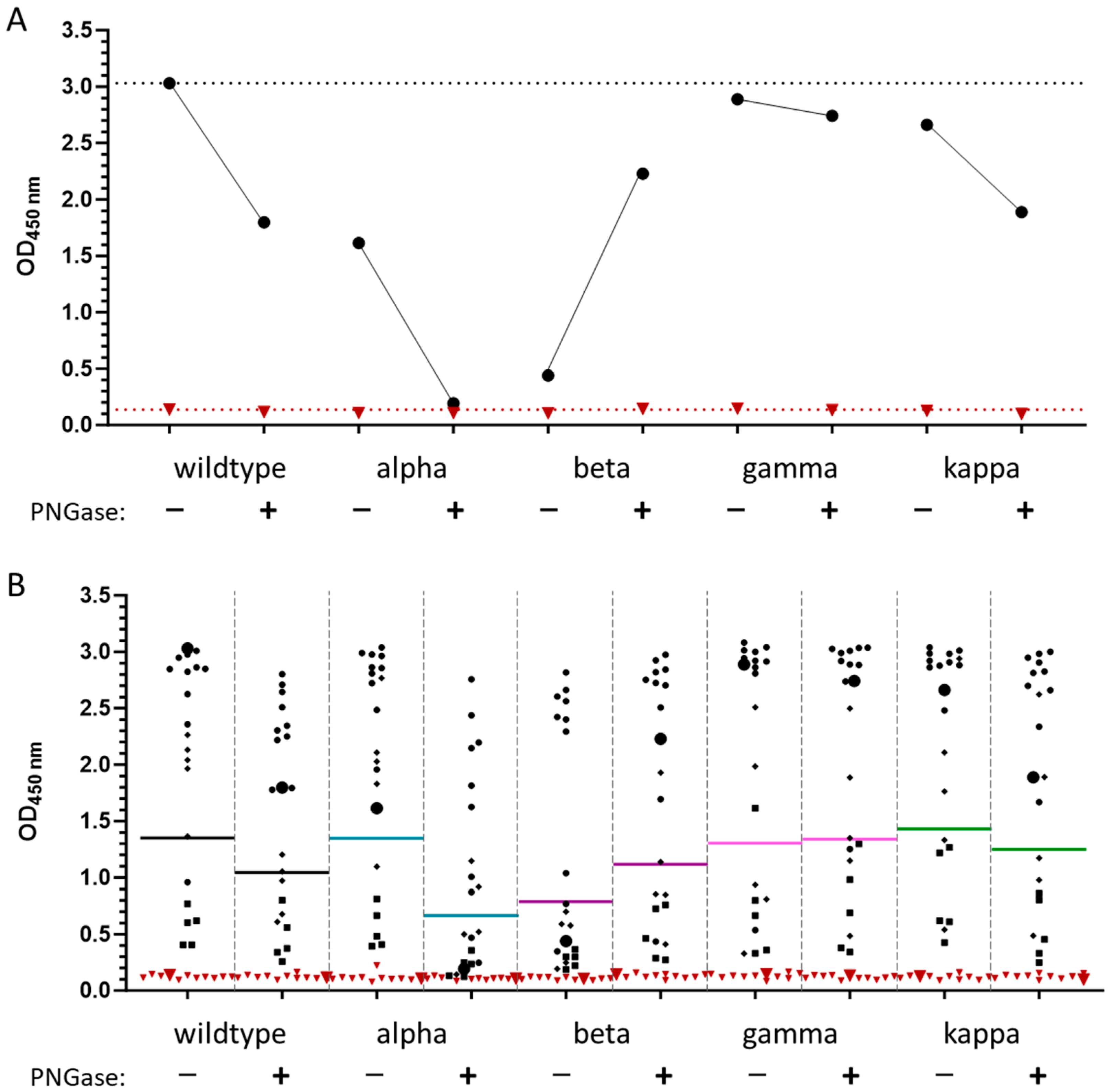
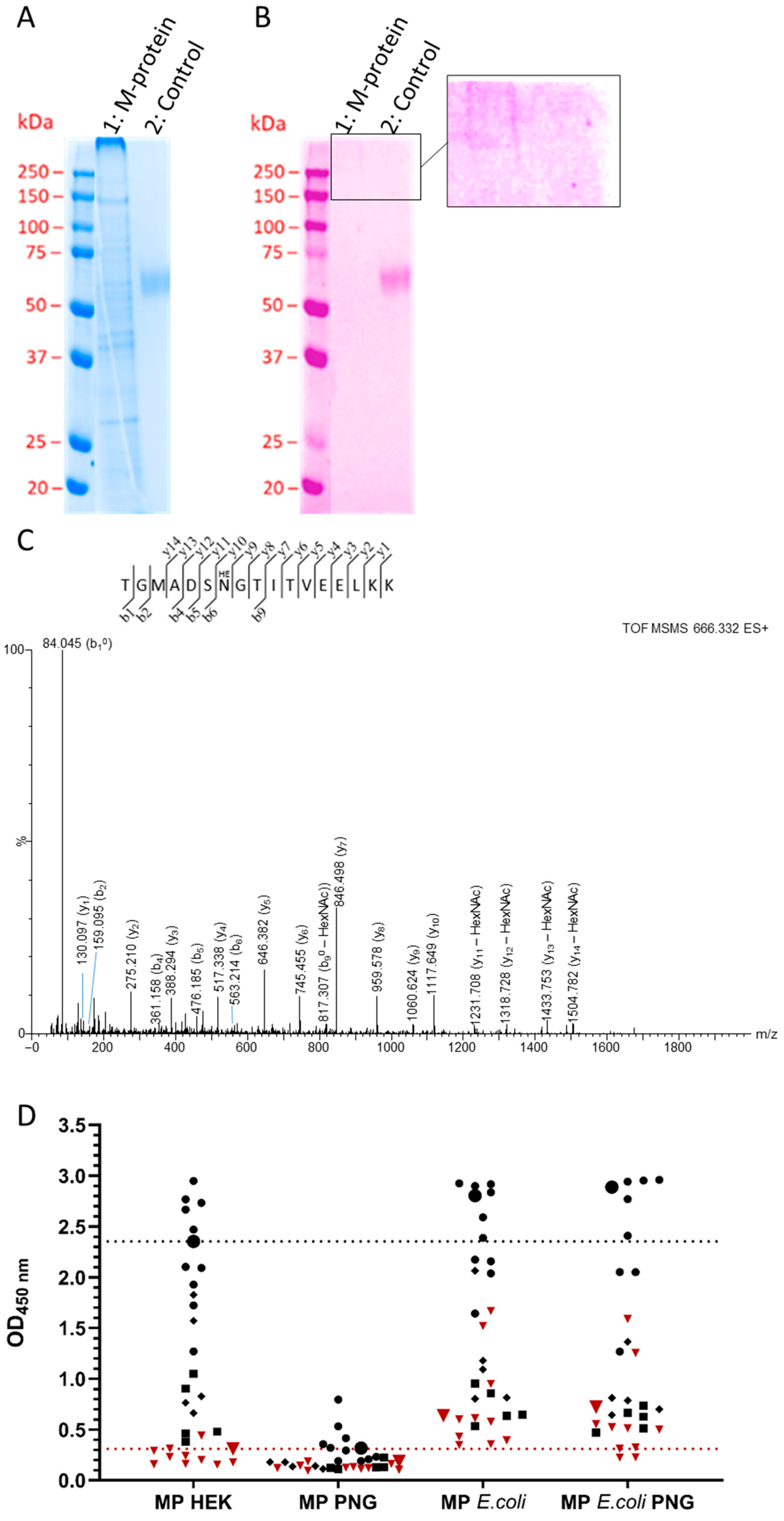
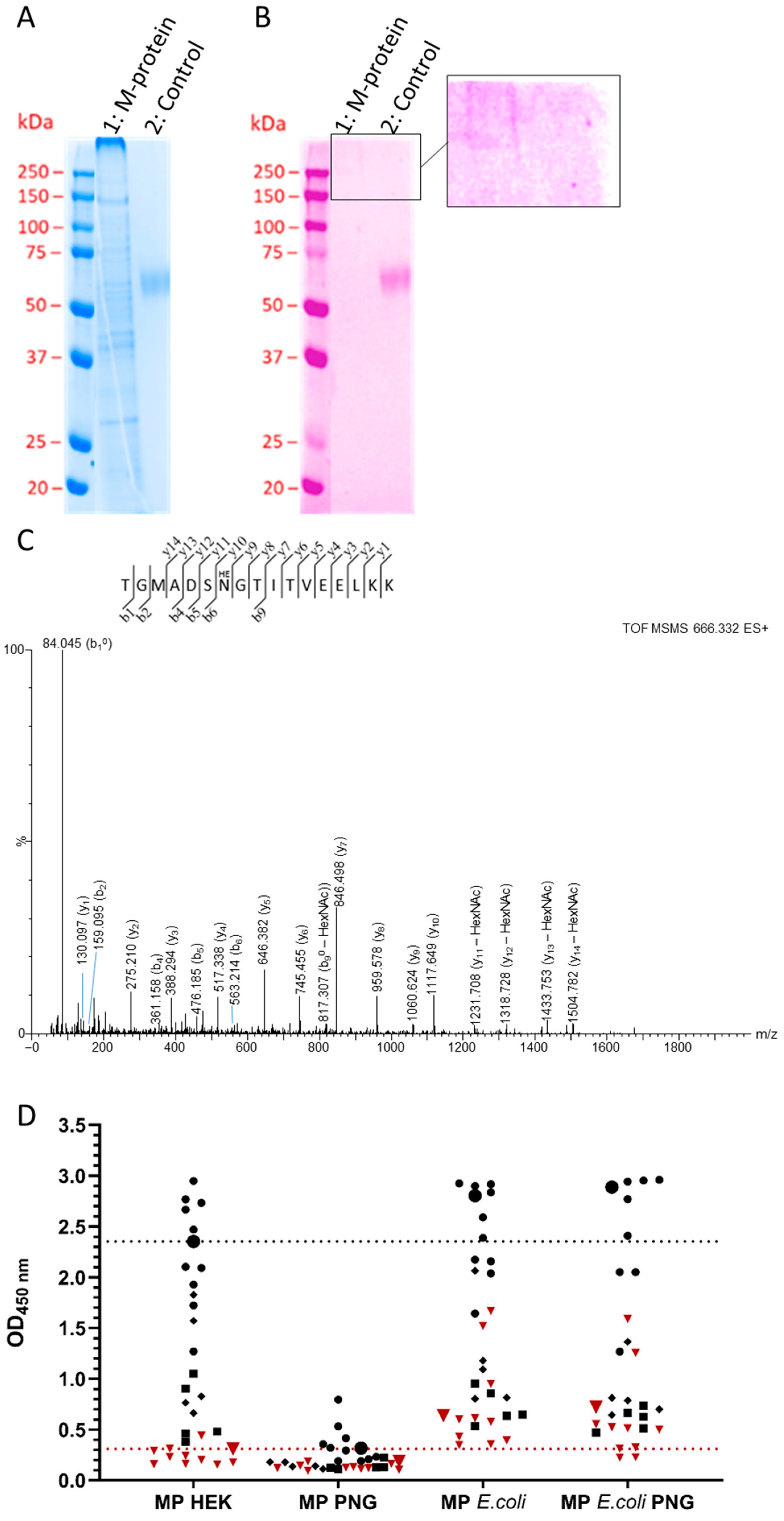
3.5. Membrane Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajpoot, S.; Ohishi, T.; Kumar, A.; Pan, Q.; Banerjee, S.; Zhang, K.Y.J.; Baig, M.S. A Novel Therapeutic Peptide Blocks SARS-CoV-2 Spike Protein Binding with Host Cell ACE2 Receptor. Drugs R D 2021, 21, 273–283. [Google Scholar] [CrossRef]
- Chathappady House, N.N.; Palissery, S.; Sebastian, H. Corona Viruses: A Review on SARS, MERS and COVID-19. Microbiol. Insights 2021, 14, 11786361211002481. [Google Scholar] [CrossRef]
- Casalino, L.; Gaieb, Z.; Goldsmith, J.A.; Hjorth, C.K.; Dommer, A.C.; Harbison, A.M.; Fogarty, C.A.; Barros, E.P.; Taylor, B.C.; McLellan, J.S.; et al. Beyond Shielding: The Roles of Glycans in the SARS-CoV-2 Spike Protein. ACS Cent. Sci. 2020, 6, 1722–1734. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Qin, S.; Dai, L.; Tian, Z. The glycosylation in SARS-CoV-2 and its receptor ACE2. Signal Transduct. Target. Ther. 2021, 6, 396. [Google Scholar] [CrossRef] [PubMed]
- Dolan, K.A.; Dutta, M.; Kern, D.M.; Kotecha, A.; Voth, G.A.; Brohawn, S.G. Structure of SARS-CoV-2 M protein in lipid nanodiscs. eLife 2022, 11, e81702. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Nomura, N.; Muramoto, Y.; Ekimoto, T.; Uemura, T.; Liu, K.; Yui, M.; Kono, N.; Aoki, J.; Ikeguchi, M.; et al. Structure of SARS-CoV-2 membrane protein essential for virus assembly. Nat. Commun. 2022, 13, 4399. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Toussi, S.S.; Hammond, J.L.; Gerstenberger, B.S.; Anderson, A.S. Therapeutics for COVID-19. Nat. Microbiol. 2023, 8, 771–786. [Google Scholar] [CrossRef]
