Simple Summary
Humans are diploid organisms, carrying one set of chromosomes from our mother and one from our father. Plants can exist as diploid or polyploid, carrying multiple chromosome copies. Plant breeders can cross polyploid crops in unique and beneficial ways that are not possible in diploid crops. It will be necessary to produce food in a more sustainable manner, with less virgin land turned over and fewer resources consumed, therefore the study of hybrid polyploid crops is essential for the improvement of our food system.
Abstract
The improvement in quantitative traits (e.g., yield, size) in F1 offspring over parent lines is described as hybrid vigour, or heterosis. There exists a fascinating relationship between parental genetic distance and genome dosage (polyploidy), and heterosis effects. The contribution of nuclear heterozygosity to heterosis is not uniform across diploid and polyploid crops, even within same species, thus demonstrating that polyploid crops should be part of any discussion on the mechanisms of heterosis. This review examines the records of correlating heterosis with parental genetic distance and the influence of adding supplementary genomes in wide crosses. Increasing nuclear heterozygosity through parental genetic distance has been shown to be an imperfect predictor for heterosis in a variety of commercial crops such as maize, rice, and pepper. However, increasing the ploidy level raises the maximum number of alleles that can be harboured at any one locus, and studies on crops such as oilseed rape, potato, alfalfa, maize, and rice have demonstrated that heterosis may be maximised upon increasing multi-locus nuclear heterozygosity. The novel heterotic phenotypes observed above the diploid level will contribute to our understanding on the mechanisms of heterosis and aid plant breeders in achieving the righteous goal of producing more food with fewer inputs.
1. Introduction
In 1912, Edward M. East and Herbert K. Hayes of the United States published the first comprehensive review of hybrid breeding in crops of commercial interest, chiefly maize and tobacco [1]. The phenomenon of hybrid vigour, or heterosis, was of great interest to “add many millions of dollars annually to the nation’s resources”. It was well established by then that a so-called state of heterozygosis in a first-generation offspring (filial 1; F1) from inbred parents would regularly produce taller and/or higher yielding crops. East and Hayes interrogated the data as they knew this, to try and determine why certain “subdivisions” of species produced vigorous offspring upon cross-fertilisation, while others produced weaker crops or were completely unviable. Over 100 years later, there have been considerable developments on the genetic and epigenetic consequences, as well as the molecular mechanisms at play when plants are cross-fertilised.
This review brings together the published records of wide crosses at various ploidy levels where there were no fatal fertility problems. The review will only consider crosses through sexual hybridisation, not regenerated plants from artificial means such as somatic hybridisation or embryo sac chromosome doubling [2]. This review highlights how limits for heterosis through increasing multi-locus nuclear heterozygosity at the diploid level may be overcome through whole genome duplication events or the addition of supplementary genomes. This review begins with a framing of the heterosis phenomenon with a number of valuable references if the reader wishes to learn more, but does not set out to be an all-encompassing review of the potential mechanisms at play; for that the author suggests the excellent reviews by Schnable and Springer [3] and Labroo et al. [4]. Discussions regarding different techniques for estimating genetic distance and their usefulness in predicting heterosis have been covered by dos Santos Dias et al. [5]. Finally, the author wishes to complement previous work on this topic by Washburn and Birchler [6], which was the inspiration for this review.
2. Crop Improvement through Heterosis
An F1 hybrid offspring is the genetic composite of the biparental nuclear genomes inherited from both parents and the maternal cytoplasmic genome. An F1 hybrid displays heterosis when it displays enhanced characteristics over one, or both, parents [7]. From a commercial viewpoint, a more practical definition of heterosis is the superior performance of an F1 hybrid over the best parent [8]. Commercial F1 hybrid seed has contributed between 10–50% of yield increases since the mid-20th century for many crops, including bread wheat (Triticum aestivum), maize (Zea mays), sorghum (Sorghum bicolor), soybean (Glycine max), sugar beet (Beta vulgaris), and upland cotton (Gossypium hirsutum), to name just six [9,10,11,12,13,14,15]. While the significance of heterosis to commercial crops is widely documented, an all-inclusive mechanism explaining heterosis is lacking. See Box 1 for a description of the molecular models used to explain the heterosis phenomenon and valuable references where these models have been tested and discussed. Whatever the contributing molecular event(s), this will be followed by a functional mechanism that will give rise to heterosis. Proposed mechanisms include the following: optimisation of rate-limiting enzyme activity in maize [16]; increase in gibberellic acid in maize [17] and wheat [18]; epigenetic changes to circadian regulatory genes in tetraploid crosses between Arabidopsis thaliana and Arabidopsis arenosa [19]; specific leaf transcriptomics in maize and Arabidopsis thaliana [20]; improved energy-use efficiency for growth and protein metabolism [21]; changes in 24 nt small-interfering RNA (siRNAs) in maize [22]; repression of stress-responsive genes in Arabidopsis thaliana [23]; and an increase in potassium uptake and transport in tobacco (Nicotiana tabacum) [24]. From this list, it is clear that there exists specific heterotic traits that arise from specific genetic or epigenetic changes in specific F1 hybrid plants.
Box 1. Molecular models to explain heterosis.
Contribution of dominance to heterosis
The dominance theory of heterosis may be most
easily comprehended as the opposite of inbreeding depression. It suggests
that hybridising genetically distinct genomes will lead to heterosis.
Assuming both parents harbour different recessive, slightly deleterious
alleles in a homozygous state, these will be complemented by a dominant,
superior copy from the other parent in the F1 hybrid. Thus, the
deleterious effect of these recessive alleles will be masked in the
heterozygous state. The practical challenge of achieving a homozygous (or
near-homozygous) condition in the parent lines in order to demonstrate a
heterosis effect on crop yield was clearly demonstrated in the pioneering
work of George H. Shull presented to the American Breeder’s Association [25,26].
Contribution of over-dominance to heterosis
It was addressed in the early literature by
Shull [27], Jones [28], and East [29] that a simple dominance mode of action does
not account for all the heterosis effects observed in many F1
plants. The over-dominance theory proposes that heterozygosity at a specific
locus or loci will lead to heterosis, while either homozygous condition will
not. This was skilfully demonstrated in tomato (Solanum lycopersicum)
by Krieger et al. [30], where considerable yield heterosis was
demonstrated when there was heterozygosity for loss-of-function alleles of SINGLE
FLOWER TRUSS (SFT). F1 hybrids homozygous for the sft
mutation do not display heterosis. Linked genes can make the task of
determining dominant from over-dominant mechanisms difficult. For example, it
was reported that over-dominant effects at several quantitative trait loci
(QTL) were responsible for maize heterosis [31]. These may be dominant effects. The QTL with
the largest influence identified were later found to be an effect called
“pseudo-overdominance”: the QTL were further dissected into (at least) two
distinct genes that were in a state of repulsion phase linkage. These linked
loci both contribute to heterosis via dominant mechanisms [3,32].
Contribution of epistasis to heterosis
Further nuance is required when considering
the epistasis theory of heterosis. This theory is defined as novel
interactions between alleles at two or more different loci leading to a
heterotic effect. For example, an over-dominant acting locus may interact
with a dominant-acting locus elsewhere in the genome of an F1
hybrid, thus contributing to heterosis [33]. This theory was first described in detail by
Powers [34]. Epistasis mechanisms have been proposed to account for
the majority of the heterotic effects in certain crosses of wild tomato Lycopersicon
hirsutum through the use of near-isogenic lines [35] and in
rice through chromosome segment substitution lines [36]. Both of
these approaches are examples of marker assisted selection, which is
admirable due to the large number of controlled crosses required to isolate
specific genomic regions and their associated allelic frequency.
Contribution of epigenetics to heterosis
While genetically disparate individuals may
induce heterosis through genetic mechanisms, it is probable that increased genetic differences are also associated with increased epigenetic differences [37]. In the context of heterosis, epigenetic effects can be
defined as heritable changes in gene activity which are unrelated to
underlying changes in DNA sequences [38]. To determine if parental epigenetic effects
alone can cause heterosis, it is necessary to maintain a population of inbred
lines that vary only for a segregating epigenetic marker. Such epigenetic
recombinant inbred lines (epiRILs) in A. thaliana have been crossed
with the relevant wild-type accession to demonstrate how changes in DNA
methylation, independent of genetic differences, can positively influence
plant height [39] and leaf area [40,41]. Attempts to create epiRILs in agronomically
important crops like maize [42] and rice [43,44] have been met with obstacles such as strong
lethality following alterations to the DNA methylation network.
