

Extracellular Vesicles Contribute to Oxidized LDL-Induced Stromal Cell Proliferation in Benign Prostatic Hyperplasia

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and High-Fat Diet (HFD)
2.2. Primary Cultures of Prostatic Stromal Cells and Cell Lines
2.3. Proliferation Assays
2.4. Isolation of HPSCs-Derived EVs
2.5. Cell and EV Characterization by Transmission Electron Microscopy (TEM)
2.6. EV-Induced Cell Proliferation
2.7. Statistical Analysis
3. Results
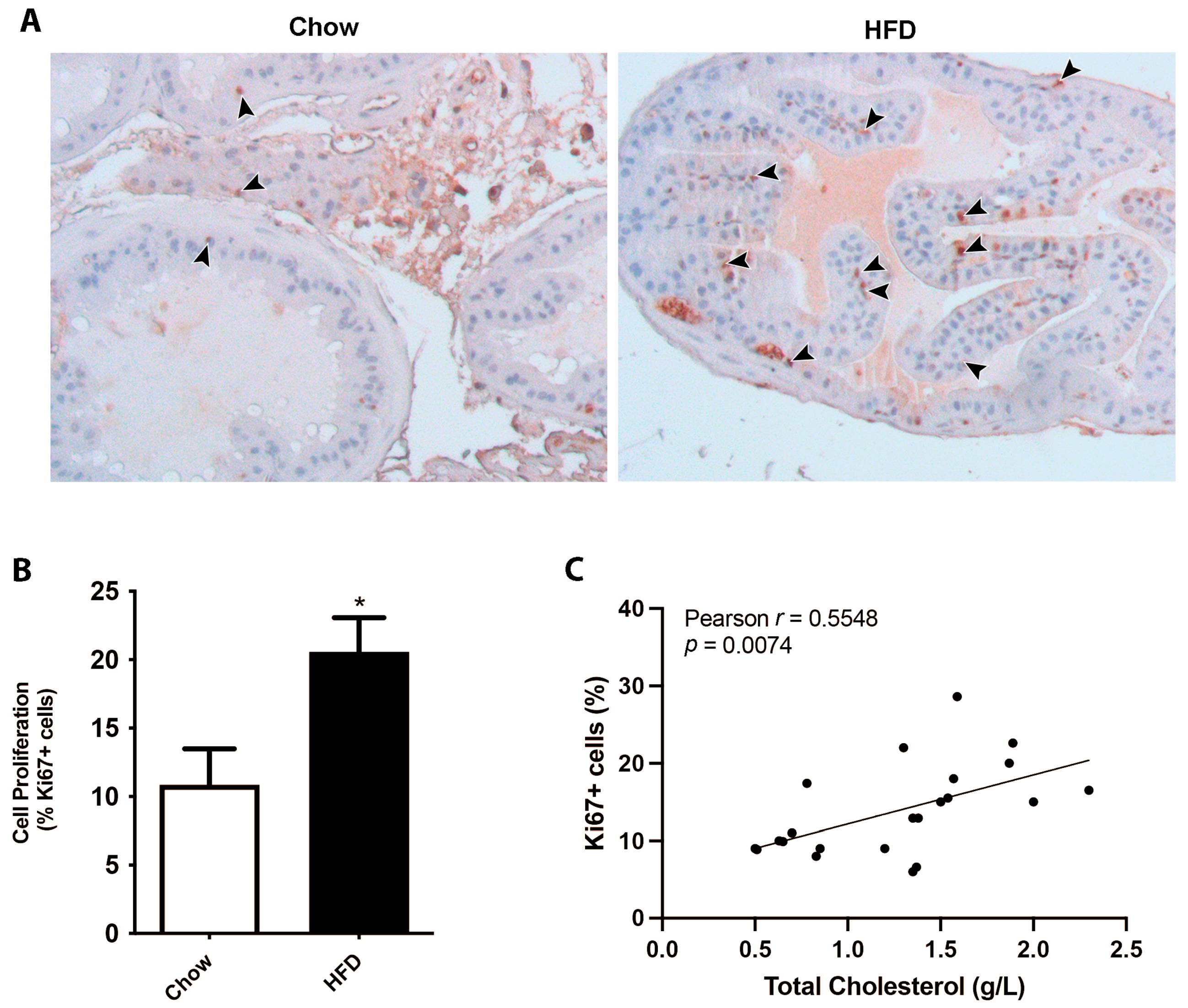
3.1. Mice on a HFD Displayed an Increase in Prostate Cell Proliferation
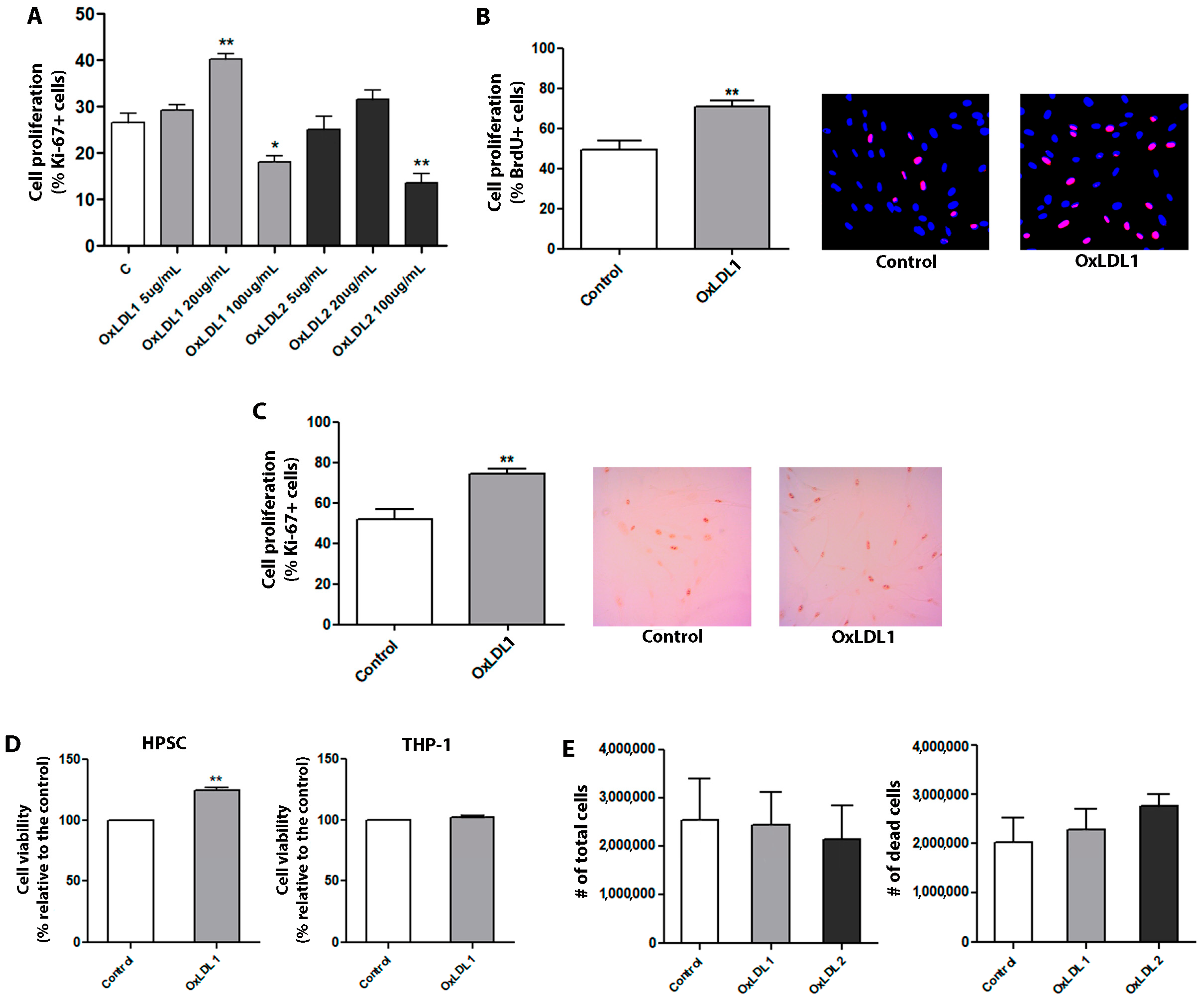
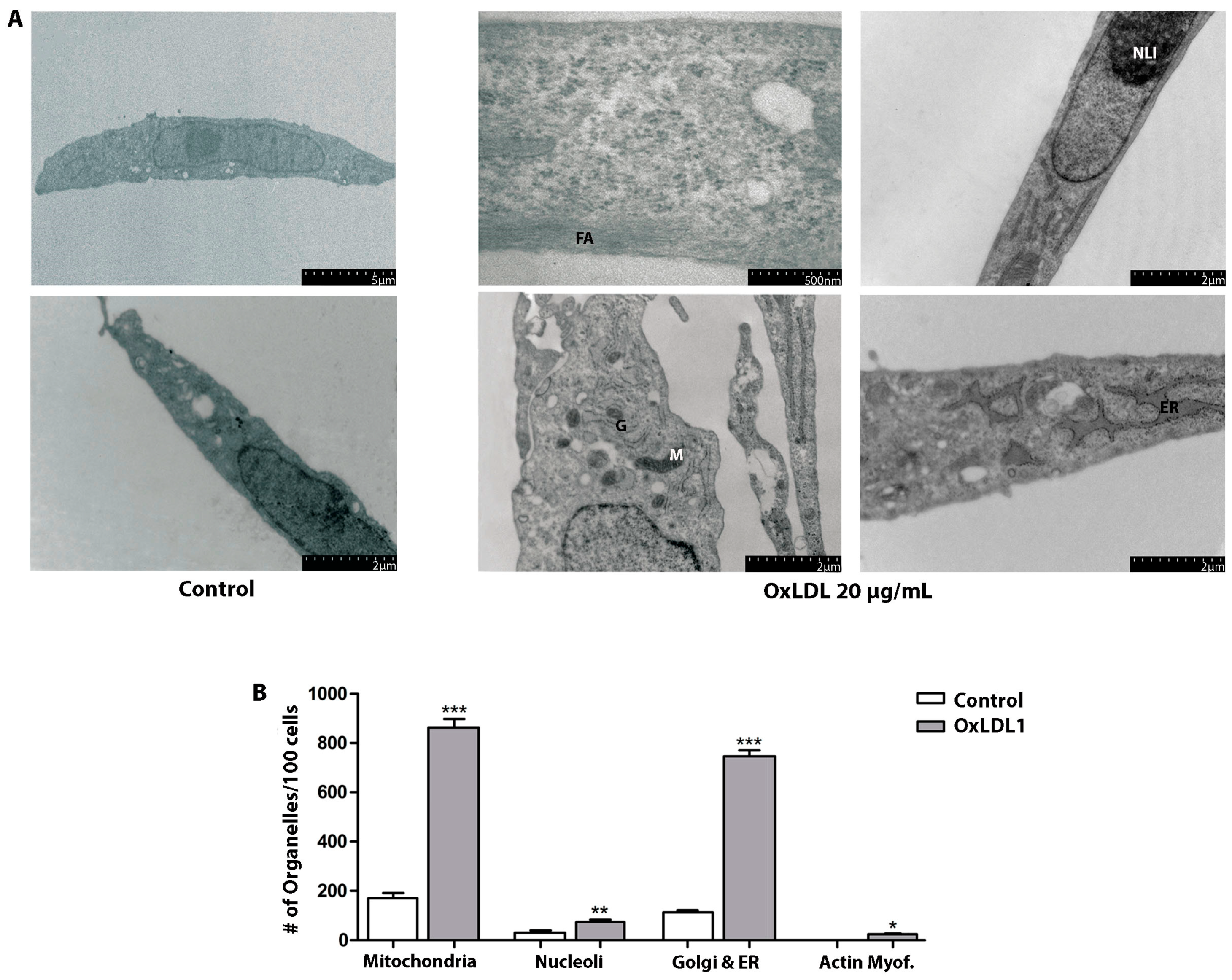
3.2. OxLDL Promoted Proliferation and Phenotypic Changes in Prostatic Stromal Cells Derived from Patients with BPH
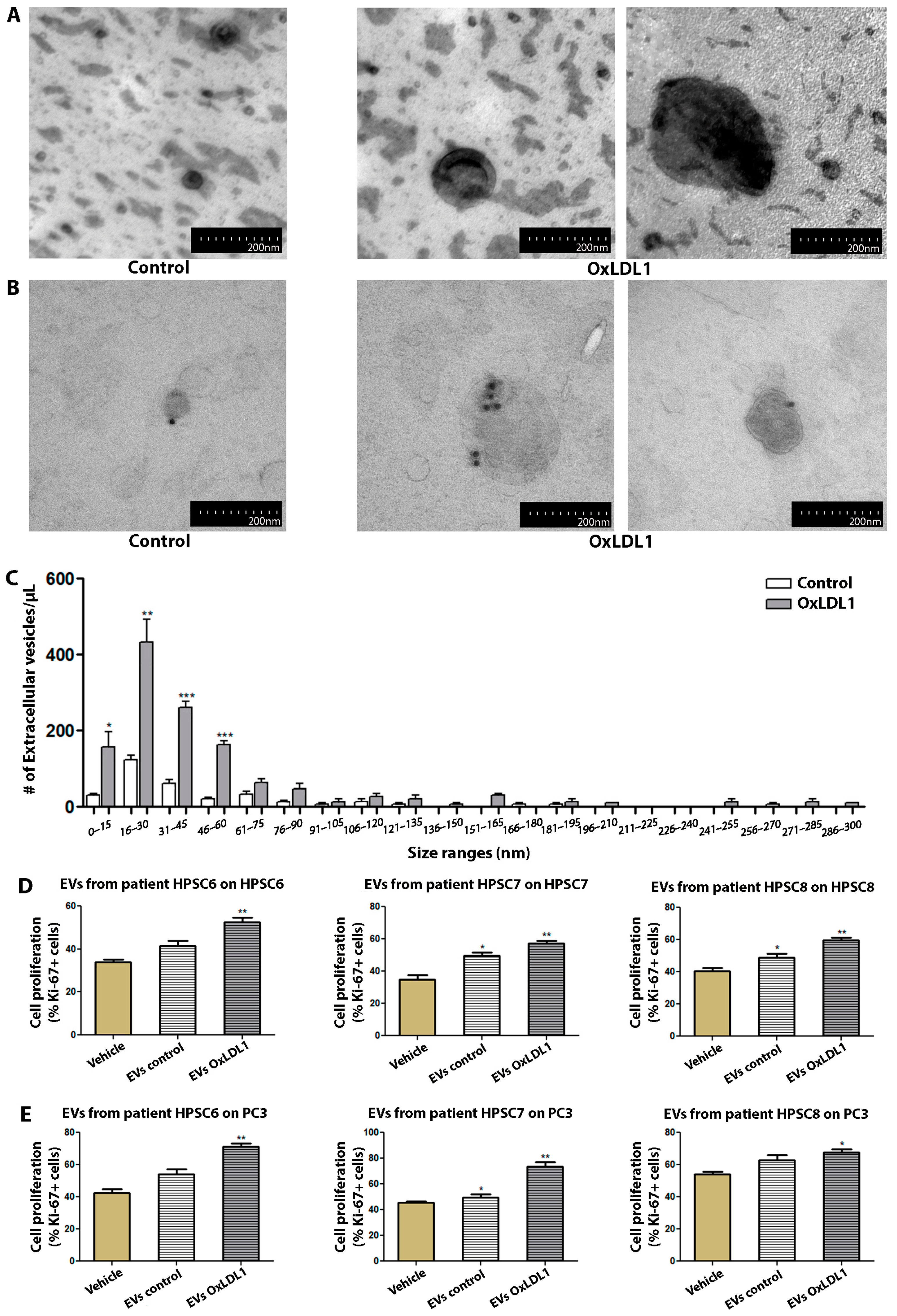
3.3. HPSCs Secreted EVs with Proliferative Activity in Response to OxLDL Stimulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannarella, R.; Condorelli, R.A.; Barbagallo, F.; La Vignera, S.; Calogero, A.E. Endocrinology of the Aging Prostate: Current Concepts. Front. Endocrinol. 2021, 12, 554078. [Google Scholar] [CrossRef] [PubMed]
- Vuichoud, C.; Loughlin, K.R. Benign prostatic hyperplasia: Epidemiology, economics and evaluation. Can. J. Urol. 2015, 22, 1–6. [Google Scholar] [PubMed]
- Smith, R.D.; Patel, A. Transurethral resection of the prostate revisited and updated. Curr. Opin. Urol. 2011, 21, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Fisac, J.L.; Royo-Bordonada, M.Á.; Rodríguez-Artalejo, F. Riesgos asociados a la dieta occidental y al sedentarismo: La epidemia de obesidad. Gac. Sanit. 2006, 20, 48–54. [Google Scholar] [CrossRef]
- Reiner, Ž. Hypertriglyceridaemia and risk of coronary artery disease. Nat. Rev. Cardiol. 2017, 14, 401–411. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, E139–E596. [Google Scholar] [CrossRef]
