
Prospects and Challenges of Bacteriophage Substitution for Antibiotics in Livestock and Poultry Production

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Brief Introduction of Bacteriophages
3. Application of Bacteriophage in Substituting Antibiotics in Livestock and Poultry Production

{kind=link}
{kind=link}
| Bacteriophage | Model | Effects | References |
|---|---|---|---|
| Mixture 1* | Weaned piglet | An amount of 400 mg/kg of bacteriophage mixture supplement significantly elevated villi height (VH)/crypt depth (CD) ratio (duodenum, jejunum, and ileum), significantly increased the relative mRNA expression of ZO-1, Claudin-1, and Occludin, and significantly decreased interleukin-1β and tumor necrosis factor-α in serum, as well as reduced diarrhea incidence. | Zeng et al. [31] |
| Cocktail 2* | Weanling pig | An amount of 1.0 g/kg of bacteriophage cocktail supplement significantly increased average daily gain, ileal L. spp. and villus height (duodenum and jejunum), and decreased relative abundance of Coliforms and Clostridium spp. (ileum). | Kim et al. [32] |
| Cocktail 3* | Barrows | An amount of 1.0 g/kg of bacteriophage cocktail supplement increased the average daily gain by 10.58%. With the increase in bacteriophage dose (0, 0.5, 1.0, and 1.5 g/kg), the abundance of Bifidobacterium spp. and L. spp. increased linearly, and the abundance of C. spp. and Coliforms decreased linearly in feces. | Kim et al. [33] |
| Cocktail 4* | Broiler chicken | An amount of 400 mg/kg of bacteriophage cocktail supplement significantly increased the relative abundance of Bifidobacterium, Prevotella, and L. salivarius; the relative abundance of L. aviarius decreased compared with antibiotic treatment (ileum). | Upadhaya et al. [37] |
| Cocktail 5* | Broiler chicken | An amount of 1.5 g/kg of bacteriophage cocktail supplement significantly increased the abundance of Lactobacillus (ileum), the serum concentrations of propionate (cecum), VH/CD, the serum concentrations of total antibody, immunoglobulin M (Ig M), and IgG, and thus significantly decreased the abundance of Coliform bacteria compared with the control group. | Sarrami et al. [39] |
| Mixture 6* | Laying hen | An amount of 10 mg/kg of bacteriophage mixture supplement significantly decreased the colonization of Salmonella typhimurium (spleen, oviduct, caecum, and excreta) and the mRNA expressions of IFNγ, HSP-27, and TNF-α compared with antibiotic treatment (liver). | Lee et al. [36] |
4. The Feasibility of Bacteriophages in Substituting Antibiotics
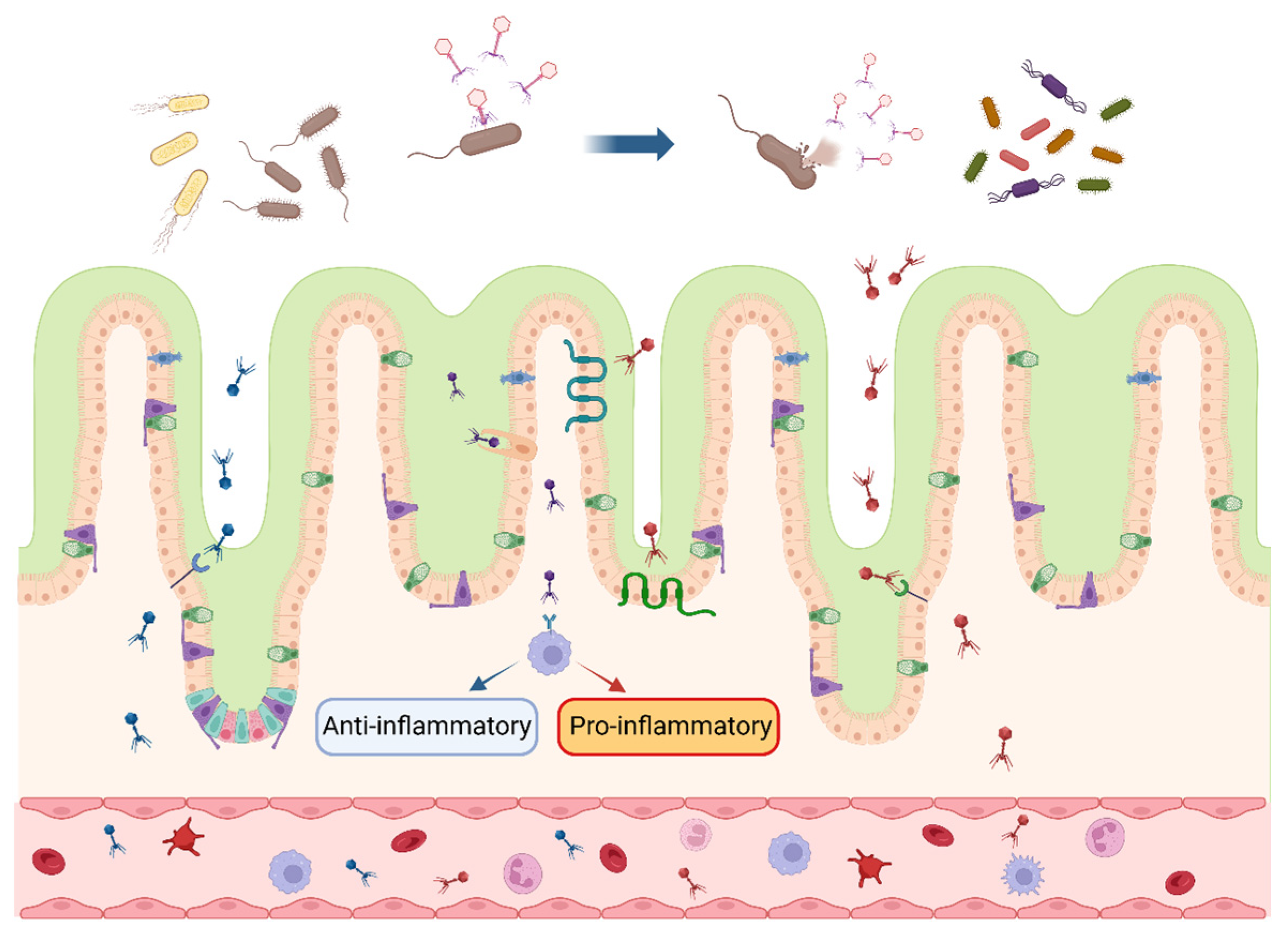
4.1. Regulation of Intestinal Microecology
4.2. Targeted Inhibition of Pathogens
4.3. Mediation of Immune Response
4.4. Improvement in Intestinal Morphology
5. The Challenge of Bacteriophage Application
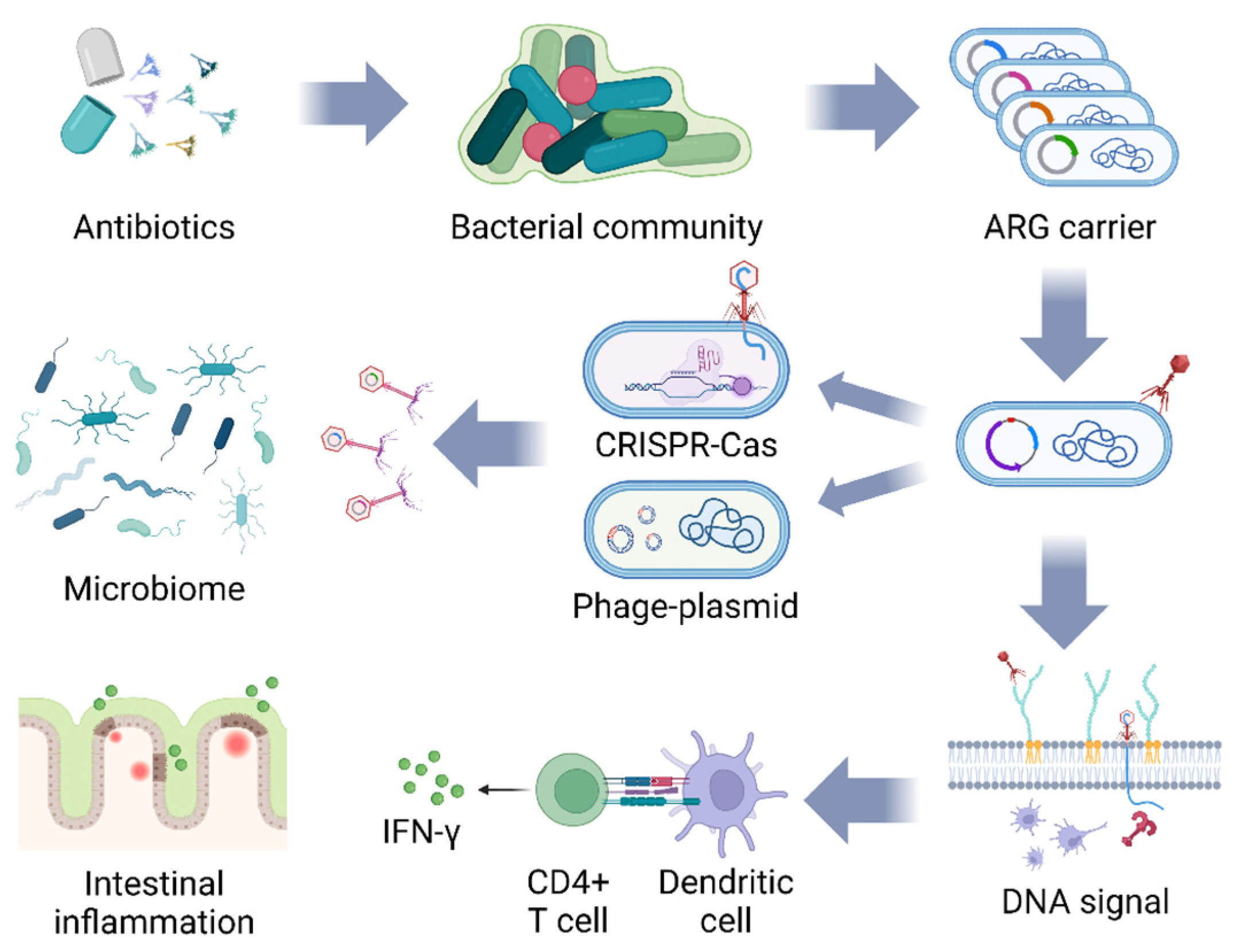
5.1. The Battle between Bacteriophage and Bacteria
5.2. Challenges of ARG Diffusion
5.3. The Instability of Bacteriophages
6. Prospects
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castanon, J.I. History of the use of antibiotic as growth promoters in european poultry feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.K.C.; Ng, R.W.Y.; Leung, S.S.Y.; Hui, M.; Ip, M. Overcoming the rising incidence and evolving mechanisms of antibiotic resistance by novel drug delivery approaches—An overview. Adv. Drug Deliv. Rev. 2022, 181, 114078. [Google Scholar] [CrossRef] [PubMed]