3. Polyploid Heterosis
Increasing the number of chromosome sets in the nucleus, including whether the extra genome(s) is inherited maternally or paternally, can provide a neat experimental design to further test the limits of heterosis. In Arabidopsis thaliana, for example, there exists a parent-of-origin genome dosage effect on F1 triploid heterosis. If an F1 triploid inherits two paternal genomes, the heterosis effect on seed size and leaf area exceeds that seen in F1 diploid hybrids. However, this heterosis effect is ablated or indeed reversed if the F1 triploid hybrid inherits two maternal genomes [45,46,47]. It is hypothesised that the ratio of maternal:paternal genomes in the endosperm has a pronounced impact on inter-ploidy F1 seed size, with larger seeds giving rise to larger plants, although growth rates remain the same, demonstrating a “hidden” heterosis trait at the diploid level. The functional mechanism behind this effect on seed size remains to be fully explained [47]. A similar genome dosage effect on F1 triploid heterosis has been demonstrated in maize, although some heterotic traits are genotype dependent [48].
At the tetraploid level, it has been demonstrated in several plants (discussed below) that increasing heterozygosity in a double-cross (beyond a single cross) can increase the heterosis effect. This phenomenon has been termed ‘progressive heterosis’ and is accepted to be absent in diploids. This is because a tetraploid double-cross F1 hybrid can harbour up to four different alleles at any one locus (Figure 1). This suggests that increasing multi-locus nuclear heterozygosity by adding extra genomes could maximise heterosis.
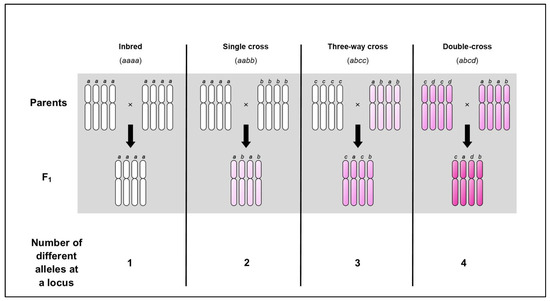
Figure 1.
F1 autotetraploid from a double-cross can harbour four different alleles at a locus. An autotetraploid contains four identical chromosome sets within its nucleus. Let us propose a hypothetical gene at a certain locus that has four different alleles, denoted a, b, c, and d. Inbred (100% homozygous) lines will possess a mono-allelic locus for this gene, i.e., a simplex genotype meaning four identical copies of one allele (aaaa and bbbb). Following cross-fertilisation, the single cross F1 hybrid will possess a di-allelic locus for this gene, i.e., a duplex genotype meaning two copies of two different alleles (aabb). This single cross F1 hybrid can be crossed to a different inbred line (cccc) and the resulting three-way cross F1 hybrid will possess a tri-allelic locus for this gene, i.e., a trigenic genotype meaning one copy of two different alleles and two identical copies of one allele (abcc). Alternatively, the single cross F1 hybrid can be crossed to a different single cross F1 hybrid (ccdd) and the resulting double-cross F1 hybrid will possess a tetra-allelic locus, i.e., a tetragenic genotype meaning one copy of four different alleles (abcd). The increasing intensity of the colour pink reflects the increasing levels of heterozygosity.
4. Increasing Multi-Locus Nuclear Heterozygosity and Heterosis
The F1 progeny of genetically disparate parents will harbour considerable nuclear heterozygosity. According to the dominance theory of heterosis, any increase beyond the homozygous state at a locus will maximise the heterosis effect. Thus, heterosis should correlate with nuclear heterozygosity. This experiment has been run a number of times with a diverse range of plant species—those both autogamous and allogamous in nature, with different growing habits, and at a range of ploidy levels.
For diploid plants of the same species (“autodiploid”), crossing genetically diverse parent lines in a single cross to significantly increase nuclear heterozygosity is an imprecise method of maximising heterosis. Crossing genetically diverse parent lines in maize appears to maximise heterosis, although a certain threshold is reached, according to Moll [49]. Early work with restriction fragment link polymorphisms (RFLPs) also indicated a significant [50] and intermediate [51] relationship between genetic distance and heterosis. However, subsequent work with a variety of molecular markers has reached the opposite conclusion [52,53,54,55,56]. Perhaps the most comprehensive work was by Reif et al. [55], where maize F1 hybrids of European and United States inbred lines were examined at multi-location field trials. Likewise, for pepper (Capsicum annuum) [57], rice (Oryza sativa) [58], soybean (Glycine max) [59], sesame (Sesamum indicum) [60], white clover (Trifolium repens) [61], chickpea (Cicer arietinum) [62], eggplant (Solanum melongena) [63], pearl millet (Pennisetum glaucum) [64], broad bean (Vicia faba) [65]), oil palm (Elais guineensis) [66], pigeon pea (Cajanus cajan) [67], melon (Cucumis melo) [68], the amphidiploid Ethiopian mustard (Brassica carinata) [69], and the model organism Arabidopsis thaliana [70,71], heterosis at the diploid level is not maximised through crossing genetically diverse parent lines. Outliers to this broad conclusion are found in diploid cocoa (Theobroma cacao) [72,73] and diploid Robusta coffee (Coffea canephora) [74], as well as limited evidence in sorghum (Sorghum bicolor) [75]. Likewise, in diploid sunflower (Helianthus annus) there were early reports that heterosis correlates with genetic distance, although the authors found differentiating highly heterozygous open-pollinated varieties difficult [76]. Subsequent work with elite Russian varieties also found a significant correlation [77]. Interesting results have been reported in diploid carrot (Daucus carota) where RFLP markers suggest a relationship between genetic distance and total yield in F1 hybrids, but amplified fragment length polymorphism (AFLP) markers—which are generally accepted to be more sensitive [78]—do not [79]. Interspecific hybridisation (mating across taxonomically-defined species boundaries) may produce viable plants. At the diploid level (“allodiploid”) there are some prominent examples of heterosis when crossing members of the radish (Raphanus) and Brassica genus [80,81]. However, pre- and post-fertilisation barriers commonly prevent successful allodiploid formation.
There is a strong record of maximising heterosis through crossing genetically diverse parent lines in polyploid plants (Table 1). The F1 hybrids in Table 1 represent controlled crosses among well-defined genotypes, i.e., inbred lines, near-inbred lines, or elite commercial varieties, not distant crosses with wild relatives. Progressive heterosis has been recorded in many crops which are commercially grown at the tetraploid level, such as potato [82] and alfalfa [83,84]. Of note, the effect has also been recorded in maize [53,85] and rice [86,87], which gives further credence to the view that heterosis at the diploid level does not predict heterosis at the polyploid level. It is important to highlight when authors investigate the same plant species but use different methodologies to determine genetic differences. In oilseed rape (Brassica napus), contrasting conclusions have been drawn by Diers et al. [88] and Riaz et al. [89], with the former using RFLPs and the latter using sequence-related amplified polymorphism (SRAP) markers. Diers and colleagues conclude that there is no meaningful relationship between genetic distance and heterosis, whereas Riaz and colleagues found a significant relationship between the two. An earlier study using knowledge of the geographical origin of oilseed rape varieties also found a significant relationship [90]. For bread wheat using either PCR sequence tagged sites (STSs) or RFLPs, both demonstrate an absence of any relationship between genetic distance and heterosis (Table 1).