- Sebastianelli, A.; Gacci, M. Current Status of the Relationship Between Metabolic Syndrome and Lower Urinary Tract Symptoms. Eur. Urol. Focus. 2018, 4, 25–27. [Google Scholar] [CrossRef]
- Gacci, M.; Corona, G.; Vignozzi, L.; Salvi, M.; Serni, S.; De Nunzio, C.; Tubaro, A.; Oelke, M.; Carini, M.; Maggi, M. Metabolic syndrome and benign prostatic enlargement: A systematic review and meta-analysis. BJU Int. 2015, 115, 24–31. [Google Scholar] [CrossRef]
- Carvajal-Carvajal, C. LDL oxidada y la aterosclerosis. Med. Leg Costa Rica 2015, 32, 161–169. [Google Scholar]
- Yu, S.; Wong, S.L.; Lau, C.W.; Huang, Y.; Yu, C.M. Oxidized LDL at low concentration promotes in-vitro angiogenesis and activates nitric oxide synthase through PI3K/Akt/eNOS pathway in human coronary artery endothelial cells. Biochem. Biophys. Res. Commun. 2011, 407, 44–48. [Google Scholar] [CrossRef]
- Zhang, C.; Adamos, C.; Oh, M.J.; Baruah, J.; Ayee, M.A.; Mehta, D.; Wary, K.K.; Levitan, I. oxLDL induces endothelial cell proliferation via Rho/ROCK/Akt/p27kip1 signaling: Opposite effects of oxLDL and cholesterol loading. Am. J. Physiol.-Cell Physiol. 2017, 313, C340–C351. [Google Scholar] [CrossRef] [PubMed]
- Galle, J.; Hansen-Hagge, T.; Wanner, C.; Seibold, S. Impact of oxidized low density lipoprotein on vascular cells. Atherosclerosis 2006, 185, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Seasholtz, T.M.; Zhang, T.; Morissette, M.R.; Howes, A.L.; Yang, A.H.; Brown, J.H. Increased expression and activity of RhoA are associated with increased DNA synthesis and reduced p27(Kip1) expression in the vasculature of hypertensive rats. Circ. Res. 2001, 89, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Vignozzi, L.; Rastrelli, G.; Corona, G.; Gacci, M.; Forti, G.; Maggi, M. Benign prostatic hyperplasia: A new metabolic disease? J. Endocrinol. Investig. 2014, 37, 313–322. [Google Scholar] [CrossRef]
