
Molecular Diet Analysis of Asian Clams for Supplementary Biodiversity Monitoring: A Case Study of Nakdong River Estuary



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
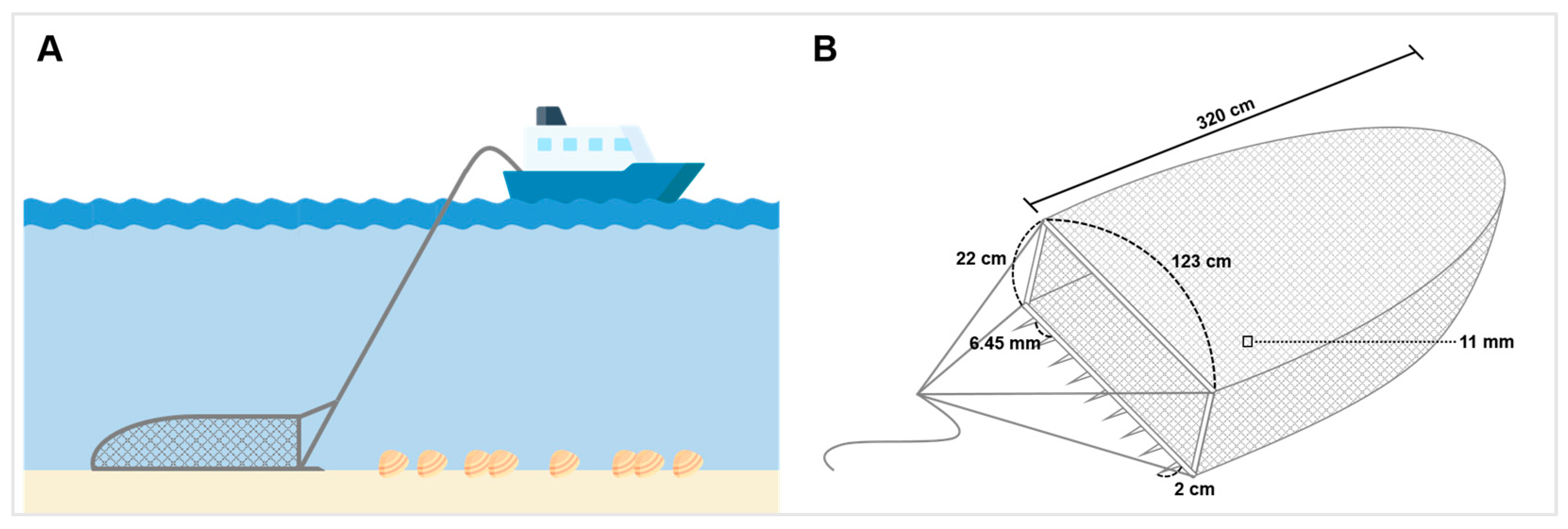
2.1. Study Site
2.2. Water Quality Survey
2.3. C. fluminea Sampling and Pretreatment
2.4. DNA Extraction and Amplification
2.5. DNA Sequence Analysis
2.6. Statistical Analysis and Reference Data
3. Results
3.1. Water Parameters
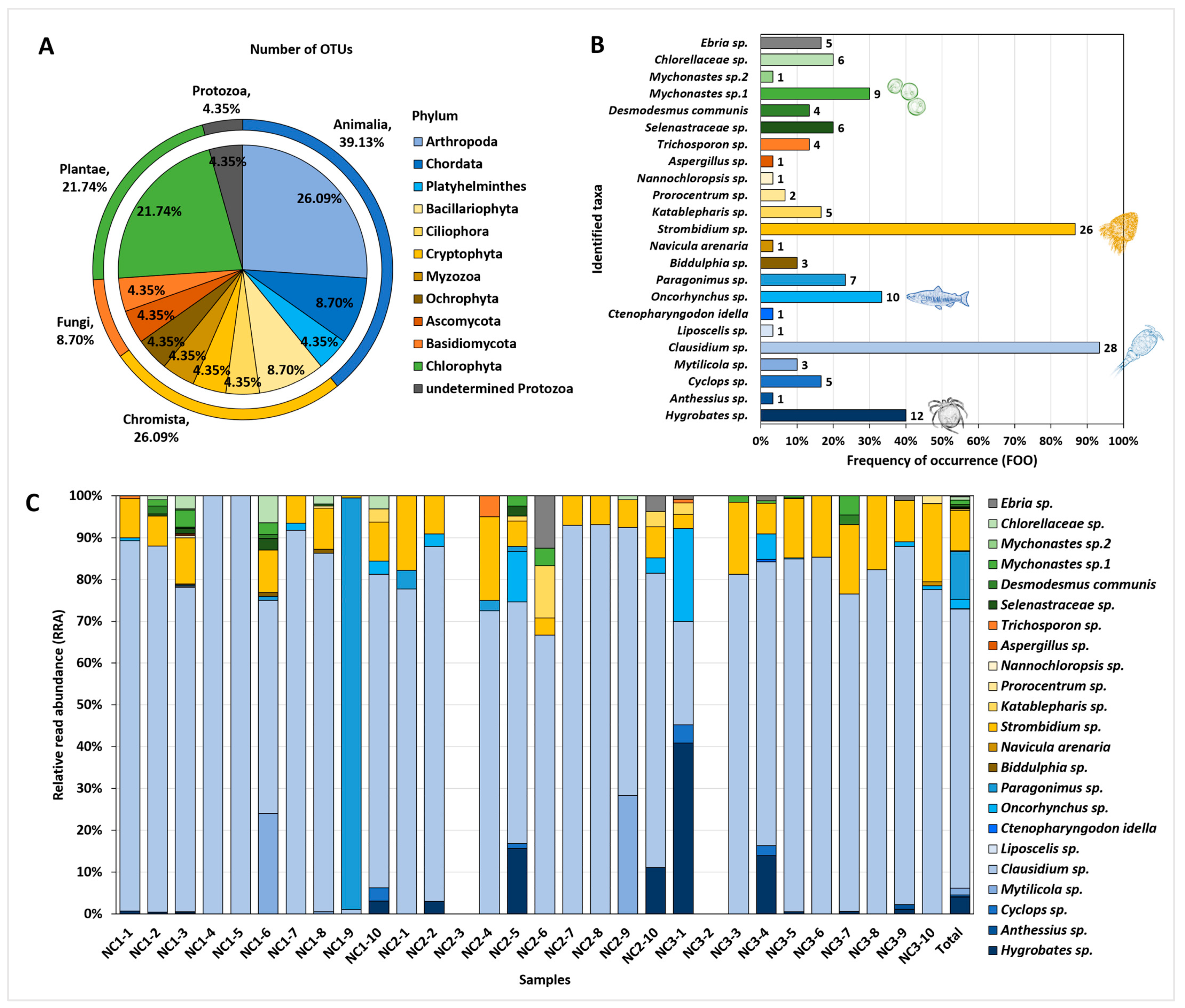
3.2. eDNA from the Gut Contents of C. fluminea
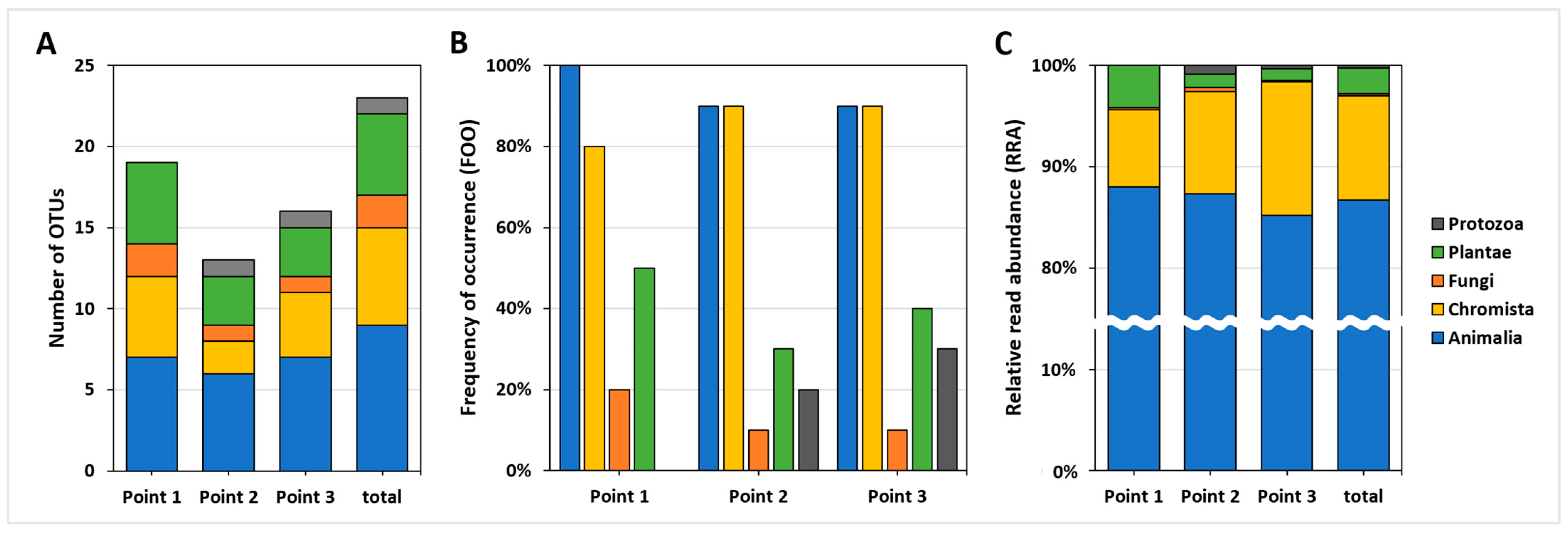
3.3. Comparison between Sampling Points
3.4. Comparison with Conventional Field Study
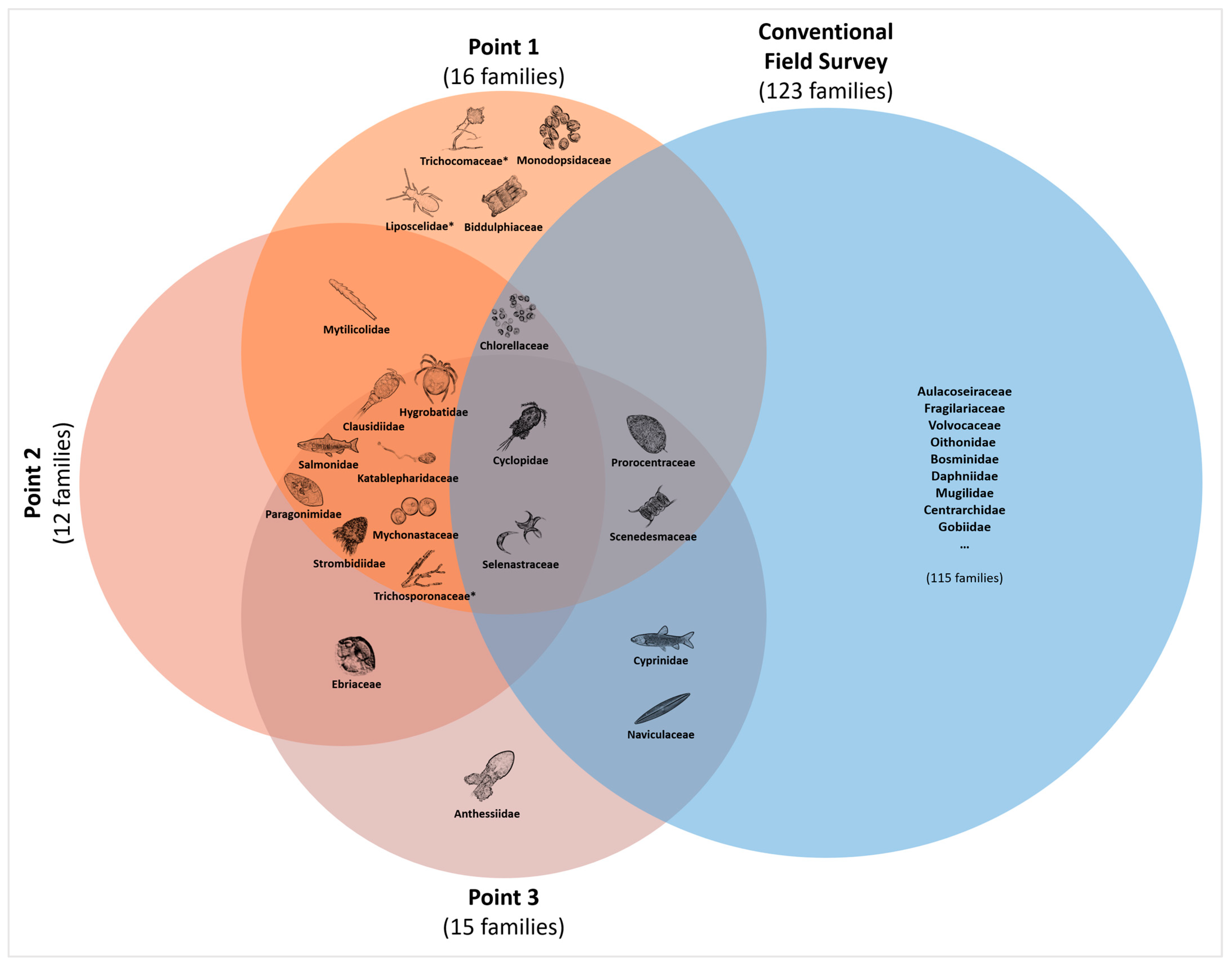
4. Discussion
4.1. Potential of C. fluminea as an eDNA Sampler for Supplementary Biodiversity Monitoring
4.2. Challenges of C. fluminea as an eDNA Sampler and Future Research
4.3. Limitations in Molecular Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Genus + Species | Habitat Environments |
|---|---|---|
| Arthropoda | Hygrobates sp. | marine, fresh 1 [33] |
| Anthessius sp. | marine | |
| Cyclops sp. | fresh | |
| Mytilicola sp. | marine | |
| Clausidium sp. | marine | |
| Liposcelis sp. | terrestrial | |
| Chordata | Ctenopharyngodon idella | brackish, fresh |
| Oncorhynchus sp. | marine, brackish, fresh | |
| Platyhelminthes | Paragonimus sp. | fresh, terrestrial |
| Bacillariophyta | Biddulphia sp. | marine |
| Navicula arenaria | marine, brackish | |
| Ciliophora | Strombidium sp. | marine, brackish, fresh |
