
A New Chengjiang Worm Sheds Light on the Radiation and Disparity in Early Priapulida


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Preservation
2.2. Imaging
2.3. Element Mapping
2.4. Measurement
2.5. Phylogenetic Analysis
2.6. Terminology
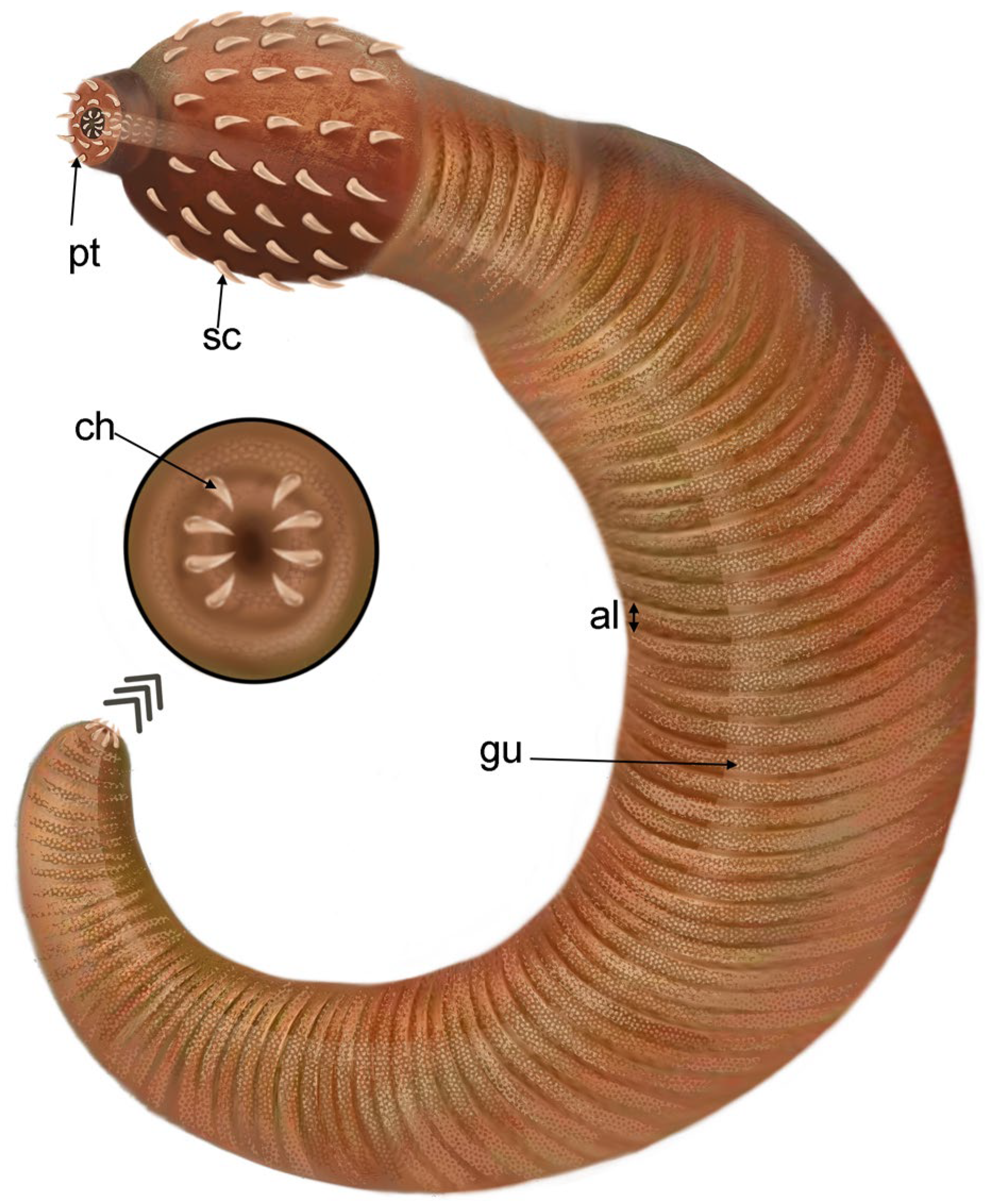
3. Results
Systematic Paleontology
4. Discussion
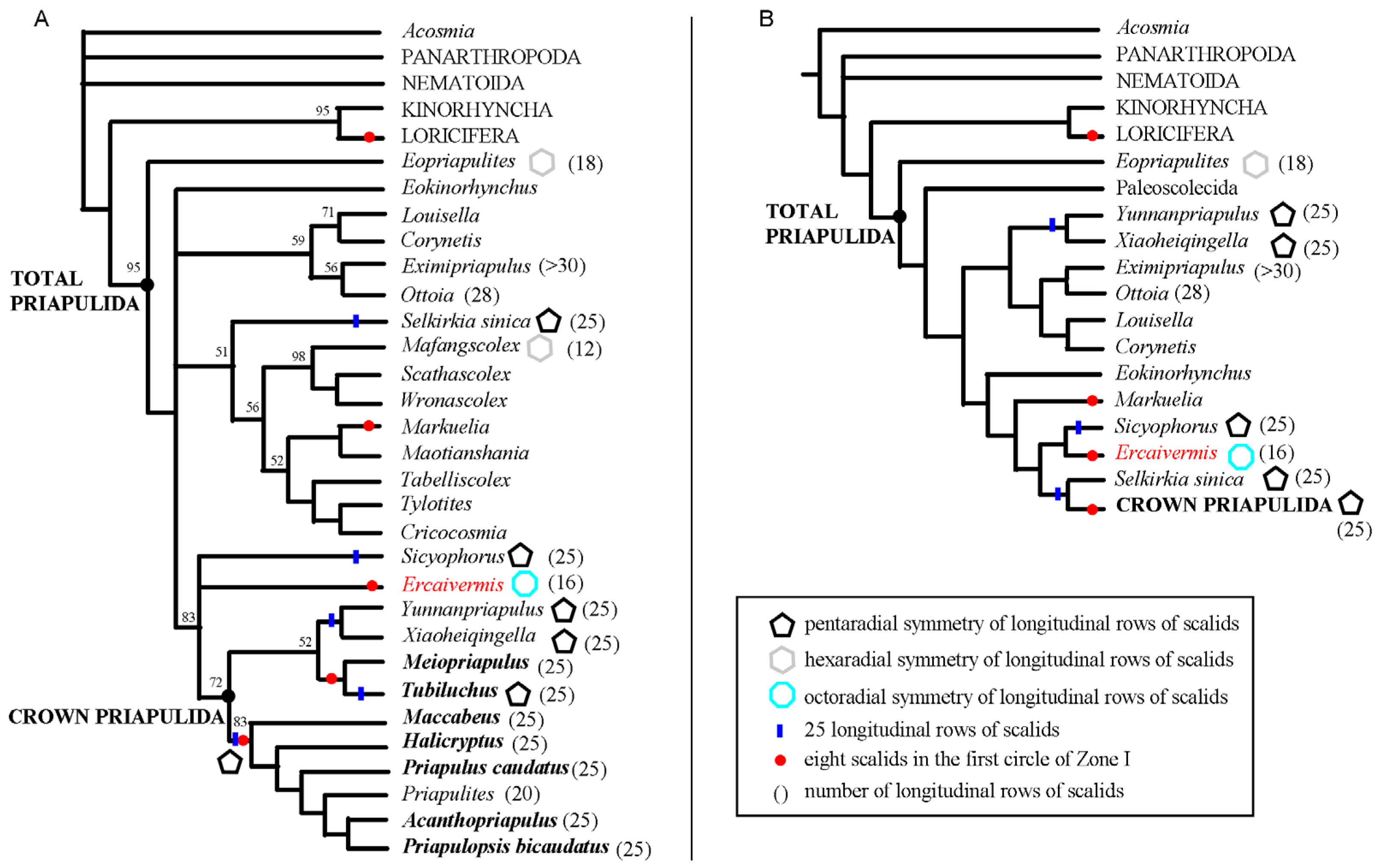
4.1. Phylogenetic Position of Ercaivermis among Priapulids