- Van Niel, G.; Carter, D.R.F.; Clayton, A.; Lambert, D.W.; Raposo, G.; Vader, P. Challenges and directions in studying cell-cell communication by extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2022, 23, 369–382. [Google Scholar] [CrossRef]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Falda, M.; Bussolati, B.; Tetta, C.; et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef]
- Hergenreider, E.; Heydt, S.; Tréguer, K.; Boettger, T.; Horrevoets, A.J.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.S.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat. Cell Biol. 2012, 14, 249–256. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, Y.; Cai, X.; Zhang, Y.; Yan, S.; Wang, J.; Zhang, S.; Yin, T.; Yang, C.; Yang, J. Extracellular vesicles derived from M1 macrophages deliver miR-146a-5p and miR-146b-5p to suppress trophoblast migration and invasion by targeting TRAF6 in recurrent spontaneous abortion. Theranostics 2021, 11, 5813–5830. [Google Scholar] [CrossRef]
- Dong, L.; Pu, Y.; Zhang, L.; Qi, Q.; Xu, L.; Li, W.; Wei, C.; Wang, X.; Zhou, S.; Zhu, J.; et al. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles promote lung adenocarcinoma growth by transferring miR-410. Cell Death Dis. 2018, 9, 218. [Google Scholar] [CrossRef]
- He, S.; Wu, C.; Xiao, J.; Li, D.; Sun, Z.; Li, M. Endothelial extracellular vesicles modulate the macrophage phenotype: Potential implications in atherosclerosis. Scand. J. Immunol. 2018, 87, 12648. [Google Scholar] [CrossRef]
- Nguyen, M.A.; Karunakaran, D.; Geoffrion, M.; Cheng, H.S.; Tandoc, K.; Perisic Matic, L.; Hedin, U.; Maegdefessel, L.; Fish, J.E.; Rayner, K.J. Extracellular Vesicles Secreted by Atherogenic Macrophages Transfer MicroRNA to Inhibit Cell Migration. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 49–63. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Z.H.; Kong, J.; Yang, M.Y.; Jiang, G.H.; Wang, X.P.; Zhong, M.; Zhang, Y.; Deng, J.T.; Zhang, W. Oxidized low-density lipoprotein-dependent platelet-derived microvesicles trigger procoagulant effects and amplify oxidative stress. Mol. Med. 2012, 18, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Roldán Gallardo, F.F.; Quintar, A.A. The pathological growth of the prostate gland in atherogenic contexts. Exp. Gerontol. 2021, 148, 111304. [Google Scholar] [CrossRef]
- Gómez-Elías, M.D.; Rainero Cáceres, T.S.; Giaccagli, M.M.; Guazzone, V.A.; Dalton, G.N.; De Siervi, A.; Cuasnicu, P.S.; Cohen, D.J.; Da Ros, V.G. Association between high-fat diet feeding and male fertility in high reproductive performance mice. Sci. Rep. 2019, 9, 18546. [Google Scholar] [CrossRef] [PubMed]
- Peinetti, N.; Rubio, M.M.C.; Sosa, L.D.V.; Scalerandi, M.V.; Alasino, R.V.; Peyret, V.; Nicola, J.P.; Beltramo, D.M.; Quintar, A.A.; Maldonado, C.A. Testosterone-loaded GM1 micelles targeted to the intracellular androgen receptor for the specific induction of genomic androgen signaling. Int. J. Pharm. 2020, 591, 119985. [Google Scholar] [CrossRef]
- Zago, V.; Sanguinetti, S.; Brites, F.; Berg, G.; Verona, J.; Basilio, F.; Wikinski, R.; Schreier, L. Impaired high density lipoprotein antioxidant activity in healthy postmenopausal women. Atherosclerosis 2004, 177, 203–210. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.; Erdbrügger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef] [PubMed]
- Peinetti, N.; Scalerandi, M.V.; Cuello Rubio, M.M.; Leimgruber, C.; Nicola, J.P.; Torres, A.I.; Quintar, A.A.; Maldonado, C.A. The Response of Prostate Smooth Muscle Cells to Testosterone Is Determined by the Subcellular Distribution of the Androgen Receptor. Endocrinology 2018, 159, 945–956. [Google Scholar] [CrossRef]
- Oregel-Cortez, M.I.; Frayde-Gómez, H.; Quintana-González, G.; García-González, V.; Vazquez-Jimenez, J.G.; Galindo-Hernández, O. Resistin Induces Migration and Invasion in PC3 Prostate Cancer Cells: Role of Extracellular Vesicles. Life 2023, 13, 2321. [Google Scholar] [CrossRef]