Why do many polyploid crops tend to demonstrate heterosis upon large increases in nuclear heterozygosity, but diploid crops do not? Taking the examples in Table 1, it appears that for many crops, increasing the number of different alleles has a positive dosage effect on vigour. Intralocus (i.e., epistatic) interactions have been attributed to the heterosis effects in polyploid potato [91], although it has been demonstrated that a three-way cross may be no more beneficial than a two-way cross (Figure 1) [92], but this finding appears to be in the minority (Table 1). This suggests that favourable alleles are scattered across the genome and their positive impact are fully actualised in a polyploid plant. However, concluding that multiple alleles at a certain locus are the chief driver of heterotic effects may overlook the presence of a tight collection of linked genes along a small chromosomal segment—a ‘linkat’—which may be indistinguishable from multiple alleles [93]. In alfalfa, it has been suggested that the number of different alleles at a locus is only one part of the explanation for progressive heterosis, and the effect of complementary action between genes could be more influential [94]. For example, suppose heterosis arising from a certain gene action is present in equivalent diploid and autotetraploid plants, but the differences between disomic and tetrasomic segregation will produce a higher frequency of heterozygotes in the autotetraploid; thus, an F1 arising from tetrasomic segregation is more likely to possess loci containing at a least one dominant allele, potentially masking inferior, recessive alleles (i.e., over-dominance) [95] (see schematic below). In addition, the influence of complementary gene action on heterosis in alfalfa is clearly outlined by Bingham et al. [94] using the example of “two-allele populations”. Taking equivalent diploid and autotetraploid populations which possess a maximum of two different alleles at a locus (e.g., genotype Aa and genotype AAaa), populations are maintained for a number of generations (e.g., self and sib-mating). Bingham and colleagues discuss the expression of heterosis vs. inbreeding depression in these populations and explain that the presence of different alleles in the population (A, A1, A2, A3, …Aj) existing at ‘linkats’, which display additive effects on the phenotype, are likely responsible (an additive effect on the phenotype would manifest as a linear increase, whereas a non-additive effect on the phenotype would be non-linear). The tetrasomic segregation of favourable alleles at ‘linkats’ in autotetraploids present greater opportunities for complementary gene actions than seen in diploids. The converse of this, upon repeated selfing, autotetraploid alfalfa display inbreeding depression at much higher levels than would be expected considering they take a longer time to reach homozygosity than an equivalent diploid [96]. The difference in levels of inbreeding depression between identical genotypes cannot be attributed to recessive alleles alone, but rather the loss of complementary gene action in the autotetraploid as allelic dosage changes faster than homozygosity [94]. Remarkably, a swift display of inbreeding depression has also been reported in autotetraploids of maize [97] and sugar beet [98], while autotetraploids of rye (Secale cereale) show a similar level of inbreeding depression to their equivalent diploids [99,100]. This suggests the breakdown of complementary gene action upon inbreeding is not a rare occurrence in autotetraploids.
If autotetraploidy is a reliable route to maximising heterosis, what are the barriers to widespread adoption of double-cross autotetraploid hybrids? The answer can be species or genotype specific. While potato, alfalfa, and perennial ryegrass are commercially grown at the tetraploid level, maize is commercially grown at the diploid level. In maize, increasing the ploidy level can have an immediate, negative impact on plant growth in some genotypes [101]. Tetraploid maize has lower fertility than equivalent diploids [102] and they can potentially exhibit ‘double reduction’ in meiosis, where incorrect separation of sister chromatids during the first meiotic division means the resulting gamete contains both sister chromatids [103]. Thus, the progeny from a double-cross tetraploid maize could possess different genotypes. Doubling the genome dosage can likewise have a detrimental effect on various morphological characteristics in some plants [104]. Autotetraploid rice has a less than ideal fertility level for commercial purposes [105]. Upon increasing the genome dosage, a tetraploid plant typically displays a slower growth rate [106]. This change would need to be incorporated into agronomic practices, e.g., growing degree days and chemical applications. Lastly, selecting inbred lines in a heterozygous autotetraploid population can be challenging in certain circumstances [107]. If the objective is to remove a deleterious allele at a certain locus (i.e., achieve complete dominance), tetrasomic segregation in an autotetraploid makes selection more cumbersome than disomic segregation in an equivalent diploid. For example, assume the hypothetical allele A shows incomplete dominance over allele a. Upon selfing, segregation at a heterozygous locus Aa in a diploid will segregate so that 1/4 of the progeny will show complete dominance.
| ♀ | A | a | |
| ♂ | |||
| A | AA | Aa | |
| a | Aa | aa | |
The same heterozygous locus AAaa in an autotetraploid will produce far more undesirable genotypes harbouring the recessive allele upon selfing (assuming full tetrasomic segregation), and only 1/36 of the progeny will show complete dominance.
| ♀ | AA | Aa | Aa | Aa | Aa | aa | |
| ♂ | |||||||
| AA | AAAA | AAAa | AAAa | AAAa | AAAa | AAaa | |
| Aa | AAAa | AAaa | AAaa | AAaa | AAaa | Aaaa | |
| Aa | AAAa | AAaa | AAaa | AAaa | AAaa | Aaaa | |
| Aa | AAAa | AAaa | AAaa | AAaa | AAaa | Aaaa | |
| Aa | AAAa | AAaa | AAaa | AAaa | AAaa | Aaaa | |
| aa | AAaa | Aaaa | Aaaa | Aaaa | Aaaa | aaaa | |
Wide crosses within cultivated wheat genotypes do not reliably induce heterosis (Table 1). Raising the “yield plateau” in wheat [108,109] to meet future demand is limited by the narrow genetic base among cultivated genotypes from a loss in variation during domestication and selection [110,111,112,113]. This suggests wheat breeders may need to focus on organising present germplasm into heterotic groups [114] or look to wild relatives to introduce novel alleles [115,116,117,118]. However, a commendable research effort producing over 1500 wheat F1 hybrids from European genotypes demonstrated the potential for an approximate 10% increase in yield above average, and the majority of heterotic effects were attributed to epistatic interactions [119]. This shows the complex heritability of yield in wheat, likely controlled by multiple loci, each with small effects. A hybrid between wheat and rye, Triticale, is common across Europe as a feed, energy, and food crop [120]. Also present at the allohexaploid level, commercial Triticale genotypes can exhibit heterosis in the F1 following crosses with genetically diverse parents [121]. The link was small but significant, suggesting that seeking out genetic diversity among cultivated Triticale could be an essential prerequisite when breeding for improved varieties. Likewise, cultivated forage grass Timothy (Phleum pratense) is an allohexaploid crop, and starting with parents with commercially favourable yield it is possible to improve yield further upon wide crosses [122]. This suggests that despite the presence of homeologous genomes and after a certain amount of selection, these crops continue to harbour potentially recessive allelic combinations.

Table 1.
An overview of the relationship between nuclear heterozygosity and heterosis in polyploid crops.
Table 1.
An overview of the relationship between nuclear heterozygosity and heterosis in polyploid crops.
| Ploidy Status | Crop | Association Observed between Crossing Genetically Diverse Parent Lines in a Single Cross and Heterosis in F1? | Reference |
|---|---|---|---|
| Autotriploid | Sugar beet (Beta vulgaris subsp. vulgaris) | No | [123] |
| Autotetraploid | Rye (Secale cereale) | Yes | [99] |
| Potato (Solanum tuberosum) | Yes | [82,92,124] | |
| No | [125] | ||
| Alfalfa (Medicago sativa) | Yes | [83,84] | |
| Maize (Zea mays) | Yes | [53,85] | |
| Perennial ryegrass (Lolium perenne) | Yes | [126] | |
| Rice (Oryza sativa) | Yes | [86,87] | |
| Bahiagrass (Paspalum notatum) | Yes | [127] | |
| Allotriploid | Willow ((Salix koriyanagi × S. purpurea) × S. miyabeana | Yes | [128] |
| Allotetraploid | Peanut (Arachis hypogaea) | No | [129] |
| Oilseed rape (Brassica napus) | Yes | [89,90] | |
| No | [88] | ||
| Arabica coffee (Coffea arabica) | Yes | [130] | |
| Upland cotton (Gossypium hirsutum) | No | [131] | |
| Allohexaploid | Bread wheat (Triticum aestivum subsp. aestivum) | No | [132,133] |
| Triticale | Yes | [121] | |
| Timothy (Phleum pratense) | Yes | [122] |
5. Commercial Breeding
For a hybrid breeding program to be commercially successful, the F1 must possess economically valuable traits that cover the cost of the program in question. The attributes of the crop (e.g., outcrossing or self-fertilising, availability of cytoplasmic male sterile lines, annual or perennial) will determine the most appropriate breeding plan to adopt. Regardless, F1 hybrid breeding programs can be made more efficient with some form of predictive value. Determining the genetic relatedness between parent lines can help develop clearly defined populations or heterotic groups, from which superior F1 hybrids can been formed through knowledge of combining ability.