- Sabino, Y.N.V.; Santana, M.F.; Oyama, L.B.; Santos, F.G.; Moreira, A.J.S.; Huws, S.A.; Mantovani, H.C. Characterization of antibiotic resistance genes in the species of the rumen microbiota. Nat. Commun. 2019, 10, 5252. [Google Scholar] [CrossRef] [PubMed]
- Nguyet, L.T.Y.; Keeratikunakorn, K.; Kaeoket, K.; Ngamwongsatit, N. Antibiotic resistant Escherichia coli from diarrheic piglets from pig farms in thailand that harbor colistin-resistant mcr genes. Sci. Rep. 2022, 12, 9083. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, R.; Wang, N.; Deng, Y.; Tan, B.; Yin, Y.; Qi, M.; Wang, J. Ellagic acid alleviates oxidative stress by mediating nrf2 signaling pathways and protects against paraquat-induced intestinal injury in piglets. Antioxidants 2022, 11, 252. [Google Scholar] [CrossRef]
- Fu, Q.; Cui, Q.; Yang, Y.; Zhao, X.; Song, X.; Wang, G.; Bai, L.; Chen, S.; Tian, Y.; Zou, Y.; et al. Effect of resveratrol dry suspension on immune function of piglets. Evid. Based Complement. Alternat. Med. 2018, 2018, 5952707. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; He, Z.; Chen, Y.; Li, Z.; Meng, T.; Li, Y.; Cao, Y. In vitro and in vivo antioxidant activity of eucalyptus leaf polyphenols extract and its effect on chicken meat quality and cecum microbiota. Food Res. Int. 2020, 136, 109302. [Google Scholar] [CrossRef]
- Wang, H.; Wang, C.; Zou, Y.; Hu, J.; Li, Y.; Cheng, Y. Natural polyphenols in drug delivery systems: Current status and future challenges. Giant 2020, 3, 100022. [Google Scholar] [CrossRef]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. Probiotic strains as a potential tool for limiting the use of antibiotics, and improving the growth and health of pigs and chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- Leenay, R.T.; Vento, J.M.; Shah, M.; Martino, M.E.; Leulier, F.; Beisel, C.L. Genome editing with crispr-cas9 in lactobacillus plantarum revealed that editing outcomes can vary across strains and between methods. Biotechnol. J. 2019, 14, e1700583. [Google Scholar] [CrossRef]
- Wang, H.; Long, W.; Chadwick, D.; Zhang, X.; Zhang, S.; Piao, X.; Hou, Y. Dietary acidifiers as an alternative to antibiotics for promoting pig growth performance: A systematic review and meta-analysis. Anim. Feed. Sci. Technol. 2022, 289, 115320. [Google Scholar] [CrossRef]
- Rahman, M.R.T.; Fliss, I.; Biron, E. Insights in the development and uses of alternatives to antibiotic growth promoters in poultry and swine production. Antibiotics 2022, 11, 766. [Google Scholar] [CrossRef]
- Shkoporov, A.N.; Turkington, C.J.; Hill, C. Mutualistic interplay between bacteriophages and bacteria in the human gut. Nat. Rev. Microbiol. 2022, 20, 737–749. [Google Scholar] [CrossRef]
- Chevallereau, A.; Pons, B.J.; van Houte, S.; Westra, E.R. Interactions between bacterial and phage communities in natural environments. Nat. Rev. Microbiol. 2022, 20, 49–62. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef]