- Luo, F.; Guo, Y.; Ruan, G.Y.; Long, J.K.; Zheng, X.L.; Xia, Q.; Zhao, S.P.; Peng, D.Q.; Fang, Z.F.; Li, X.P. Combined use of metformin and atorvastatin attenuates atherosclerosis in rabbits fed a high-cholesterol diet. Sci. Rep. 2017, 7, 2169. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.S.; Huang, H.R.; Zhang, L.Y.; Kim, S.; He, Y.; Li, D.L.; Farischon, C.; Zhang, K.; Zheng, X.; Du, Z.Y.; et al. Mechanistic study of inhibitory effects of metformin and atorvastatin in combination on prostate cancer cells in vitro and in vivo. Biol. Pharm. Bull. 2017, 40, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- Pytlowanciv, E.Z.; Ribeiro, D.L.; Tamarindo, G.H.; Taboga, S.R.; Góes, R.M. High-fat diet during sexual maturation induces hyperplastic differentiation of rat prostate and higher expression of AR45 isoform and ERα. Reprod. Biol. 2022, 22, 100674. [Google Scholar] [CrossRef] [PubMed]
- Galle, J.; Heinloth, A.; Wanner, C.; Heermeier, K. Dual effect of oxidized LDL on cell cycle in human endothelial cells through oxidative stress. Kidney Int. 2001, 59, S120–S123. [Google Scholar] [CrossRef]
- Seibold, S.; Schürle, D.; Heinloth, A.; Wolf, G.; Wagner, M.; Galle, J. Oxidized LDL induces proliferation and hypertrophy in human umbilical vein endothelial cells via regulation of p27Kip1 expression: Role of RhoA. J. Am. Soc. Nephrol. 2004, 15, 3026–3034. [Google Scholar] [CrossRef]
- González-Chavarría, I.; Fernandez, E.; Gutierrez, N.; González-Horta, E.E.; Sandoval, F.; Cifuentes, P.; Castillo, C.; Cerro, R.; Sanchez, O.; Toledo, J.R. LOX-1 activation by oxLDL triggers an epithelial mesenchymal transition and promotes tumorigenic potential in prostate cancer cells. Cancer Lett. 2018, 414, 34–43. [Google Scholar] [CrossRef]