| Cryptophyta | Katablepharis sp. | marine, fresh, terrestrial |
| Myzozoa | Prorocentrum sp. | marine, brackish, fresh |
| Ochrophyta | Nannochloropsis sp. | marine, brackish, fresh |
| Ascomycota | Aspergillus sp. | marine, brackish, fresh, terrestrial |
| Basidiomycota | Trichosporon sp. | marine, terrestrial |
| Chlorophyta | Selenastraceae sp. | fresh, terrestrial |
| Desmodesmus communis | fresh | |
| Mychonastes sp.1 | marine, fresh, terrestrial | |
| Mychonastes sp.2 | marine, fresh, terrestrial | |
| Chlorellaceae sp. | marine, brackish, fresh, terrestrial | |
| undetermined | Ebria sp. | marine, brackish |
| Source | df | SS | MS | Pseudo-F | P (Perm) | Unique Perms |
|---|---|---|---|---|---|---|
| Sampling sites | 2 | 2140.2 | 1070.1 | 0.74744 | 0.822 | 997 |
| Residuals | 27 | 38,655 | 1431.7 | |||
| Total | 29 | 40,795 |
References
- Levin, L.A.; Boesch, D.F.; Covich, A.; Dahm, C.; Erséus, C.; Ewel, K.C.; Kneib, R.T.; Moldenke, A.; Palmer, M.A.; Snelgrove, P.; et al. The Function of Marine Critical Transition Zones and the Importance of Sediment Biodiversity. Ecosystems 2001, 4, 430–451. [Google Scholar] [CrossRef]
- John, W.; Day, J.; Crump, B.C.; Kemp, W.M.; Yáñez-Arancibia, A. Estuarine Ecology, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem: Ecology, Threats and Management; OUP Oxford: Oxford, UK, 2004. [Google Scholar]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B.C. Depletion, Degradation, and Recovery Potential of Estuaries and Coastal Seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- Kennish, M.J. Environmental threats and environmental future of estuaries. Environ. Conserv. 