- Bao, J.; Wu, N.; Zeng, Y.; Chen, L.; Li, L.; Yang, L.; Zhang, Y.; Guo, M.; Li, L.; Li, J.; et al. Non-active antibiotic and bacteriophage synergism to successfully treat recurrent urinary tract infection caused by extensively drug-resistant Klebsiella pneumoniae. Emerg. Microbes Infect. 2020, 9, 771–774. [Google Scholar] [CrossRef]
- Cheng, R.; Li, X.; Jiang, L.; Gong, L.; Geslin, C.; Shao, Z. Virus diversity and interactions with hosts in deep-sea hydrothermal vents. Microbiome 2022, 10, 235. [Google Scholar] [CrossRef]
- Li, Z.; Pan, D.; Wei, G.; Pi, W.; Zhang, C.; Wang, J.H.; Peng, Y.; Zhang, L.; Wang, Y.; Hubert, C.R.J.; et al. Deep sea sediments associated with cold seeps are a subsurface reservoir of viral diversity. ISME J. 2021, 15, 2366–2378. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Mushegian, A.R. Are there 1031 virus particles on earth, or more, or fewer? J. Bacteriol. 2020, 202, e00052-20. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef]
- Yan, M.; Pratama, A.A.; Somasundaram, S.; Li, Z.; Jiang, Y.; Sullivan, M.B.; Yu, Z. Interrogating the viral dark matter of the rumen ecosystem with a global virome database. Nat. Commun. 2023, 14, 5254. [Google Scholar] [CrossRef]
- Tao, S.; Tao, H.; Li, J.; Wei, H. Landscapes of enteric virome signatures in early-weaned piglets. Microbiol. Spectr. 2022, 10, e01698-22. [Google Scholar] [CrossRef]
- Blanco-Picazo, P.; Gomez-Gomez, C.; Aguilo-Castillo, S.; Fernandez-Orth, D.; Cerda-Cuellar, M.; Muniesa, M.; Rodriguez-Rubio, L. Chicken liver is a potential reservoir of bacteriophages and phage-derived particles containing antibiotic resistance genes. Microb. Biotechnol. 2022, 15, 2464–2475. [Google Scholar] [CrossRef]
- Le, S.; Chen, L.G.; Zhu, T.Y. Chinese expert recommendation on phage therapy in the clinical practice. Chin. J. Infect. Dis. 2023, 41, 631–639. (In Chinese) [Google Scholar] [CrossRef]
- Quality, Safety and Efficacy of Bacteriophages as Veterinary Medicines—Scientific Guideline. Available online: https://www.ema.europa.eu/en/quality-safety-efficacy-bacteriophages-veterinary-medicines-scientific-guideline (accessed on 24 December 2023).
- Hampton, H.G.; Watson, B.N.J.; Fineran, P.C. The arms race between bacteria and their phage foes. Nature 2020, 577, 327–336. [Google Scholar] [CrossRef]
- Pires, D.P.; Melo, L.D.R.; Azeredo, J. Understanding the complex phage-host interactions in biofilm communities. Annu. Rev. Virol. 2021, 8, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Kredo-Russo, S.; Valdés-Mas, R.; Kviatcovsky, D.; Weinstock, E.; Matiuhin, Y.; Silberberg, Y.; Atarashi, K.; Furuichi, M.; Oka, A.; et al. Targeted suppression of human ibd-associated gut microbiota commensals by phage consortia for treatment of intestinal inflammation. Cell 2022, 185, 2879–2898.e2824. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wang, Z.; Zou, T.; Chen, J.; Li, G.; Zheng, L.; Li, S.; You, J. Bacteriophage as an alternative to antibiotics promotes growth performance by regulating intestinal inflammation, intestinal barrier function and gut microbiota in weaned piglets. Front. Vet. Sci. 2021, 8, 623899. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Hosseindoust, A.; Lee, S.H.; Choi, Y.H.; Kim, M.J.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage cocktail and multi-strain probiotics in the feed for weanling pigs: Effects on intestine morphology and targeted intestinal coliforms and clostridium. Animal 2017, 11, 45–53. [Google Scholar] [CrossRef]