- Haga, N.; Akaihata, H.; Hata, J.; Hiraki, H.; Honda, R.; Tanji, R.; Onagi, A.; Koguchi, T.; Hoshi, S.; Ogawa, S.; et al. The association between local arteriosclerosis of the prostatic arteries and chronic inflammation in human benign prostatic enlargement. Prostate 2019, 79, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Van Haastert, P.J.M. Unified control of amoeboid pseudopod extension in multiple organisms by branched F-actin in the front and parallel F-actin/myosin in the cortex. PLoS ONE 2020, 15, 0243442. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Wang, W.; Chen, Y.; Liu, J.; Zhang, Z.; Wu, C.; Jiang, X.; Liang, Y.; Zhang, J. Bioelectric fields coordinate wound contraction and re-epithelialization process to accelerate wound healing via promoting myofibroblast transformation. Bioelectrochemistry 2022, 148, 108247. [Google Scholar] [CrossRef]
- Barron, D.A.; Rowley, D.R. The reactive stroma microenvironment and prostate cancer progression. Endocr. Relat. Cancer 2012, 19, 187–204. [Google Scholar] [CrossRef]
- Villa, M.; Cerda-Opazo, P.; Jimenez-Gallegos, D.; Garrido-Moreno, V.; Chiong, M.; Quest, A.F.; Toledo, J.; Garcia, L. Pro-fibrotic effect of oxidized LDL in cardiac myofibroblasts. Biochem. Biophys. Res. Commun. 2020, 524, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Vignozzi, L.; Gacci, M.; Cellai, I.; Santi, R.; Corona, G.; Morelli, A.; Rastrelli, G.; Comeglio, P.; Sebastanelli, A.; Maneschi, E.; et al. Fat boosts, while androgen receptor activation counteracts, BPH-associated prostate inflammation. Prostate 2013, 73, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Morale, M.G.; Tamura, R.E.; Rubio, I.G.S. Metformin and Cancer Hallmarks: Molecular Mechanisms in Thyroid, Prostate and Head and Neck Cancer Models. Biomolecules 2022, 12, 357. [Google Scholar] [CrossRef]
- Deza, Z.; Caimi, G.R.; Noelia, M.; Coli, L.; Ridruejo, E.; Alvarez, L. Atorvastatin shows antitumor effect in hepatocellular carcinoma development by inhibiting angiogenesis via TGF-β1/pERK signaling pathway. Mol. Carcinog. 2022, 62, 398–407. [Google Scholar] [CrossRef]