2002, 29, 78–107. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D. Drivers of change in estuarine-coastal ecosystems: Discoveries from four decades of study in San Francisco Bay. Rev. Geophys. 2012, 50, RG4001. [Google Scholar] [CrossRef]
- Hobbs, J.; Bright, D. Environmental DNA: Implementation for Resource Development Projects in BC and Beyond; University of British Columbia Library: Vancouver, BC Canada, 2016. [Google Scholar]
- Zimmermann, J.; Glöckner, G.; Jahn, R.; Enke, N.; Gemeinholzer, B. Metabarcoding vs. morphological identification to assess diatom diversity in environmental studies. Mol. Ecol. Resour. 2015, 15, 526–542. [Google Scholar] [CrossRef]
- Ko, H.-L.; Wang, Y.-T.; Chiu, T.-S.; Lee, M.-A.; Leu, M.-Y.; Chang, K.-Z.; Chen, W.-Y.; Shao, K.-T. Evaluating the Accuracy of Morphological Identification of Larval Fishes by Applying DNA Barcoding. PLoS ONE 2013, 8, e53451. [Google Scholar] [CrossRef] [PubMed]
- Polanco, F.A.; Mutis Martinezguerra, M.; Marques, V.; Villa-Navarro, F.; Borrero Pérez, G.H.; Cheutin, M.-C.; Dejean, T.; Hocdé, R.; Juhel, J.-B.; Maire, E.; et al. Detecting aquatic and terrestrial biodiversity in a tropical estuary using environmental DNA. Biotropica 2021, 53, 1606–1619. [Google Scholar] [CrossRef]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA: For Biodiversity Research and Monitoring; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Beng, K.C.; Corlett, R.T. Applications of environmental DNA (eDNA) in ecology and conservation: Opportunities, challenges and prospects. Biodivers. Conserv. 2020, 29, 2089–2121. [Google Scholar] [CrossRef]