- Pfizer. Pfizer Receives U.S. FDA Emergency Use Authorization for Novel COVID-19 Oral Antiviral Treatment (22 December 2021). Available online: https://go.nature.com/3KdyjBk (accessed on 5 January 2024).
- Gote, V.; Bolla, P.K.; Kommineni, N.; Butreddy, A.; Nukala, P.K.; Palakurthi, S.S.; Khan, W. A Comprehensive Review of mRNA Vaccines. Int. J. Mol. Sci. 2023, 24, 2700. [Google Scholar] [CrossRef]
- Aloor, A.; Aradhya, R.; Venugopal, P.; Gopalakrishnan Nair, B.; Suravajhala, R. Glycosylation in SARS-CoV-2 variants: A path to infection and recovery. Biochem. Pharmacol. 2022, 206, 115335. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Lai, Y.-J.; Liao, C.-C.; Yang, W.-F.; Huang, K.-B.; Lee, I.-J.; Chou, W.-C.; Wang, S.-H.; Wang, L.-H.; Hsu, J.-M.; et al. Targeting conserved N-glycosylation blocks SARS-CoV-2 variant infection in vitro. eBioMedicine 2021, 74, 103712. [Google Scholar] [CrossRef]
- Hargett, A.A.; Renfrow, M.B. Glycosylation of viral surface proteins probed by mass spectrometry. Curr. Opin. Virol. 2019, 36, 56–66. [Google Scholar] [CrossRef]
- Fung, T.S.; Liu, D.X. Post-translational modifications of coronavirus proteins: Roles and function. Future Virol. 2018, 13, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Parsons, L.M.; Jankowska, E.; Cipollo, J.F. Site-Specific Glycosylation Patterns of the SARS-CoV-2 Spike Protein Derived from Recombinant Protein and Viral WA1 and D614G Strains. Front. Chem. 2021, 9, 767448. [Google Scholar] [CrossRef]
- Li, Q.; Wu, J.; Nie, J.; Zhang, L.; Hao, H.; Liu, S.; Zhao, C.; Zhang, Q.; Liu, H.; Nie, L.; et al. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity. Cell 2020, 182, 1284–1294.e9. [Google Scholar] [CrossRef]