- Zhu, Z.; Cao, Y.; Liu, L.; Zhao, Z.; Yin, H.; Wang, H. Atorvastatin regulates the migration and invasion of prostate cancer through the epithelial-mesenchymal transformation and matrix metalloproteinase pathways. Investig. Clin. Urol. 2022, 63, 350. [Google Scholar] [CrossRef]
- Saraei, P.; Asadi, I.; Kakar, M.A.; Moradi-Kor, N. The beneficial effects of metformin on cancer prevention and therapy: A comprehensive review of recent advances. Cancer Manag. Res. 2019, 11, 3295. [Google Scholar] [CrossRef]
- Maaninka, K.; Neuvonen, M.; Kerkelä, E.; Hyvärinen, K.; Palviainen, M.; Kamali-Moghaddam, M.; Federico, A.; Greco, D.; Laitinen, S.; Öörni, K.; et al. OxLDL sensitizes platelets for increased formation of extracellular vesicles capable of finetuning macrophage gene expression. Eur. J. Cell Biol. 2023, 102, 151311. [Google Scholar] [CrossRef]
- Wang, X.; Xu, F.; Kou, H.; Zheng, Y.; Yang, J.; Xu, Z.; Fang, Y.; Sun, W.; Zhu, S.; Jiang, Q.; et al. Stromal cell-derived small extracellular vesicles enhance radioresistance of prostate cancer cells via interleukin-8-induced autophagy. J. Extracell. Vesicles 2023, 12, 12342. [Google Scholar] [CrossRef] [PubMed]
- Aalberts, M.; Stout, T.A.E.; Stoorvogel, W. Prostasomes: Extracellular vesicles from the prostate. Reproduction 2014, 147, R1–R14. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Q.; Hosen, M.R.; Zietzer, A.; Flender, A.; Levermann, P.; Schmitz, T.; Frühwald, D.; Goody, P.; Nickenig, G.; et al. Atherosclerotic Conditions Promote the Packaging of Functional MicroRNA-92a-3p Into Endothelial Microvesicles. Circ. Res. 2019, 124, 575–587. [Google Scholar] [CrossRef]
- Vlaeminck-Guillem, V. Extracellular vesicles in prostate cancer carcinogenesis, diagnosis, and management. Front. Oncol. 2018, 8, 222. [Google Scholar] [CrossRef]