- Rivera, S.F.; Rimet, F.; Vasselon, V.; Vautier, M.; Domaizon, I.; Bouchez, A. Fish eDNA metabarcoding from aquatic biofilm samples: Methodological aspects. Mol. Ecol. Resour. 2022, 22, 1440–1453. [Google Scholar] [CrossRef]
- Tran, B.T.; Kim, K.-Y.; Heo, J.S.; Kim, K.-S.; Lee, H.J.; Park, T.G. Dietary analysis of three important mariculture species in South Korea using DNA metabarcoding in fecal samples. Aquac. Rep. 2023, 30, 101606. [Google Scholar] [CrossRef]
- Curran, T.G.; Browett, S.S.; O’Neill, D.; O’Hanlon, A.; O’Reilly, C.; Harrington, A.P.; McDevitt, A.D.; O’Meara, D.B. One bat’s waste is another man’s treasure: A DNA metabarcoding approach for the assessment of biodiversity and ecosystem services in Ireland using bat faeces. Biodivers. Conserv. 2022, 31, 2699–2722. [Google Scholar] [CrossRef]
- Nørgaard, L.; Olesen, C.R.; Trøjelsgaard, K.; Pertoldi, C.; Nielsen, J.L.; Taberlet, P.; Ruiz-González, A.; De Barba, M.; Iacolina, L. eDNA metabarcoding for biodiversity assessment, generalist predators as sampling assistants. Sci. Rep. 2021, 11, 6820. [Google Scholar] [CrossRef]