- Kim, K.H.; Ingale, S.L.; Kim, J.S.; Lee, S.H.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage and probiotics both enhance the performance of growing pigs but bacteriophage are more effective. Anim. Feed Sci. Technol. 2014, 196, 88–95. [Google Scholar] [CrossRef]
- Xue, C.; Goldenfeld, N. Coevolution maintains diversity in the stochastic “kill the winner” model. Phys. Rev. Lett. 2017, 119, 268101. [Google Scholar] [CrossRef]
- Grabowski, L.; Wegrzyn, G.; Wegrzyn, A.; Podlacha, M. Phage therapy vs. The use of antibiotics in the treatment of salmonella-infected chickens: Comparison of effects on hematological parameters and selected biochemical markers. Antibiotics 2022, 11, 1787. [Google Scholar] [CrossRef]
- Lee, M.; Hosseindoust, A.; Oh, S.; Ko, H.; Cho, E.; Sa, S.; Kim, Y.; Choi, J.; Kim, J. Impact of an anti-salmonella. Typhimurium bacteriophage on intestinal microbiota and immunity status of laying hens. J. Anim. Physiol. Anim. Nutr. 2021, 105, 952–959. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Ahn, J.M.; Cho, J.H.; Kim, J.Y.; Kang, D.K.; Kim, S.W.; Kim, H.B.; Kim, I.H. Bacteriophage cocktail supplementation improves growth performance, gut microbiome and production traits in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 49. [Google Scholar] [CrossRef]
- Auffret, M.D.; Dewhurst, R.J.; Duthie, C.A.; Rooke, J.A.; John Wallace, R.; Freeman, T.C.; Stewart, R.; Watson, M.; Roehe, R. The rumen microbiome as a reservoir of antimicrobial resistance and pathogenicity genes is directly affected by diet in beef cattle. Microbiome 2017, 5, 159. [Google Scholar] [CrossRef]
- Sarrami, Z.; Sedghi, M.; Mohammadi, I.; Kim, W.K.; Mahdavi, A.H. Effects of bacteriophage supplement on the growth performance, microbial population, and pgc-1alpha and tlr4 gene expressions of broiler chickens. Sci. Rep. 2022, 12, 14391. [Google Scholar] [CrossRef]
- Chatterjee, A.; Willett, J.L.E.; Dunny, G.M.; Duerkop, B.A. Phage infection and sub-lethal antibiotic exposure mediate Enterococcus faecalis type vii secretion system dependent inhibition of bystander bacteria. PLoS Genet. 2021, 17, e1009204. [Google Scholar] [CrossRef] [PubMed]
- Hsu, B.B.; Gibson, T.E.; Yeliseyev, V.; Liu, Q.; Lyon, L.; Bry, L.; Silver, P.A.; Gerber, G.K. Dynamic modulation of the gut microbiota and metabolome by bacteriophages in a mouse model. Cell Host Microbe 2019, 25, 803–814.e805. [Google Scholar] [CrossRef] [PubMed]
- Vandenheuvel, D.; Lavigne, R.; Brussow, H. Bacteriophage therapy: Advances in formulation strategies and human clinical trials. Annu. Rev. Virol. 2015, 2, 599–618. [Google Scholar] [CrossRef]
- Ge, H.; Hu, M.; Zhao, G.; Du, Y.; Xu, N.; Chen, X.; Jiao, X. The “fighting wisdom and bravery” of tailed phage and host in the process of adsorption. Microbiol. Res. 2020, 230, 126344. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef]