- Shajahan, A.; Pepi, L.E.; Kumar, B.; Murray, N.B.; Azadi, P. Site specific N- and O-glycosylation mapping of the spike proteins of SARS-CoV-2 variants of concern. Nature 2023, 13, 10053. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Baudys, J.; Osman, S.H.; Barr, J.R. Analysis of the N-glycosylation profiles of the spike proteins from the Alpha, Beta, Gamma, and Delta variants of SARS-CoV-2. Anal. Bioanal. Chem. 2023, 415, 4779–4793. [Google Scholar] [CrossRef]
- Amrun, S.N.; Lee, C.Y.-P.; Lee, B.; Fong, S.-W.; Young, B.E.; Chee, R.S.-L.; Yeo, N.K.-W.; Torres-Ruesta, A.; Carissimo, G.; Poh, C.M.; et al. Linear B-cell epitopes in the spike and nucleocapsid proteins as markers of SARS-CoV-2 exposure and disease severity. eBioMedicine 2020, 58, 102911. [Google Scholar] [CrossRef]
- Wang, H.; Wu, X.; Zhang, X.; Hou, X.; Liang, T.; Wang, D.; Teng, F.; Dai, J.; Duan, H.; Guo, S.; et al. SARS-CoV-2 Proteome Microarray for Mapping COVID-19 Antibody Interactions at Amino Acid Resolution. ACS Cent. Sci. 2020, 6, 2238–2249. [Google Scholar] [CrossRef]
- Grant, O.C.; Montgomery, D.; Ito, K.; Woods, R.J. Analysis of the SARS-CoV-2 spike protein glycan shield reveals implications for immune recognition. Sci. Rep. 2020, 10, 14991. [Google Scholar] [CrossRef]
- Engel, C.; Wirkner, K.; Zeynalova, S.; Baber, R.; Binder, H.; Ceglarek, U.; Enzenbach, C.; Fuchs, M.; Hagendorff, A.; Henger, S.; et al. Cohort Profile: The LIFE-Adult-Study. Int. J. Epidemiol. 2022, 52, e66–e79. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Engel, C.; Ahnert, P.; Alfermann, D.; Arelin, K.; Baber, R.; Beutner, F.; Binder, H.; Brähler, E.; Burkhardt, R.; et al. The LIFE-Adult-Study: Objectives and design of a population-based cohort study with 10,000 deeply phenotyped adults in Germany. BMC Public Health 2015, 15, 691. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, M.; Luo, J.; Brakel, A.; Krizsan, A.; Lakowa, N.; Grünewald, T.; Lehmann, C.; Wolf, J.; Borte, S.; Milkovska-Stamenova, S.; et al. Evaluation of S- and M-Proteins Expressed in Escherichia coli and HEK Cells for Serological Detection of Antibodies in Response to SARS-CoV-2 Infections and mRNA-Based Vaccinations. Pathogens 2022, 11, 1515. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, M.; Krizsan, A.; Brakel, A.; Pohl, F.; Volke, D.; Hoffmann, R. Cross-Reactivity of IgG Antibodies and Virus Neutralization in mRNA-Vaccinated People Against Wild-Type SARS-CoV-2 and the Five Most Common SARS-CoV-2 Variants of Concern. Front. Immunol. 2022, 13, 915034. [Google Scholar] [CrossRef]
- Milkovska-Stamenova, S.; Wölk, M.; Hoffmann, R. Evaluation of Sample Preparation Strategies for Human Milk and Plasma Proteomics. Molecules 2021, 26, 6816. [Google Scholar] [CrossRef] [PubMed]
- Deeb, S.J.; Cox, J.; Schmidt-Supprian, M.; Mann, M. N-linked glycosylation enrichment for in-depth cell surface proteomics of diffuse large B-cell lymphoma subtypes. Mol. Cell. Proteom. 2014, 13, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; Aebi, M.; Darvill, A.G.; Kinoshita, T.; Packer, N.H.; Prestegard, J.H.; et al. (Eds.) Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015. [Google Scholar]
- Reeves, P.J.; Callewaert, N.; Contreras, R.; Khorana, H.G. Structure and function in rhodopsin: High-level expression of rhodopsin with restricted and homogeneous N-glycosylation by a tetracycline-inducible N-acetylglucosaminyltransferase I-negative HEK293S stable mammalian cell line. Proc. Natl. Acad. Sci. USA 2002, 99, 13419–13424. [Google Scholar] [CrossRef] [PubMed]