- Weber, S.; Junk, I.; Brink, L.; Wörner, M.; Künzel, S.; Veith, M.; Teubner, D.; Klein, R.; Paulus, M.; Krehenwinkel, H. Molecular diet analysis in mussels and other metazoan filter feeders and an assessment of their utility as natural eDNA samplers. Mol. Ecol. Resour. 2023, 23, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Siegenthaler, A.; Wangensteen, O.S.; Soto, A.Z.; Benvenuto, C.; Corrigan, L.; Mariani, S. Metabarcoding of shrimp stomach content: Harnessing a natural sampler for fish biodiversity monitoring. Mol. Ecol. Resour. 2019, 19, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Jeunen, G.-J.; Cane, J.S.; Ferreira, S.; Strano, F.; von Ammon, U.; Cross, H.; Day, R.; Hesseltine, S.; Ellis, K.; Urban, L.; et al. Assessing the utility of marine filter feeders for environmental DNA (eDNA) biodiversity monitoring. Mol. Ecol. Resour. 2023, 23, 771–786. [Google Scholar] [CrossRef]
- Mariani, S.; Baillie, C.; Colosimo, G.; Riesgo, A. Sponges as natural environmental DNA samplers. Curr. Biol. 2019, 29, R401–R402. [Google Scholar] [CrossRef]
- Gutiérrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Yoon, J.-D.; Jang, M.-H.; Jo, H.-B.; Jeong, K.-S.; Kim, G.-Y.; Joo, G.-J. Changes of fish assemblages after construction of an estuary barrage in the lower Nakdong River, South Korea. Limnology 2016, 17, 183–197. [Google Scholar] [CrossRef]
- Lee, C.-W.; Jang, J.-D.; Jeong, K.-S.; Kim, D.-K.; Joo, G.-J. Patterning habitat preference of avifaunal assemblage on the Nakdong River estuary (South Korea) using self-organizing map. Ecol. Inform. 2010, 5, 89–96. [Google Scholar] [CrossRef]
- Busan Marine Fisheries Resources Research Institute. Available online: https://www.busan.go.kr/fisheries/index (accessed on 16 August 2023).