To support commercial breeding programs in this endeavour, there exists a number of molecular biology tools. For example, gene expression analysis of messenger RNA (mRNA) can reveal transcriptional variation between parents which may be exploited for a heterotic effect in the F1 [134,135]. Regardless of ploidy level, this approach is difficult as it requires a specific developmental stage to be sampled, on precise tissue, across similar environmental conditions. Alternatively, genomic DNA can be sampled to create molecular markers (e.g., RFLPs, SSRs) across a set of parent lines and offspring which can be correlated with heterosis. High-throughput sequencing technologies can improve the efficiency of this process, but random markers may demonstrate poor linkage to QTL controlling traits of interest as more genetic backgrounds are introduced [136]. An improved use of molecular markers includes genomic selection, where a computer model is trained on high-density markers throughout the entire genome in combination with phenotype scores [137]. As new genotypes are added to the model, accurate phenotype prediction can improve commercial breeding resources [138,139]. In addition, fluorescent in situ hybridisation (FISH) can determine physical changes in chromosomes and identify copy number differences between parent lines and progeny. Such labelled DNA is useful to help visualise the presence or absence of chromosomes or chromosomal fragments. However, this technique is open to added complexity in polyploid plants due to genes with multiple copies throughout the genome from chromosomal duplications [140]. Combining genetic and metabolite markers to predict heterosis (e.g., single nucleotide polymorphisms (SNPs) with sugar, amino acid, organic acid profiles) can create “biomarkers” [141,142,143], although knowledge of the essential metabolites in the crop of interest and when to sample for the varying analyses is challenging.
A simple predictor of heterosis could be genetic distance between parents, as wide crosses could exploit novel gene recombination. Following such a cross, nuclear heterozygosity is substantially increased. However, diploid crops cannot exploit this level of heterozygosity whereas many polyploid crops can (Table 1). For the majority of diploid crops, wide crosses potentially introduce multiple loci with negative or neutral effects on heterosis. The documented exceptions to this observation are all out-crossing diploids (cocoa, coffee, sunflower), suggesting a preference to maintain a highly heterozygous genotype for plant development. Thus, together with all the molecular biology tools available to streamline the development of valuable F1 hybrids, polyploid breeding programs have potentially more favourable crosses at their disposal than diploid breeding programs.
6. Conclusions
To explain the presence or absence of heterosis, a number of molecular theories have been tested (Box 1). We now know that studying diploid crops in isolation means a number of hidden heterotic effects are absent from the discussion [6]. East and Hayes [1] noted that the heterozygous condition was generally favourable, although some F1 hybrids from genetically dissimilar parents showed little or no increase in vigour. If East and Hayes could have converted these diploid maize lines to the tetraploid state, heterosis may have been observed in the F1 [53,85]. This is because polyploid crops, even commercial lines, may be improved further through increasing multi-locus nuclear heterozygosity. Over 100 years following East and Hayes’ influential publication, adding a genome dosage variable to the heterosis equation typifies this essential, albeit enigmatic, tool for plant breeders.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analysed in this Review. Data sharing is not applicable.
Conflicts of Interest
The author previously worked for a company (Dairygold Agri Business Limited) that was a commercial distributor of hybrid seed.
References
- East, E.M.; Hayes, H.K. Heterozygosis in Evolution and in Plant Breeding; U.S. Government Printing Office: Washington, WA, USA, 1912.
- Cheng, S.; Zhu, X.; Liao, T.; Li, Y.; Yao, P.; Suo, Y.; Zhang, P.; Wang, J.; Kang, X. Gene Expression Differences between High-Growth Populus Allotriploids and Their Diploid Parents. Forests 2015, 6, 839–857. [Google Scholar] [CrossRef]
- Schnable, P.S.; Springer, N.M. Progress toward Understanding Heterosis in Crop Plants. Annu. Rev. Plant Biol. 2013, 64, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Labroo, M.R.; Studer, A.J.; Rutkoski, J.E. Heterosis and Hybrid Crop Breeding: A Multidisciplinary Review. Front. Genet. 2021, 12, 643761. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.A.D.S.; Picoli, E.A.D.T.; Rocha, R.B.; Alfenas, A.C. A priori choice of hybrid parents in plants. Genet. Mol. Res. 2004, 3, 356–368. [Google Scholar] [PubMed]
- Washburn, J.D.; Birchler, J.A. Polyploids as a ‘model system’ for the study of heterosis. Plant Reprod. 2014, 27, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Shull, G.H. What Is ‘Heterosis’? Genetics 1948, 33, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Acquaah, G. Principles of Plant Genetics and Breeding; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Miller, F.R.; Kebede, Y. Genetic Contributions to Yield Gains in Sorghum, 1950 to 1980. In Genetic Contributions to Yield Gains of Five Major Crop Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1984; pp. 1–14. [Google Scholar]
- Russell, W.A. Genetic Improvement of Maize Yields. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 1991; Volume 46, pp. 245–298. [Google Scholar]
- Brancourt-Hulmel, M.; Doussinault, G.; Lecomte, C.; Bérard, P.; Buanec, B.L.; Trottet, M. Genetic Improvement of Agronomic Traits of Winter Wheat Cultivars Released in France from 1946 to 1992. Crop Sci. 2003, 43, 37–45. [Google Scholar] [CrossRef]
- Duvick, D.N. The Contribution of Breeding to Yield Advances in maize (Zea mays L.). In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2005; Volume 86, pp. 83–145. [Google Scholar]
- Egli, D.B. Comparison of Corn and Soybean Yields in the United States: Historical Trends and Future Prospects. Agron. J. 2008, 100, S-79–S-88. [Google Scholar] [CrossRef]
- Campbell, B.T.; Chee, P.W.; Lubbers, E.; Bowman, D.T.; Meredith, W.R.; Johnson, J.; Fraser, D.; Bridges, W.; Jones, D.C. Dissecting Genotype × Environment Interactions and Trait Correlations Present in the Pee Dee Cotton Germplasm Collection following Seventy Years of Plant Breeding. Crop Sci. 2012, 52, 690–699. [Google Scholar] [CrossRef]
- Loel, J.; Kenter, C.; Märländer, B.; Hoffmann, C.M. Assessment of breeding progress in sugar beet by testing old and new varieties under greenhouse and field conditions. Eur. J. Agron. 2014, 52, 146–156. [Google Scholar] [CrossRef]