- Shajahan, A.; Supekar, N.T.; Gleinich, A.S.; Azadi, P. Deducing the N- and O-glycosylation profile of the spike protein of novel coronavirus SARS-CoV-2. Glycobiology 2020, 30, 981–988. [Google Scholar] [CrossRef]
- Elder, J.H.; Alexander, S. endo-beta-N-acetylglucosaminidase F: Endoglycosidase from Flavobacterium meningosepticum that cleaves both high-mannose and complex glycoproteins. Proc. Natl. Acad. Sci. USA 1982, 79, 4540–4544. [Google Scholar] [CrossRef]
- Masuda, A.; Xu, J.; Mitsudome, T.; Nagata, Y.; Morokuma, D.; Mon, H.; Banno, Y.; Kusakabe, T.; Lee, J.M. Mass Production of an Active Peptide-N-Glycosidase F Using Silkworm-Baculovirus Expression System. Mol. Biotechnol. 2015, 57, 735–745. [Google Scholar] [CrossRef]
- An, H.J.; Froehlich, J.W.; Lebrilla, C.B. Determination of glycosylation sites and site-specific heterogeneity in glycoproteins. Curr. Opin. Chem. Biol. 2009, 13, 421–426. [Google Scholar] [CrossRef]
- Wong, C.-H. Homogeneous Glycoproteins: Overview. In Glycoscience: Biology and Medicine, 1st ed.; Taniguchi, N., Ed.; Springer Reference: Tokyo, Japan, 2015; pp. 959–962. ISBN 978-4-431-54840-9. [Google Scholar]
- Li, Y.; Ma, M.-L.; Lei, Q.; Wang, F.; Hong, W.; Lai, D.-Y.; Hou, H.; Xu, Z.-W.; Zhang, B.; Chen, H.; et al. Linear epitope landscape of the SARS-CoV-2 Spike protein constructed from 1,051 COVID-19 patients. Cell Rep. 2021, 34, 108915. [Google Scholar] [CrossRef] [PubMed]