- Wandro, S.; Ghatbale, P.; Attai, H.; Hendrickson, C.; Samillano, C.; Suh, J.; Dunham, S.J.B.; Pride, D.T.; Whitesona, K. Phage cocktails constrain the growth of enterococcus. Msystems 2022, 7, e00019-22. [Google Scholar] [CrossRef]
- Lee, H.Y.; Biswas, D.; Ahn, J. In-vitro adhesion and invasion properties of Salmonella typhimurium competing with bacteriophage in epithelial cells and chicken macrophages. Rev. Bras. Ciência Avícola 2015, 17, 427–432. [Google Scholar] [CrossRef]
- Saez, A.C.; Zhang, J.Y.; Rostagno, M.H.; Ebner, P.D. Direct feeding of microencapsulated bacteriophages to reduce salmonella colonization in pigs. Foodborne Pathog. Dis. 2011, 8, 1269–1274. [Google Scholar] [CrossRef]
- Seo, B.-J.; Song, E.-T.; Lee, K.; Kim, J.-W.; Jeong, C.-G.; Moon, S.-H.; Son, J.S.; Kang, S.H.; Cho, H.-S.; Jung, B.Y.; et al. Evaluation of the broad-spectrum lytic capability of bacteriophage cocktails against various salmonella serovars and their effects on weaned pigs infected with Salmonella typhimurium. J. Vet. Med. Sci. 2018, 80, 851–860. [Google Scholar] [CrossRef]
- Guo, M.; Gao, Y.; Xue, Y.; Liu, Y.; Zeng, X.; Cheng, Y.; Ma, J.; Wang, H.; Sun, J.; Wang, Z.; et al. Bacteriophage cocktails protect dairy cows against mastitis caused by drug resistant Escherichia coli infection. Front. Cell Infect. Microbiol. 2021, 11, 690377. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.W.; Huggins, M.B. Effectiveness of phages in treating experimental Escherichia coli diarrhoea in calves, piglets and lambs. J. Gen. Microbiol. 1983, 129, 2659–2675. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, A.R.; Fortier, A.; Roush, C.; Lessing, A.J.; Bender, R.G.; Barahman, R.; Grant, R.; Chan, B.K.; Turner, P.E. Pleiotropy complicates a trade-off between phage resistance and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 11207–11216. [Google Scholar] [CrossRef] [PubMed]
- Miernikiewicz, P.; Dabrowska, K. Endocytosis of bacteriophages. Curr. Opin. Virol. 2022, 52, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.; Baker, K.; Padman, B.S.; Patwa, R.; Dunstan, R.A.; Weston, T.A.; Schlosser, K.; Bailey, B.; Lithgow, T.; Lazarou, M.; et al. Bacteriophage transcytosis provides a mechanism to cross epithelial cell layers. mBio 2017, 8, e01874-17. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S.; et al. Bacteriophage adhering to mucus provide a non-host-derived immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Butcher, J.; Schramm, L.; Mack, D.R.; Stintzi, A. Multiomic spatial analysis reveals a distinct mucosa-associated virome. Gut Microbes 2023, 15, 2177488. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hosseindoust, A.; Goel, A.; Choi, Y.; Kwon, I.K.; Chae, B. Effects of dietary supplementation of bacteriophage with or without zinc oxide on the performance and gut development of weanling pigs. Ital. J. Anim. Sci. 2016, 15, 412–418. [Google Scholar] [CrossRef]
- Hosseindoust, A.R.; Lee, S.H.; Kim, J.S.; Choi, Y.H.; Noh, H.S.; Lee, J.H.; Jha, P.K.; Kwon, I.K.; Chae, B.J. Dietary bacteriophages as an alternative for zinc oxide or organic acids to control diarrhoea and improve the performance of weanling piglets. Veterinární Medicína 2017, 62, 53–61. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Y.; Lv, P.; Huang, J.; Tai, R.; Jin, X.; Wang, J.; Wang, X. Salmonella phages affect the intestinal barrier in chicks by altering the composition of early intestinal flora: Association with time of phage use. Front. Microbiol. 2022, 13, 947640. [Google Scholar] [CrossRef]