- Hageman, R.H.; Leng, E.R.; Dudley, J.W. A Biochemical Approach to Corn Breeding. In Advances in Agronomy; Norman, A.G., Ed.; Academic Press: Cambridge, MA, USA, 1967; Volume 19, pp. 45–86. [Google Scholar]
- Rood, S.B.; Buzzell, R.I.; Mander, L.N.; Pearce, D.; Pharis, R.P. Gibberellins: A Phytohormonal Basis for Heterosis in Maize. Science 1988, 241, 1216–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ni, Z.; Yao, Y.; Nie, X.; Sun, Q. Gibberellins and heterosis of plant height in wheat (Triticum aestivum L.). BMC Genet. 2007, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Kim, E.-D.; Ha, M.; Lackey, E.; Liu, J.; Zhang, Y.; Sun, Q.; Chen, Z.J. Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nature 2009, 457, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Stokes, D.; Fraser, F.; Morgan, C.; O’neill, C.M.; Dreos, R.; Magusin, A.; Szalma, S.; Bancroft, I. An association transcriptomics approach to the prediction of hybrid performance. Mol. Breed. 2010, 26, 91–106. [Google Scholar] [CrossRef]
- Goff, S.A. A unifying theory for general multigenic heterosis: Energy efficiency, protein metabolism, and implications for molecular breeding. New Phytol. 2011, 189, 923–937. [Google Scholar] [CrossRef]
- Barber, W.T.; Zhang, W.; Win, H.; Varala, K.K.; Dorweiler, J.E.; Hudson, M.E.; Moose, S.P. Repeat associated small RNAs vary among parents and following hybridization in maize. Proc. Natl. Acad. Sci. USA 2012, 109, 10444–10449. [Google Scholar] [CrossRef]
- Miller, M.; Song, Q.; Shi, X.; Juenger, T.E.; Chen, Z.J. Natural variation in timing of stress-responsive gene expression predicts heterosis in intraspecific hybrids of Arabidopsis. Nat. Commun. 2015, 6, 7453. [Google Scholar] [CrossRef]
- Mo, Z.; Luo, W.; Pi, K.; Duan, L.; Wang, P.; Ke, Y.; Zeng, S.; Jia, R.; Liang, T.; Huang, Y.; et al. Comparative transcriptome analysis between inbred lines and hybrids provides molecular insights into K+ content heterosis of tobacco (Nicotiana tabacum L.). Front. Plant Sci. 2022, 13, 940787. [Google Scholar] [CrossRef] [PubMed]
- Shull, G.H. A pure-line method in corn breeding. J. Hered. 1909, 1, 51–58. [Google Scholar] [CrossRef]
- Shull, G.H. Hybridization Methods in Corn Breeding. J. Hered. 1910, 1, 98–107. [Google Scholar] [CrossRef]
- Shull, G.H. The Genotypes of Maize. Am. Nat. 1911, 45, 234–252. [Google Scholar] [CrossRef]
- Jones, D.F. Dominance of Linked Factors as a Means of Accounting for Heterosis. Proc. Natl. Acad. Sci. USA 1917, 3, 310–312. [Google Scholar] [CrossRef] [PubMed]
- East, E.M. Heterosis. Genetics 1936, 21, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Krieger, U.; Lippman, Z.B.; Zamir, D. The flowering gene SINGLE FLOWER TRUSS drives heterosis for yield in tomato. Nat. Genet. 2010, 42, 459–463. [Google Scholar] [CrossRef]
- Stuber, C.W.; Lincoln, S.E.; Wolff, D.W.; Helentjaris, T.; Lander, E.S. Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers. Genetics 1992, 132, 823–839. [Google Scholar] [CrossRef]
- Graham, G.I.; Wolff, D.W.; Stuber, C.W. Characterization of a Yield Quantitative Trait Locus on Chromosome Five of Maize by Fine Mapping. Crop Sci. 1997, 37, 1601–1610. [Google Scholar] [CrossRef]
- Springer, N.M.; Stupar, R.M. Allelic variation and heterosis in maize: How do two halves make more than a whole? Genome Res. 2007, 17, 264–275. [Google Scholar] [CrossRef]
- Powers, L. An Expansion of Jones’s Theory for the Explanation of Heterosis. Am. Nat. 1944, 78, 275–280. [Google Scholar] [CrossRef]
- Monforte, A.J.; Tanksley, S.D. Fine mapping of a quantitative trait locus (QTL) from Lycopersicon hirsutum chromosome 1 affecting fruit characteristics and agronomic traits: Breaking linkage among QTLs affecting different traits and dissection of heterosis for yield. Theor. Appl. Genet. 2000, 100, 471–479. [Google Scholar] [CrossRef]
- Yu, C.; Wan, J.; Zhai, H.; Wang, C.; Jiang, L.; Xiao, Y.; Liu, Y. Study on heterosis of inter-subspecies between indica and japonica rice (Oryza sativa L.) using chromosome segment substitution lines. Chin. Sci. Bull. 2005, 50, 131–136. [Google Scholar] [CrossRef]
- Schmitz, R.J.; Schultz, M.D.; Urich, M.A.; Nery, J.R.; Pelizzola, M.; Libiger, O.; Alix, A.; McCosh, R.B.; Chen, H.; Schork, N.J.; et al. Patterns of population epigenomic diversity. Nature 2013, 495, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.T.; Spector, T.D. A twin approach to unraveling epigenetics. Trends Genet. 2011, 27, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Johannes, F.; Porcher, E.; Teixeira, F.K.; Saliba-Colombani, V.; Simon, M.; Agier, N.; Bulski, A.; Albuisson, J.; Heredia, F.; Audigier, P.; et al. Assessing the Impact of Transgenerational Epigenetic Variation on Complex Traits. PLoS Genet. 2009, 5, e1000530. [Google Scholar] [CrossRef]
- Dapp, M.; Reinders, J.; Bédiée, A.; Balsera, C.; Bucher, E.; Theiler, G.; Granier, C.; Paszkowski, J. Heterosis and inbreeding depression of epigenetic Arabidopsis hybrids. Nat. Plants 2015, 1, 15092. [Google Scholar] [CrossRef]
- Lauss, K.; Wardenaar, R.; Oka, R.; van Hulten, M.H.A.; Guryev, V.; Keurentjes, J.J.B.; Stam, M.; Johannes, F. Parental DNA Methylation States Are Associated with Heterosis in Epigenetic Hybrids. Plant Physiol. 2018, 176, 1627–1645. [Google Scholar] [CrossRef]
- Li, Q.; Eichten, S.R.; Hermanson, P.J.; Zaunbrecher, V.M.; Song, J.; Wendt, J.; Rosenbaum, H.; Madzima, T.F.; Sloan, A.E.; Huang, J.; et al. Genetic Perturbation of the Maize Methylome. Plant Cell 2015, 26, 4602–4616. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, N.; Xu, C.; Zhong, S.; Lin, X.; Yang, J.; Zhou, T.; Yuliang, A.; Wu, Y.; Chen, Y.-R.; et al. Mutation of a major CG methylase in rice causes genome-wide hypomethylation, dysregulated genome expression, and seedling lethality. Proc. Natl. Acad. Sci. USA 2014, 111, 10642–10647. [Google Scholar] [CrossRef]
- Yamauchi, T.; Johzuka-Hisatomi, Y.; Terada, R.; Nakamura, I.; Iida, S. The MET1b gene encoding a maintenance DNA methyltransferase is indispensable for normal development in rice. Plant Mol. Biol. 2014, 85, 219–232. [Google Scholar] [CrossRef]
- Miller, M.; Zhang, C.; Chen, Z.J. Ploidy and Hybridity Effects on Growth Vigor and Gene Expression in Arabidopsis thaliana Hybrids and Their Parents. G3 Genes Genomes Genet. 2012, 2, 505–513. [Google Scholar] [CrossRef]
- Fort, A.; Ryder, P.; McKeown, P.C.; Wijnen, C.; Aarts, M.G.; Sulpice, R.; Spillane, C. Disaggregating polyploidy, parental genome dosage and hybridity contributions to heterosis in Arabidopsis thaliana. New Phytol. 2016, 209, 590–599. [Google Scholar] [CrossRef]