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012; Volume 10. [Google Scholar]
- Rak, A.; Khalid, N.; Syed Omar, S. The distribution and length size of Corbicula fluminea (ETAK) in Sungai Pergau at Gunung Reng. Int. J. Eng. Technol. 2018, 7, 279–281. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- National Institute of Biological Resources. Available online: https://species.nibr.go.kr/index.do (accessed on 21 July 2023).
- World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org/ (accessed on 21 July 2023).
- National Center for Biotechnology Information (NCBI). Taxonomy Browser. Available online: https://www.ncbi.nlm.nih.gov/datasets/taxonomy/tree/ (accessed on 21 July 2023).
- Pešić, V.; Jovanović, M.; Manović, A.; Zawal, A.; Bańkowska, A.; Broda, Ł.; Martin, P.; Dabert, M. Two new species from the Hygrobates nigromaculatus-complex (Acariformes, Hydrachnidia, Hygrobatidae), based on morphological and molecular evidence. Acarologia 2020, 60, 753–768. [Google Scholar] [CrossRef]
- Deagle, B.E.; Thomas, A.C.; McInnes, J.C.; Clarke, L.J.; Vesterinen, E.J.; Clare, E.L.; Kartzinel, T.R.; Eveson, J.P. Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Mol. Ecol. 2019, 28, 391–406. [Google Scholar] [CrossRef] [PubMed]
- K-Water, Report of Water Quality and Aquatic Ecosystem Monitoring at Nakdong River Estuary; Korea Water Resources Corporation: Busan, Republic of Korea, 2022.
- Sousa, R.; Antunes, C.; Guilhermino, L. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: An overview. Ann. Limnol.-Int. J. Limnol. 2008, 44, 85–94. [Google Scholar] [CrossRef]
- Su, L.; Cai, H.; Kolandhasamy, P.; Wu, C.; Rochman, C.M.; Shi, H. Using the Asian clam as an indicator of microplastic pollution in freshwater ecosystems. Environ. Pollut. 2018, 234, 347–355. [Google Scholar] [CrossRef]
- Li, Z.; He, X.; Feng, C. A review of freshwater benthic clams (Corbicula fluminea): Accumulation capacity, underlying physiological mechanisms and environmental applications. Sci. Total Environ. 2023, 857, 159431. [Google Scholar] [CrossRef] [PubMed]
- Way, C.M.; Hornbach, D.J.; Miller-Way, C.A.; Payne, B.S.; Miller, A.C. Dynamics of filter feeding in Corbicula fluminea (Bivalvia: Corbiculidae). Can. J. Zool. 1990, 68, 115–120. [Google Scholar] [CrossRef]
- Schabacker, J.C.; Amish, S.J.; Ellis, B.K.; Gardner, B.; Miller, D.L.; Rutledge, E.A.; Sepulveda, A.J.; Luikart, G. Increased eDNA detection sensitivity using a novel high-volume water sampling method. Environ. DNA 2020, 2, 244–251. [Google Scholar] [CrossRef]
- Sepulveda, A.J.; Schabacker, J.; Smith, S.; Al-Chokhachy, R.; Luikart, G.; Amish, S.J. Improved detection of rare, endangered and invasive trout in using a new large-volume sampling method for eDNA capture. Environ. DNA 2019, 1, 227–237. [Google Scholar] [CrossRef]
- Crespo, D.; Dolbeth, M.; Leston, S.; Sousa, R.; Pardal, M.Â. Distribution of Corbicula fluminea (Müller, 1774) in the invaded range: A geographic approach with notes on species traits variability. Biol. Invasions 2015, 17, 2087–2101. [Google Scholar] [CrossRef]
- Gosling, E. Bivalve Molluscs: Biology, Ecology and Culture; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Xin, Y.; Guo, Y.; Sun, M.; Yu, G.; Ma, Z.; Pei, K.; Qin, C. Optimal Conditions to Quantify the Relationship between eDNA Concentration and Biomass in Acanthopagrus latus. Water 2022, 14, 3521. [Google Scholar] [CrossRef]
- Lauritsen, D.D. Filter-Feeding in Corbicula fluminea and Its Effect on Seston Removal. J. N. Am. Benthol. Soc. 1986, 5, 165–172. [Google Scholar] [CrossRef]