- Koskella, B.; Brockhurst, M.A. Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Hao, G.; Yuan, C.; Shu, R.; Jia, Y.; Zhao, S.; Xie, S.; Liu, M.; Zhou, H.; Sun, S.; Wang, H. O-antigen serves as a two-faced host factor for bacteriophage njs1 infecting nonmucoid Klebsiella pneumoniae. Microb. Pathog. 2021, 155, 104897. [Google Scholar] [CrossRef]
- Chatterjee, A.; Johnson, C.N.; Luong, P.; Hullahalli, K.; McBride, S.W.; Schubert, A.M.; Palmer, K.L.; Carlson, P.E., Jr.; Duerkop, B.A. Bacteriophage resistance alters antibiotic-mediated intestinal expansion of enterococci. Infect. Immun. 2019, 87, e00085-19. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.-J.; Henning, U. Superinfection exclusion by t-even-type coliphages. Trends Microbiol. 1994, 2, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Vasu, K.; Nagaraja, V. Diverse functions of restriction-modification systems in addition to cellular defense. Microbiol. Mol. Biol. Rev. 2013, 77, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. Crispr provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Blasco, L.; López-Hernández, I.; Rodríguez-Fernández, M.; Pérez-Florido, J.; Casimiro-Soriguer, C.S.; Djebara, S.; Merabishvili, M.; Pirnay, J.-P.; Rodríguez-Baño, J.; Tomás, M.; et al. Case report: Analysis of phage therapy failure in a patient with a Pseudomonas aeruginosa prosthetic vascular graft infection. Front. Med. 2023, 10, 1199657. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.R.; Dobias, D.T.; Weitz, J.S.; Barrick, J.E.; Quick, R.T.; Lenski, R.E. Repeatability and contingency in the evolution of a key innovation in phage lambda. Science 2012, 335, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Piya, D.; Vara, L.; Russell, W.K.; Young, R.; Gill, J.J. The multicomponent antirestriction system of phage p1 is linked to capsid morphogenesis. Mol. Microbiol. 2017, 105, 399–412. [Google Scholar] [CrossRef]
- Guo, T.W.; Bartesaghi, A.; Yang, H.; Falconieri, V.; Rao, P.; Merk, A.; Eng, E.T.; Raczkowski, A.M.; Fox, T.; Earl, L.A.; et al. Cryo-em structures reveal mechanism and inhibition of DNA targeting by a crispr-cas surveillance complex. Cell 2017, 171, 414–426.e412. [Google Scholar] [CrossRef]
- Huiting, E.; Cao, X.; Ren, J.; Athukoralage, J.S.; Luo, Z.; Silas, S.; An, N.; Carion, H.; Zhou, Y.; Fraser, J.S.; et al. Bacteriophages inhibit and evade cgas-like immune function in bacteria. Cell 2023, 186, 864–876.e821. [Google Scholar] [CrossRef]
- Camara-Wilpert, S.; Mayo-Munoz, D.; Russel, J.; Fagerlund, R.D.; Madsen, J.S.; Fineran, P.C.; Sorensen, S.J.; Pinilla-Redondo, R. Bacteriophages suppress crispr-cas immunity using rna-based anti-crisprs. Nature 2023, 623, 601–607. [Google Scholar] [CrossRef]