- Castillo-Bravo, R.; Fort, A.; Cashell, R.; Brychkova, G.; McKeown, P.C.; Spillane, C. Parent-of-Origin Effects on Seed Size Modify Heterosis Responses in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 835219. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Gray, A.D.; Auger, D.L.; Birchler, J.A. Genomic dosage effects on heterosis in triploid maize. Proc. Natl. Acad. Sci. USA 2013, 110, 2665–2669. [Google Scholar] [CrossRef] [PubMed]
- Moll, R.H.; Lonnquist, J.H.; Fortuno, J.V.; Johnson, E.C. The Relationship of Heterosis and Genetic Divergence in Maize. Genetics 1965, 52, 139–144. [Google Scholar] [CrossRef]
- Lee, M.; Godshalk, E.B.; Lamkey, K.R.; Woodman, W.W. Association of Restriction Fragment Length Polymorphisms among Maize Inbreds with Agronomic Performance of Their Crosses. Crop Sci. 1989, 29, 1067–1071. [Google Scholar] [CrossRef]
- Smith, O.S.; Smith, J.S.C.; Bowen, S.L.; Tenborg, R.A.; Wall, S.J. Similarities among a group of elite maize inbreds as measured by pedigree, F1 grain yield, grain yield, heterosis, and RFLPs. Theor. Appl. Genet. 1990, 80, 833–840. [Google Scholar] [CrossRef]
- Benchimol, L.L.; de Souza, C.L.; Garcia, A.A.F.; Kono, P.M.S.; Mangolin, C.A.; Barbosa, A.M.M.; Coelho, A.S.G.; de Souza, A.P. Genetic diversity in tropical maize inbred lines: Heterotic group assignment and hybrid performance determined by RFLP markers. Plant Breed. 2000, 119, 491–496. [Google Scholar] [CrossRef]
- Riddle, N.C.; Birchler, J.A. Comparative analysis of inbred and hybrid maize at the diploid and tetraploid levels. Theor. Appl. Genet. 2008, 116, 563–576. [Google Scholar] [CrossRef]
- Frisch, M.; Thiemann, A.; Fu, J.; Schrag, T.A.; Scholten, S.; Melchinger, A.E. Transcriptome-based distance measures for grouping of germplasm and prediction of hybrid performance in maize. Theor. Appl. Genet. 2010, 120, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Reif, J.C.; Fischer, S.; Schrag, T.A.; Lamkey, K.R.; Klein, D.; Dhillon, B.S.; Utz, H.F.; Melchinger, A.E. Broadening the genetic base of European maize heterotic pools with US Cornbelt germplasm using field and molecular marker data. TAG Theor. Appl. Genet. Theor. Angew. Genet. 2010, 120, 301–310. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Lee, M.; Lamkey, K.R.; Hallauer, A.R.; Woodman, W.L. Genetic diversity for restriction fragment length polymorphisms and heterosis for two diallel sets of maize inbreds. Theor. Appl. Genet. 1990, 80, 488–496. [Google Scholar] [CrossRef]
- Geleta, L.F.; Labuschagne, M.T.; Viljoen, C.D. Relationship between heterosis and genetic distance based on morphological traits and AFLP markers in pepper. Plant Breed. 2004, 123, 467–473. [Google Scholar] [CrossRef]
- Zhang, Q.; Gao, Y.J.; Maroof, M.A.S.; Yang, S.H.; Li, J.X. Molecular divergence and hybrid performance in rice. Mol. Breed. 1995, 1, 133–142. [Google Scholar] [CrossRef]
- Cerna, F.J.; Cianzio, S.R.; Rafalski, A.; Tingey, S.; Dyer, D. Relationship between seed yield heterosis and molecular marker heterozygosity in soybean. Theor. Appl. Genet. 1997, 95, 460–467. [Google Scholar] [CrossRef]
- Pandey, S.K.; Dasgupta, T.; Rathore, A.; Vemula, A. Relationship of Parental Genetic Distance with Heterosis and Specific Combining Ability in Sesame (Sesamum indicum L.) Based on Phenotypic and Molecular Marker Analysis. Biochem. Genet. 2018, 56, 188–209. [Google Scholar] [CrossRef] [PubMed]
- Joyce, T.A.; Abberton, M.T.; Michaelson-Yeates, T.P.T.; Forster, J.W. Relationships between genetic distance measured by RAPD-PCR and heterosis in inbred lines of white clover (Trifolium repens L.). Euphytica 1999, 107, 159–165. [Google Scholar] [CrossRef]
- Sant, V.J.; Patankar, A.G.; Sarode, N.D.; Mhase, L.B.; Sainani, M.N.; Deshmukh, R.B.; Ranjekar, P.K.; Gupta, V.S. Potential of DNA markers in detecting divergence and in analysing heterosis in Indian elite chickpea cultivars. Theor. Appl. Genet. 1999, 98, 1217–1225. [Google Scholar] [CrossRef]
- Kaushik, P. Genetic Analysis for Fruit Phenolics Content, Flesh Color, and Browning Related Traits in Eggplant (Solanum melongena). Int. J. Mol. Sci. 2019, 20, 2990. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.S.; Gupta, S.K.; Marathi, B.; Danam, S.; Thatikunta, R.; Rathore, A.; Das, R.R.; Dangi, K.S.; Yadav, O.P. African and Asian origin pearl millet populations: Genetic diversity pattern and its association with yield heterosis. Crop Sci. 2020, 60, 3035–3048. [Google Scholar] [CrossRef]
- Zeid, M.; Schön, C.-C.; Link, W. Hybrid performance and AFLP- based genetic similarity in faba bean. Euphytica 2004, 139, 207–216. [Google Scholar] [CrossRef]
- Abdullah, N.; Yusop, M.R.; Ithnin, M.; Saleh, G.; Latif, M.A. Genetic variability of oil palm parental genotypes and performance of its’ progenies as revealed by molecular markers and quantitative traits. C. R. Biol. 2011, 334, 290–299. [Google Scholar] [CrossRef]
- Chandra, D.; Verma, S.; Gaur, A.; Bisht, C.; Gautam, A.; Chauhan, C.; Yadav, H. Heterosis, Combining Ability, Genetic Diversity and their Interrelationship in Pigeonpea [Cajanus cajan (L.) Millspaugh]. LEGUME Res.—Int. J. 2024, 47, 183–189. [Google Scholar] [CrossRef]
- Napolitano, M.; Terzaroli, N.; Kashyap, S.; Russi, L.; Jones-Evans, E.; Albertini, E. Exploring Heterosis in Melon (Cucumis melo L.). Plants 2020, 9, 282. [Google Scholar] [CrossRef] [PubMed]
- Teklewold, A.; Becker, H.C. Comparison of phenotypic and molecular distances to predict heterosis and F1 performance in Ethiopian mustard (Brassica carinata A. Braun). Theor. Appl. Genet. 2006, 112, 752–759. [Google Scholar] [CrossRef]
- Meyer, R.C.; Törjék, O.; Becher, M.; Altmann, T. Heterosis of Biomass Production in Arabidopsis. Establishment during Early Development. Plant Physiol. 2004, 134, 1813–1823. [Google Scholar] [CrossRef]
- Stokes, D.; Morgan, C.; O’Neill, C.; Bancroft, I. Evaluating the utility of Arabidopsis thaliana as a model for understanding heterosis in hybrid crops. Euphytica 2007, 156, 157–171. [Google Scholar] [CrossRef]
- Dias, L.A.D.S.; Kageyama, P.Y. Multivariate genetic divergence and hybrid performance of cacao (Theobroma cacao L.). Braz. J. Genet. 1997, 20, 63–70. [Google Scholar] [CrossRef]
- Dias, L.A.D.S.; Marita, J.; Cruz, C.D.; de Barros, E.G.; Salomão, T.M.F. Genetic distance and its association with heterosis in cacao. Braz. Arch. Biol. Technol. 2003, 46, 339–348. [Google Scholar] [CrossRef]
- Akpertey, A.; Padi, F.K.; Meinhardt, L.; Zhang, D. Relationship between genetic distance based on single nucleotide polymorphism markers and hybrid performance in Robusta coffee (Coffea canephora). Plant Breed. 2022, 141, 286–300. [Google Scholar] [CrossRef]
- Jordan, D.; Tao, Y.; Godwin, I.; Henzell, R.; Cooper, M.; McIntyre, C. Prediction of hybrid performance in grain sorghum using RFLP markers. Theor. Appl. Genet. 2003, 106, 559–567. [Google Scholar] [CrossRef]
- Cheres, M.T.; Miller, J.F.; Crane, J.M.; Knapp, S.J. Genetic distance as a predictor of heterosis and hybrid performance within and between heterotic groups in sunflower. Theor. Appl. Genet. 2000, 100, 889–894. [Google Scholar] [CrossRef]