- Vigan-Womas, I.; Spadoni, J.-L.; Poiret, T.; Taïeb, F.; Randrianarisaona, F.; Faye, R.; Mbow, A.A.; Gaye, A.; Dia, N.; Loucoubar, C.; et al. Linear epitope mapping of the humoral response against SARS-CoV-2 in two independent African cohorts. Sci. Rep. 2023, 13, 782. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Song, Y.; Chen, Y.; Wu, N.; Xu, J.; Sun, C.; Zhang, J.; Weng, T.; Zhang, Z.; Wu, Z.; et al. Molecular Architecture of the SARS-CoV-2 Virus. Cell 2020, 183, 730–738.e13. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.; Vasiljevic, S.; Gangadharan, B.; Hensen, M.; V Chandran, A.; Hill, M.L.; Kiappes, J.L.; Dwek, R.A.; Alonzi, D.S.; Struwe, W.B.; et al. Assessing Antigen Structural Integrity through Glycosylation Analysis of the SARS-CoV-2 Viral Spike. ACS Cent. Sci. 2021, 7, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.; Girgis, M.; Sanda, M. Site-specific glycosylation of SARS-CoV-2: Big challenges in mass spectrometry analysis. Proteomics 2022, 22, e2100322. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Tan, Z.; Zhao, K.; Zou, W.; Wang, H.; Gao, H.; Sun, S.; Bu, D.; Chai, W.; Li, Y. The effect of N-glycosylation of SARS-CoV-2 spike protein on the virus interaction with the host cell ACE2 receptor. iScience 2021, 24, 103272. [Google Scholar] [CrossRef] [PubMed]
- Dawood, A.A. Glycosylation, ligand binding sites and antigenic variations between membrane glycoprotein of COVID-19 and related coronaviruses. Vacunas 2021, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shajahan, A.; Pepi, L.E.; Rouhani, D.S.; Heiss, C.; Azadi, P. Glycosylation of SARS-CoV-2: Structural and functional insights. Anal. Bioanal. Chem. 2021, 413, 7179–7193. [Google Scholar] [CrossRef]
- uniprot.org. P0DTC5: VME1_SARS2. Available online: https://services.healthtech.dtu.dk/services/NetNGlyc-1.0/ (accessed on 27 September 2023).