- Abdelrahman, F.; Easwaran, M.; Daramola, O.I.; Ragab, S.; Lynch, S.; Oduselu, T.J.; Khan, F.M.; Ayobami, A.; Adnan, F.; Torrents, E.; et al. Phage-encoded endolysins. Antibiotics 2021, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.Y.; Jang, I.J.; Yoon, S.; Jang, K.; Yu, K.-S.; Cho, J.Y.; Seong, M.-W.; Jung, G.M.; Yoon, S.J.; Kang, S.H. Pharmacokinetics and tolerance of the phage endolysin-based candidate drug sal200 after a single intravenous administration among healthy volunteers. Antimicrob. Agents Chemother. 2017, 61, e02629-16. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.Y.; Jung, G.M.; Yoon, S.J.; Oh, M.-D.; Choi, Y.-J.; Lee, W.J.; Kong, J.-C.; Seol, J.G.; Kang, S.H. Antibacterial properties of a pre-formulated recombinant phage endolysin, sal-1. Int. J. Antimicrob. Agents 2013, 41, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Hitch, T.C.A.; Thomas, B.J.; Friedersdorff, J.C.A.; Ougham, H.; Creevey, C.J. Deep sequence analysis reveals the ovine rumen as a reservoir of antibiotic resistance genes. Environ. Pollut. 2018, 235, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Barrangou, R. Crispr/cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Haaber, J.; Leisner, J.J.; Cohn, M.T.; Catalan-Moreno, A.; Nielsen, J.B.; Westh, H.; Penades, J.R.; Ingmer, H. Bacterial viruses enable their host to acquire antibiotic resistance genes from neighbouring cells. Nat. Commun. 2016, 7, 13333. [Google Scholar] [CrossRef]
- Leclerc, Q.J.; Wildfire, J.; Gupta, A.; Lindsay, J.A.; Knight, G.M. Growth-dependent predation and generalized transduction of antimicrobial resistance by bacteriophage. Msystems 2022, 7, e00135-22. [Google Scholar] [CrossRef]
- Pfeifer, E.; Moura de Sousa, J.A.; Touchon, M.; Rocha, E.P.C. Bacteria have numerous distinctive groups of phage-plasmids with conserved phage and variable plasmid gene repertoires. Nucleic Acids Res. 2021, 49, 2655–2673. [Google Scholar] [CrossRef]
- Pfeifer, E.; Bonnin, R.A.; Rocha, E.P.C. Phage-plasmids spread antibiotic resistance genes through infection and lysogenic conversion. mBio 2022, 13, e01851-22. [Google Scholar] [CrossRef]
- Price, V.J.; Huo, W.; Sharifi, A.; Palmer, K.L. Crispr-cas and restriction-modification act additively against conjugative antibiotic resistance plasmid transfer in Enterococcus faecalis. mSphere 2016, 1, e00064-16. [Google Scholar] [CrossRef] [PubMed]
- Yosef, I.; Manor, M.; Kiro, R.; Qimron, U. Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria. Proc. Natl. Acad. Sci. USA 2015, 112, 7267–7272. [Google Scholar] [CrossRef] [PubMed]
- Otto, L.; Chris, G.; Ernst, T.R. Endotoxins of gram-negative bacteria. Pharmacol. Ther. 1982, 15, 383–402. [Google Scholar]
- Ling, H.; Lou, X.; Luo, Q.; He, Z.; Sun, M.; Sun, J. Recent advances in bacteriophage-based therapeutics: Insight into the post-antibiotic era. Acta Pharm. Sin. B 2022, 12, 4348–4364. [Google Scholar] [CrossRef] [PubMed]
- Gogokhia, L.; Buhrke, K.; Bell, R.; Hoffman, B.; Brown, D.G.; Hanke-Gogokhia, C.; Ajami, N.J.; Wong, M.C.; Ghazaryan, A.; Valentine, J.F.; et al. Expansion of bacteriophages is linked to aggravated intestinal inflammation and colitis. Cell Host Microbe 2019, 25, 285–299.e288. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.V.; Ruggles, K.V.; Zhou, H.; Heguy, A.; Tsirigos, A.; Tetz, V. Bacteriophages as potential new mammalian pathogens. Sci. Rep. 2017, 7, 7043. [Google Scholar] [CrossRef]