- Telesh, I.V.; Khlebovich, V.V. Principal processes within the estuarine salinity gradient: A review. Mar. Pollut. Bull. 2010, 61, 149–155. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D.; Schraga, T.S.; Nejad, E.; Martin, C. Ecosystem variability along the estuarine salinity gradient: Examples from long-term study of San Francisco Bay. Limnol. Oceanogr. 2017, 62, S272–S291. [Google Scholar] [CrossRef]
- Franco, T.P.; Neves, L.M.; Araújo, F.G. Better with more or less salt? The association of fish assemblages in coastal lagoons with different salinity ranges. Hydrobiologia 2019, 828, 83–100. [Google Scholar] [CrossRef]
- Matela, M.; Obolewski, K. Structural diagnosis of benthic invertebrate communities in relation to salinity gradient in Baltic coastal lake ecosystems using biological trait analysis. Sci. Rep. 2022, 12, 12750. [Google Scholar] [CrossRef]
- Heo, Y.-J.; Jo, H.; Kim, J.Y.; Kim, G.-Y.; Joo, G.-J.; Kim, H.-W. Application of DNA Metabarcoding for Identifying the Diet of Asian Clam (Corbicula fluminea, Müller, 1774). Sustainability 2023, 15, 441. [Google Scholar] [CrossRef]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef]
- Nagarajan, R.P.; Bedwell, M.; Holmes, A.E.; Sanches, T.; Acuña, S.; Baerwald, M.; Barnes, M.A.; Blankenship, S.; Connon, R.E.; Deiner, K.; et al. Environmental DNA Methods for Ecological Monitoring and Biodiversity Assessment in Estuaries. Estuaries Coasts 2022, 45, 2254–2273. [Google Scholar] [CrossRef]
- Hunter, M.E.; Ferrante, J.A.; Meigs-Friend, G.; Ulmer, A. Improving eDNA yield and inhibitor reduction through increased water volumes and multi-filter isolation techniques. Sci. Rep. 2019, 9, 5259. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lawson Handley, L.-J.; Read, D.S.; Hänfling, B. The effect of filtration method on the efficiency of environmental DNA capture and quantification via metabarcoding. Mol. Ecol. Resour. 2018, 18, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Vestheim, H.; Jarman, S.N. Blocking primers to enhance PCR amplification of rare sequences in mixed samples—A case study on prey DNA in Antarctic krill stomachs. Front. Zool. 2008, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Leray, M.; Agudelo, N.; Mills, S.C.; Meyer, C.P. Effectiveness of Annealing Blocking Primers versus Restriction Enzymes for Characterization of Generalist Diets: Unexpected Prey Revealed in the Gut Contents of Two Coral Reef Fish Species. PLoS ONE 2013, 8, e58076. [Google Scholar] [CrossRef]
- Blankenship, L.E.; Yayanos, A.A. Universal primers and PCR of gut contents to study marine invertebrate diets. Mol. Ecol. 2005, 14, 891–899. [Google Scholar] [CrossRef]
- Piñol, J.; Mir, G.; Gomez-Polo, P.; Agustí, N. Universal and blocking primer mismatches limit the use of high-throughput DNA sequencing for the quantitative metabarcoding of arthropods. Mol. Ecol. Resour. 2015, 15, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A Method for Studying Protistan Diversity Using Massively Parallel Sequencing of V9 Hypervariable Regions of Small-Subunit Ribosomal RNA Genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Albaina, A.; Aguirre, M.; Abad, D.; Santos, M.; Estonba, A. 18S rRNA V9 metabarcoding for diet characterization: A critical evaluation with two sympatric zooplanktivorous fish species. Ecol. Evol. 2016, 6, 1809–1824. [Google Scholar] [CrossRef]
- Hugerth, L.W.; Muller, E.E.L.; Hu, Y.O.O.; Lebrun, L.A.M.; Roume, H.; Lundin, D.; Wilmes, P.; Andersson, A.F. Systematic Design of 18S rRNA Gene Primers for Determining Eukaryotic Diversity in Microbial Consortia. PLoS ONE 2014, 9, e95567. [Google Scholar] [CrossRef]