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. In Biocomputing; World Scientific: Singapore, 2002; pp. 310–322. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Cutoff | Sensitivity | Specificity | Mean Value | ||
|---|---|---|---|---|---|---|
| [%] | [%] | Positive | Negative | |||
| Wildtype | >0.067 | 96.30 | 100.00 | 0.659 ± 0.63 | 0.058 ± 0.003 | |
| Alpha | >0.067 | 91.36 | 100.00 | 0.653 ± 0.61 | 0.060 ± 0.003 | |
| Beta | >0.078 | 91.36 | 90.91 | 0.548 ± 0.60 | 0.069 ± 0.021 | |
| Gamma | >0.094 | 86.42 | 100.00 | 0.619 ± 0.67 | 0.060 ± 0.012 | |
| Kappa | >0.090 | 98.77 | 90.91 | 0.843 ± 0.78 | 0.088 ± 0.037 | |
| BA.1 | >0.182 | 92.59 | 81.82 | 0.434 ± 0.31 | 0.170 ± 0.042 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarze, M.; Volke, D.; Rojas Echeverri, J.C.; Schick, R.; Lakowa, N.; Grünewald, T.; Wolf, J.; Borte, S.; Scholz, M.; Krizsan, A.; et al. Influence of Mutations and N-Glycosylation Sites in the Receptor-Binding Domain (RBD) and the Membrane Protein of SARS-CoV-2 Variants of Concern on Antibody Binding in ELISA. Biology 2024, 13, 207. https://doi.org/10.3390/biology13040207
Schwarze M, Volke D, Rojas Echeverri JC, Schick R, Lakowa N, Grünewald T, Wolf J, Borte S, Scholz M, Krizsan A, et al. Influence of Mutations and N-Glycosylation Sites in the Receptor-Binding Domain (RBD) and the Membrane Protein of SARS-CoV-2 Variants of Concern on Antibody Binding in ELISA. Biology. 2024; 13(4):207. https://doi.org/10.3390/biology13040207
Chicago/Turabian StyleSchwarze, Mandy, Daniela Volke, Juan Camilo Rojas Echeverri, Robin Schick, Nicole Lakowa, Thomas Grünewald, Johannes Wolf, Stephan Borte, Markus Scholz, Andor Krizsan, and et al. 2024. "Influence of Mutations and N-Glycosylation Sites in the Receptor-Binding Domain (RBD) and the Membrane Protein of SARS-CoV-2 Variants of Concern on Antibody Binding in ELISA" Biology 13, no. 4: 207. https://doi.org/10.3390/biology13040207
APA StyleSchwarze, M., Volke, D., Rojas Echeverri, J. C., Schick, R., Lakowa, N., Grünewald, T., Wolf, J., Borte, S., Scholz, M., Krizsan, A., & Hoffmann, R. (2024). Influence of Mutations and N-Glycosylation Sites in the Receptor-Binding Domain (RBD) and the Membrane Protein of SARS-CoV-2 Variants of Concern on Antibody Binding in ELISA. Biology, 13(4), 207. https://doi.org/10.3390/biology13040207