- Usatov, A.V.; Klimenko, A.I.; Azarin, K.V.; Gorbachenko, O.F.; Markin, N.V.; Tikhobaeva, V.E.; Kolosov, Y.A.; Usatova, O.A.; Bakoev, S.; Makarenko, M.; et al. The relationship between heterosis and genetic distances based on ssr markers in Helianthus annuus. Am. J. Agric. Biol. Sci. 2014, 9, 270–276. [Google Scholar] [CrossRef]
- Sheeja, T.E.; Kumar, I.P.V.; Giridhari, A.; Minoo, D.; Rajesh, M.K.; Babu, K.N. Amplified Fragment Length Polymorphism: Applications and Recent Developments. Methods Mol. Biol. 2021, 2222, 187–218. [Google Scholar] [PubMed]
- Jagosz, B. The relationship between heterosis and genetic distances based on RAPD and AFLP markers in carrot. Plant Breed. 2011, 130, 574–579. [Google Scholar] [CrossRef]
- Karpechenko, G.D. Hybrids of Raphanus sativus L. × ♂Brassica oleracea L. J. Genet. 1924, 14, 375–396. [Google Scholar] [CrossRef]
- Yi, G.; Shin, H.; Park, H.R.; Park, J.E.; Ahn, J.H.; Lim, S.; Lee, J.G.; Lee, E.J.; Huh, J.H. Revealing biomass heterosis in the allodiploid xBrassicoraphanus, a hybrid between Brassica rapa and Raphanus sativus, through integrated transcriptome and metabolites analysis. BMC Plant Biol. 2020, 20, 252. [Google Scholar] [CrossRef]
- Mok, D.W.S.; Peloquin, S.J. Breeding value of 2n pollen (diplandroids) in tetraploid x diploid crosses in potatoes. Theor. Appl. Genet. 1975, 46, 307–314. [Google Scholar] [CrossRef]
- Groose, R.W.; Talbert, L.E.; Kojis, W.P.; Bingham, E.T. Progressive Heterosis in Autotetraploid Alfalfa: Studies Using Two Types of Inbreds. Crop Sci. 1989, 29, 1173–1177. [Google Scholar] [CrossRef]
- Kidwell, K.K.; Bingham, E.T.; Woodfield, D.R.; Osborn, T.C. Relationships among genetic distance, forage yield and heterozygosity in isogenic diploid and tetraploid alfalfa populations. Theor. Appl. Genet. 1994, 89, 323–328. [Google Scholar] [CrossRef]
- Sockness, B.A.; Dudley, J.W. Performance of Single and Double Cross Autotetraploid Maize Hybrids with Different Levels of Inbreeding. Crop Sci. 1989, 29, 875–879. [Google Scholar] [CrossRef]
- Tu, S.; Luan, L.; Liu, Y.; Long, W.; Kong, F.; He, T.; Xu, Q.; Yan, W.; Yu, M. Production and Heterosis Analysis of Rice Autotetraploid Hybrids. Crop Sci. 2007, 47, 2356–2363. [Google Scholar] [CrossRef]
- Wu, J.-W.; Hu, C.-Y.; Shahid, M.Q.; Guo, H.-B.; Zeng, Y.-X.; Liu, X.-D.; Lu, Y.-G. Analysis on genetic diversification and heterosis in autotetraploid rice. SpringerPlus 2013, 2, 439. [Google Scholar] [CrossRef] [PubMed]
- Diers, B.W.; McVetty, P.B.E.; Osborn, T.C. Relationship between Heterosis and Genetic Distance Based on Restriction Fragment Length Polymorphism Markers in Oilseed Rape (Brassica napus L.). Crop Sci. 1996, 36, 467–473. [Google Scholar] [CrossRef]
- Riaz, A.; Li, G.; Quresh, Z.; Swati, M.S.; Quiros, C.F. Genetic diversity of oilseed Brassica napus inbred lines based on sequence-related amplified polymorphism and its relation to hybrid performance. Plant Breed. 2001, 120, 411–415. [Google Scholar] [CrossRef]
- Ali, M.; Copeland, L.O.; Elias, S.G.; Kelly, J.D. Relationship between genetic distance and heterosis for yield and morphological traits in winter canola (Brassica napus L.). Theor. Appl. Genet. 1995, 91, 118–121. [Google Scholar] [CrossRef]
- Jansky, S. Chapter 2—Breeding, Genetics, and Cultivar Development. In Advances in Potato Chemistry and Technology; Singh, J., Kaur, L., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 27–62. [Google Scholar]
- Bonierbale, M.W.; Plaisted, R.L.; Tanksley, S.D. A test of the maximum heterozygosity hypothesis using molecular markers in tetraploid potatoes. Theor. Appl. Genet. 1993, 86, 481–491. [Google Scholar] [CrossRef]
- Demarly, Y. The concept of linkat. In Proceedings of the Broadening the Genetic Base of Crops, Wageningen, The Netherlands, 3–7 July 1979; pp. 257–265. [Google Scholar]
- Bingham, E.T.; Groose, R.W.; Woodfield, D.R.; Kidwell, K.K. Complementary Gene Interactions in Alfalfa are Greater in Autotetraploids than Diploids. Crop Sci. 1994, 34, 823–829. [Google Scholar] [CrossRef]
- Moody, M.E.; Mueller, L.D.; Soltis, D.E. Genetic variation and random drift in autotetraploid populations. Genetics 1993, 134, 649–657. [Google Scholar] [CrossRef]
- Busbice, T.H.; Wilsie, C.P. Inbreeding depression and heterosis in autotetraploids with application to Medicago sativa L. Euphytica 1966, 15, 52–67. [Google Scholar] [CrossRef]
- Levings, C.S.; Dudley, J.W.; Alexander, D.E. Inbreeding and Crossing in Autotetraploid Maize. Crop Sci. 1967, 7, 72–73. [Google Scholar] [CrossRef]
- Hecker, R.J. Inbreeding depression in diploid and autotetraploid sugarbeet, Beta vulgaris L. Euphytica 1972, 21, 106–111. [Google Scholar] [CrossRef]
- Lundqvist, A. Heterosis and Inbreeding Depression in Autotetraploid Rye. Hereditas 1966, 56, 317–366. [Google Scholar] [CrossRef]
- Lundqvist, A. Some Effects of Continued Inbreeding in an Autotetraploid Highbred Strain of Rye. Hereditas 1969, 61, 361–399. [Google Scholar] [CrossRef]
- Riddle, N.C.; Kato, A.; Birchler, J.A. Genetic variation for the response to ploidy change in Zea mays L. Theor. Appl. Genet. 2006, 114, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A. Interploidy hybridization barrier of endosperm as a dosage interaction. Front. Plant Sci. 2014, 5, 281. [Google Scholar] [CrossRef]
- Bingham, E.T.; Burnham, C.R.; Gates, C.E. Double and Single Backcross Linkage Estimates in Autotetraploid Maize. Genetics 1968, 59, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Fait, A.; Tel-Zur, N. Morphological, cytological and metabolic consequences of autopolyploidization in Hylocereus (Cactaceae) species. BMC Plant Biol. 2013, 13, 173. [Google Scholar] [CrossRef]
- Huang, Q.; Xiang, M.; Ji, B. Cytological observation on double fertilization of autotetraploid ric. J. Xiangtan Norm. Univ. (Nat. Sci. Ed.) 2000, 21, 84–87. [Google Scholar]
- Levin, D.A. Polyploidy and Novelty in Flowering Plants. Am. Nat. 1983, 122, 1–25. [Google Scholar] [CrossRef]
- Gallais, A. Quantitative Genetics and Breeding Methods in Autopolyploid Plants. Editions Quae: Versailles, France, 2003. [Google Scholar]
- Fischer, R.A. Wheat physiology at CIMMYT and raising the yield plateau. In Increasing Yield Potential in Wheat: Breaking the Barriers; CIMMYT: Ciudad Obregón, Mexico, 1996. [Google Scholar]
- Joshi, A.K.; Mishra, B.; Chatrath, R.; Ferrara, G.O.; Singh, R.P. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Dvorak, J. Genome Plasticity a Key Factor in the Success of Polyploid Wheat Under Domestication. Science 2007, 316, 1862–1866. [Google Scholar] [CrossRef]
- Van De Wouw, M.; Kik, C.; Van Hintum, T.; Van Treuren, R.; Visser, B. Genetic erosion in crops: Concept, research results and challenges. Plant Genet. Resour. 2010, 8, 1–15. [Google Scholar] [CrossRef]