- Questel, J.M.; Hopcroft, R.R.; DeHart, H.M.; Smoot, C.A.; Kosobokova, K.N.; Bucklin, A. Metabarcoding of zooplankton diversity within the Chukchi Borderland, Arctic Ocean: Improved resolution from multi-gene markers and region-specific DNA databases. Mar. Biodivers. 2021, 51, 4. [Google Scholar] [CrossRef]

| DO (mg L−1) | DO (%) | pH | Temp. (°C) | Conduc. (µS cm−1) | Salinity (ppt) | Alkal. (mg L−1) | Tur. (NTU) | |
|---|---|---|---|---|---|---|---|---|
| Point 1 | 7.00 | 83.5 | 7.83 | 24.1 | 16,716 | 10.0 | 84 | 3.68 |
| Point 2 | 6.46 | 77.5 | 7.97 | 24.3 | 21,666 | 14.2 | 90 | 3.68 |
| Point 3 | 6.73 | 80.7 | 7.99 | 24.5 | 20,436 | 12.3 | 84 | 3.97 |
| Kingdom | Phylum | Genus + Species | Max Score | Identity (%) | Query (%) | Genbank Accession | Level |
|---|---|---|---|---|---|---|---|
| Animalia | Arthropoda | Hygrobates sp. 1 | 224 | 90.06 | 100 | LC552029.1 | genus |
| Anthessius sp. | 259 | 93.18 | 100 | AY627002.1 | genus | ||
| Cyclops sp. | 302 | 97.73 | 100 | AY626998.1 | genus | ||
| Mytilicola sp. | 309 | 98.30 | 100 | AY627005.1 | genus | ||
| Clausidium sp. | 270 | 94.83 | 100 | JF781553.1 | genus | ||
| Liposcelis sp. | 244 | 96.00 | 81 | AY077779.1 | genus | ||
| Chordata | Ctenopharyngodon idella | 318 | 98.34 | 100 | XR_007928648.1 | species | |
| Oncorhynchus sp. | 327 | 99.44 | 100 | XR_008060685.1 | genus | ||
| Platyhelminthes | Paragonimus sp. | 283 | 95.03 | 100 | LT855189.1 | genus | |
| Chromista | Bacillariophyta | Biddulphia sp. | 276 | 96.95 | 94 | JX401228.1 | genus |
| Navicula arenaria | 322 | 100.00 | 100 | KJ961668.1 | species | ||
| Ciliophora | Strombidium sp. | 298 | 99.39 | 93 | MZ823795.1 | genus | |
| Cryptophyta | Katablepharis sp. | 324 | 99.44 | 100 | KJ925151.1 | genus | |
| Myzozoa | Prorocentrum sp. | 270 | 94.83 | 100 | MK405477.1 | genus | |
| Ochrophyta | Nannochloropsis sp. | 305 | 99.40 | 100 | KU900229.1 | genus | |
| Fungi | Ascomycota | Aspergillus sp. | 320 | 99.43 | 100 | NG_063229.1 | genus |
| Basidiomycota | Trichosporon sp. | 320 | 99.43 | 100 | MN268783.1 | genus | |
| Plantae | Chlorophyta | Selenastraceae sp. | 309 | 98.84 | 100 | KT833591.1 | family |
| Desmodesmus communis | 316 | 99.43 | 100 | KF864475.1 | species | ||
| Mychonastes sp.1 | 311 | 98.85 | 100 | OM415709.1 | genus | ||
| Mychonastes sp.2 | 316 | 99.43 | 100 | X73996.1 | genus | ||
| Chlorellaceae sp. | 270 | 94.29 | 100 | AJ131691.1 | family | ||
| Protozoa | undetermined | Ebria sp. | 255 | 92.74 | 100 | DQ303923.1 | genus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Joo, G.-J.; Jeong, K.-S.; Gim, J.-S.; Lee, Y.; Hong, D.; Jo, H. Molecular Diet Analysis of Asian Clams for Supplementary Biodiversity Monitoring: A Case Study of Nakdong River Estuary. Biology 2023, 12, 1245. https://doi.org/10.3390/biology12091245
Kim K, Joo G-J, Jeong K-S, Gim J-S, Lee Y, Hong D, Jo H. Molecular Diet Analysis of Asian Clams for Supplementary Biodiversity Monitoring: A Case Study of Nakdong River Estuary. Biology. 2023; 12(9):1245. https://doi.org/10.3390/biology12091245
Chicago/Turabian StyleKim, Kanghui, Gea-Jae Joo, Kwang-Seuk Jeong, Jeong-Soo Gim, Yerim Lee, Donghyun Hong, and Hyunbin Jo. 2023. "Molecular Diet Analysis of Asian Clams for Supplementary Biodiversity Monitoring: A Case Study of Nakdong River Estuary" Biology 12, no. 9: 1245. https://doi.org/10.3390/biology12091245
APA StyleKim, K., Joo, G.-J., Jeong, K.-S., Gim, J.-S., Lee, Y., Hong, D., & Jo, H. (2023). Molecular Diet Analysis of Asian Clams for Supplementary Biodiversity Monitoring: A Case Study of Nakdong River Estuary. Biology, 12(9), 1245. https://doi.org/10.3390/biology12091245