- Tisza, M.J.; Buck, C.B. A catalog of tens of thousands of viruses from human metagenomes reveals hidden associations with chronic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023202118. [Google Scholar] [CrossRef]
- Norman, J.M.; Handley, S.A.; Baldridge, M.T.; Droit, L.; Liu, C.Y.; Keller, B.C.; Kambal, A.; Monaco, C.L.; Zhao, G.; Fleshner, P.; et al. Disease-specific alterations in the enteric virome in inflammatory bowel disease. Cell 2015, 160, 447–460. [Google Scholar] [CrossRef]
- Zuo, T.; Lu, X.-J.; Zhang, Y.; Cheung, C.P.; Lam, S.; Zhang, F.; Tang, W.; Ching, J.Y.L.; Zhao, R.; Chan, P.K.S.; et al. Gut mucosal virome alterations in ulcerative colitis. Gut 2019, 68, 1169–1179. [Google Scholar] [CrossRef]
- Van Belleghem, J.D.; Clement, F.; Merabishvili, M.; Lavigne, R.; Vaneechoutte, M. Pro- and anti-inflammatory responses of peripheral blood mononuclear cells induced by staphylococcus aureus and Pseudomonas aeruginosa phages. Sci. Rep. 2017, 7, 8004. [Google Scholar] [CrossRef]
- Gao, G.F.; Ji, H.; Wang, L.; Li, X. Fitness trade-offs in phage cocktail-resistant salmonella enterica serovar enteritidis results in increased antibiotic susceptibility and reduced virulence. Microbiol. Spectr. 2022, 10, e02914-22. [Google Scholar] [CrossRef] [PubMed]
- Pelyuntha, W.; Vongkamjan, K. Combined effects of salmonella phage cocktail and organic acid for controlling salmonella enteritidis in chicken meat. Food Control 2022, 133, 108653. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, J.; Mo, L.; Li, Y.; Li, Y.; Li, C.; Kuang, X.; Tao, Z.; Qu, Z.; Wu, L.; et al. Large-scale phage cultivation for commensal human gut bacteria. Cell Host Microbe 2023, 31, 665–677.e667. [Google Scholar] [CrossRef] [PubMed]
- Hatfull, G.F. Actinobacteriophages: Genomics, dynamics, and applications. Annu. Rev. Virol. 2020, 7, 37–61. [Google Scholar] [CrossRef]
- Apjok, G.; Szamel, M.; Christodoulou, C.; Seregi, V.; Vasarhelyi, B.M.; Stirling, T.; Eszenyi, B.; Sari, T.; Vidovics, F.; Nagrand, E.; et al. Characterization of antibiotic resistomes by reprogrammed bacteriophage-enabled functional metagenomics in clinical strains. Nat. Microbiol. 2023, 8, 410–423. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, A.; Liu, Z.; Lv, X.; Zhou, C.; Ran, T.; Tan, Z. Prospects and Challenges of Bacteriophage Substitution for Antibiotics in Livestock and Poultry Production. Biology 2024, 13, 28. https://doi.org/10.3390/biology13010028
Jiang A, Liu Z, Lv X, Zhou C, Ran T, Tan Z. Prospects and Challenges of Bacteriophage Substitution for Antibiotics in Livestock and Poultry Production. Biology. 2024; 13(1):28. https://doi.org/10.3390/biology13010028
Chicago/Turabian StyleJiang, Aoyu, Zixin Liu, Xiaokang Lv, Chuanshe Zhou, Tao Ran, and Zhiliang Tan. 2024. "Prospects and Challenges of Bacteriophage Substitution for Antibiotics in Livestock and Poultry Production" Biology 13, no. 1: 28. https://doi.org/10.3390/biology13010028
APA StyleJiang, A., Liu, Z., Lv, X., Zhou, C., Ran, T., & Tan, Z. (2024). Prospects and Challenges of Bacteriophage Substitution for Antibiotics in Livestock and Poultry Production. Biology, 13(1), 28. https://doi.org/10.3390/biology13010028