- Hall, A.J.; Richards, R.A. Prognosis for genetic improvement of yield potential and water-limited yield of major grain crops. Field Crops Res. 2013, 143, 18–33. [Google Scholar] [CrossRef]
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Gumber, R.K. Overview of Heterosis and Heterotic Groups in Agronomic Crops. In CSSA Special Publications; Larnkey, K.R., Staub, J.E., Eds.; Crop Science Society of America: Madison, WI, USA, 1998; pp. 29–44. [Google Scholar]
- Dewey, D.R. The Genomic System of Classification as a Guide to Intergeneric Hybridization with the Perennial Triticeae. In Gene Manipulation in Plant Improvement; Gustafson, J.P., Ed.; Springer: Boston, MA, USA, 1984; pp. 209–279. [Google Scholar]
- Lu, B.-R.; Von Bothmer, R. Production and cytogenetic analysis of the intergeneric hybrids between nine Elymus species and common wheat (Triticum aestivum L.). Euphytica 1991, 58, 81–95. [Google Scholar] [CrossRef]
- Friebe, B.; Mukai, Y.; Gill, B.S.; Cauderon, Y. C-banding and in-situ hybridization analyses of Agropyron intermedium, a partial wheat x Ag. intermedium amphiploid, and six derived chromosome addition lines. Theor. Appl. Genet. 1992, 84, 899–905. [Google Scholar] [CrossRef]
- Li, Z.; Li, B.; Tong, Y. The contribution of distant hybridization with decaploid Agropyron elongatum to wheat improvement in China. J. Genet. Genom. 2008, 35, 451–456. [Google Scholar] [CrossRef]
- Jiang, Y.; Schmidt, R.H.; Zhao, Y.; Reif, J.C. A quantitative genetic framework highlights the role of epistatic effects for grain-yield heterosis in bread wheat. Nat. Genet. 2017, 49, 1741–1746. [Google Scholar] [CrossRef]
- Eudes, F. Triticale; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Goral, H.; Tyrka, M.; Spiss, L. Assessing genetic variation to predict the breeding value of winter triticale cultivars and lines. J. Appl. Genet. 2005, 2, 125–131. [Google Scholar]
- Tanaka, T.; Tamaki, H.; Ashikaga, K.; Fujii, H.; Yamada, T. Use of molecular marker diversity to increase forage yield in timothy (Phleum pratense L.). Plant Breed. 2013, 132, 144–148. [Google Scholar] [CrossRef]
- Hallahan, B.F.; Fernandez-Tendero, E.; Fort, A.; Ryder, P.; Dupouy, G.; Deletre, M.; Curley, E.; Brychkova, G.; Schulz, B.; Spillane, C. Hybridity has a greater effect than paternal genome dosage on heterosis in sugar beet (Beta vulgaris). BMC Plant Biol. 2018, 18, 120. [Google Scholar] [CrossRef]
- Luthra, S.K.; Gopal, J.; Sharma, P.C. Genetic divergence and its relationship with heterosis in potato. Potato J. 2005, 32, 37–42. [Google Scholar]
- Sanford, J.C.; Hanneman, R.E. A possible heterotic threshold in the potato and its implications for breeding. Theor. Appl. Genet. 1982, 61, 151–159. [Google Scholar] [CrossRef]
- Humphreys, M.O. A genetic approach to the multivariate differentiation of perennial ryegrass (Lolium perenne L.) populations. Heredity 1991, 66, 437–443. [Google Scholar] [CrossRef][Green Version]
- Marcón, F.; Martínez, E.J.; Rodríguez, G.R.; Zilli, A.L.; Brugnoli, E.A.; Acuña, C.A. Genetic distance and the relationship with heterosis and reproductive behavior in tetraploid bahiagrass hybrids. Mol. Breed. 2019, 39, 89. [Google Scholar] [CrossRef]
- Serapiglia, M.J.; Gouker, F.E.; Smart, L.B. Early selection of novel triploid hybrids of shrub willow with improved biomass yield relative to diploids. BMC Plant Biol. 2014, 14, 74. [Google Scholar] [CrossRef]
- Arunachalam, V.; Bandyopadhyay, A.; Nigam, S.N.; Gibbons, R.W. Heterosis in relation to genetic divergence and specific combining ability in groundnut (Arachis hypogaea L.). Euphytica 1984, 33, 33–39. [Google Scholar] [CrossRef]
- Mohammed, W. Magnitude of Exploitable Heterosis for Yield and Quality Traits of Coffee (Coffea arabica L.) Hybrids as Affected by Distant Parents in Origin and Morphology in Ethiopia. East Afr. J. Sci. 2011, 5, 22–36. [Google Scholar]
- Geng, X.; Qu, Y.; Jia, Y.; He, S.; Pan, Z.; Wang, L.; Du, X. Assessment of heterosis based on parental genetic distance estimated with SSR and SNP markers in upland cotton (Gossypium hirsutum L.). BMC Genom. 2021, 22, 123. [Google Scholar] [CrossRef]
- Martin, J.M.; Talbert, L.E.; Lanning, S.P.; Blake, N.K. Hybrid Performance in Wheat as Related to Parental Diversity. Crop Sci. 1995, 35, 104–108. [Google Scholar] [CrossRef]
- Barbosa-Neto, J.F.; Sorrells, M.E.; Cisar, G. Prediction of heterosis in wheat using coefficient of parentage and RFLP-based estimates of genetic relationship. Genome 1996, 39, 1142–1149. [Google Scholar] [CrossRef]
- Stupar, R.M.; Gardiner, J.M.; Oldre, A.G.; Haun, W.J.; Chandler, V.L.; Springer, N.M. Gene expression analyses in maize inbreds and hybrids with varying levels of heterosis. BMC Plant Biol. 2008, 8, 33. [Google Scholar] [CrossRef]
- Thiemann, A.; Fu, J.; Schrag, T.A.; Melchinger, A.E.; Frisch, M.; Scholten, S. Correlation between parental transcriptome and field data for the characterization of heterosis in Zea mays L. Theor. Appl. Genet. 2010, 120, 401–413. [Google Scholar] [CrossRef]
- Bernardo, R. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef]
- Heffner, E.L.; Sorrells, M.E.; Jannink, J.-L. Genomic Selection for Crop Improvement. Crop Sci. 2009, 49, 1–12. [Google Scholar] [CrossRef]
- Mangin, B.; Bonnafous, F.; Blanchet, N.; Boniface, M.-C.; Bret-Mestries, E.; Carrère, S.; Cottret, L.; Legrand, L.; Marage, G.; Pegot-Espagnet, P.; et al. Genomic Prediction of Sunflower Hybrids Oil Content. Front. Plant Sci. 2017, 8, 291012. [Google Scholar] [CrossRef]
- Technow, F.; Schrag, T.A.; Schipprack, W.; Bauer, E.; Simianer, H.; Melchinger, A.E. Genome Properties and Prospects of Genomic Prediction of Hybrid Performance in a Breeding Program of Maize. Genetics 2014, 197, 1343–1355. [Google Scholar] [CrossRef]
- Chester, M.; Leitch, A.R.; Soltis, P.S.; Soltis, D.E. Review of the Application of Modern Cytogenetic Methods (FISH/GISH) to the Study of Reticulation (Polyploidy/Hybridisation). Genes 2010, 1, 166–192. [Google Scholar] [CrossRef]
- Gärtner, T.; Steinfath, M.; Andorf, S.; Lisec, J.; Meyer, R.C.; Altmann, T.; Willmitzer, L.; Selbig, J. Improved Heterosis Prediction by Combining Information on DNA- and Metabolic Markers. PLoS ONE 2009, 4, e5220. [Google Scholar] [CrossRef]
- Korn, M.; Gärtner, T.; Erban, A.; Kopka, J.; Selbig, J.; Hincha, D.K. Predicting Arabidopsis Freezing Tolerance and Heterosis in Freezing Tolerance from Metabolite Composition. Mol. Plant 2010, 3, 224–235. [Google Scholar] [CrossRef]
- Steinfath, M.; Gärtner, T.; Lisec, J.; Meyer, R.C.; Altmann, T.; Willmitzer, L.; Selbig, J. Prediction of hybrid biomass in Arabidopsis thaliana by selected parental SNP and metabolic markers. TAG Theor. Appl. Genet. Theor. Angew. Genet. 2010, 120, 239–247. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).