- Shephard, A.P.; Giles, P.; Mbengue, M.; Alraies, A.; Spary, L.K.; Kynaston, H.; Gurney, M.J.; Falcón-Pérez, J.M.; Royo, F.; Tabi, Z.; et al. Stroma-derived extracellular vesicle mRNA signatures inform histological nature of prostate cancer. J. Extracell. Vesicles 2021, 10, e12150. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Yadav, S.S.; Kumar, S.; Mehta, N.; Talwar, G.; Qadri, J.; Sarwar, S. Evaluating the role of extracellular vesicles as a biomarker under transmission electron microscope in prostate cancer and benign prostate hyperplasia patients. Urología 2022, 89, 210–215. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roldán Gallardo, F.F.; Martínez Piñerez, D.E.; Reinarz Torrado, K.F.; Berg, G.A.; Herzfeld, J.D.; Da Ros, V.G.; López Seoane, M.; Maldonado, C.A.; Quintar, A.A. Extracellular Vesicles Contribute to Oxidized LDL-Induced Stromal Cell Proliferation in Benign Prostatic Hyperplasia. Biology 2024, 13, 827. https://doi.org/10.3390/biology13100827
Roldán Gallardo FF, Martínez Piñerez DE, Reinarz Torrado KF, Berg GA, Herzfeld JD, Da Ros VG, López Seoane M, Maldonado CA, Quintar AA. Extracellular Vesicles Contribute to Oxidized LDL-Induced Stromal Cell Proliferation in Benign Prostatic Hyperplasia. Biology. 2024; 13(10):827. https://doi.org/10.3390/biology13100827
Chicago/Turabian StyleRoldán Gallardo, Franco F., Daniel E. Martínez Piñerez, Kevin F. Reinarz Torrado, Gabriela A. Berg, Jael D. Herzfeld, Vanina G. Da Ros, Manuel López Seoane, Cristina A. Maldonado, and Amado A. Quintar. 2024. "Extracellular Vesicles Contribute to Oxidized LDL-Induced Stromal Cell Proliferation in Benign Prostatic Hyperplasia" Biology 13, no. 10: 827. https://doi.org/10.3390/biology13100827
APA StyleRoldán Gallardo, F. F., Martínez Piñerez, D. E., Reinarz Torrado, K. F., Berg, G. A., Herzfeld, J. D., Da Ros, V. G., López Seoane, M., Maldonado, C. A., & Quintar, A. A. (2024). Extracellular Vesicles Contribute to Oxidized LDL-Induced Stromal Cell Proliferation in Benign Prostatic Hyperplasia. Biology, 13(10), 827. https://doi.org/10.3390/biology13100827