4.2. Eight Scalids around the First Circles: Morphological and Evolutionary Significance
4.3. How Can We Recognize a Crown-Group Priapulida?
4.4. Symmetry in Fossil Priapulids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
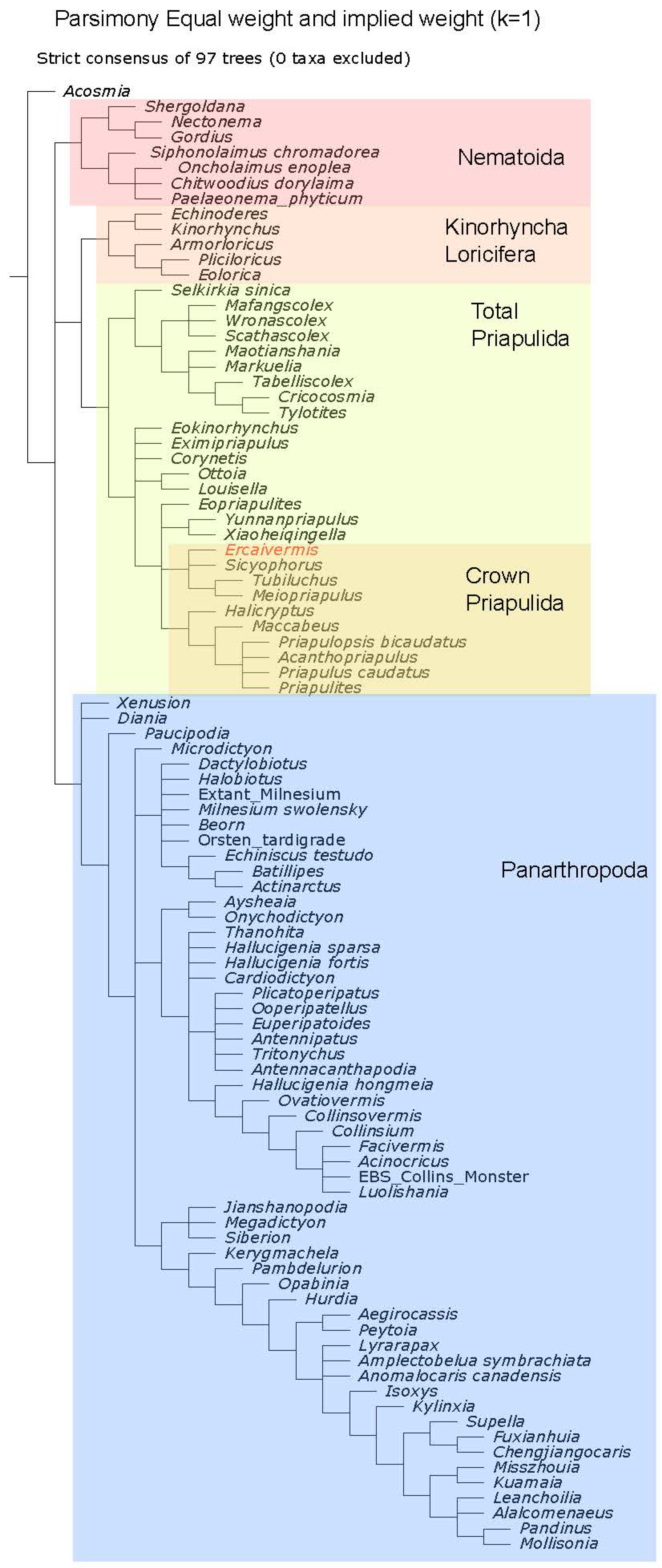
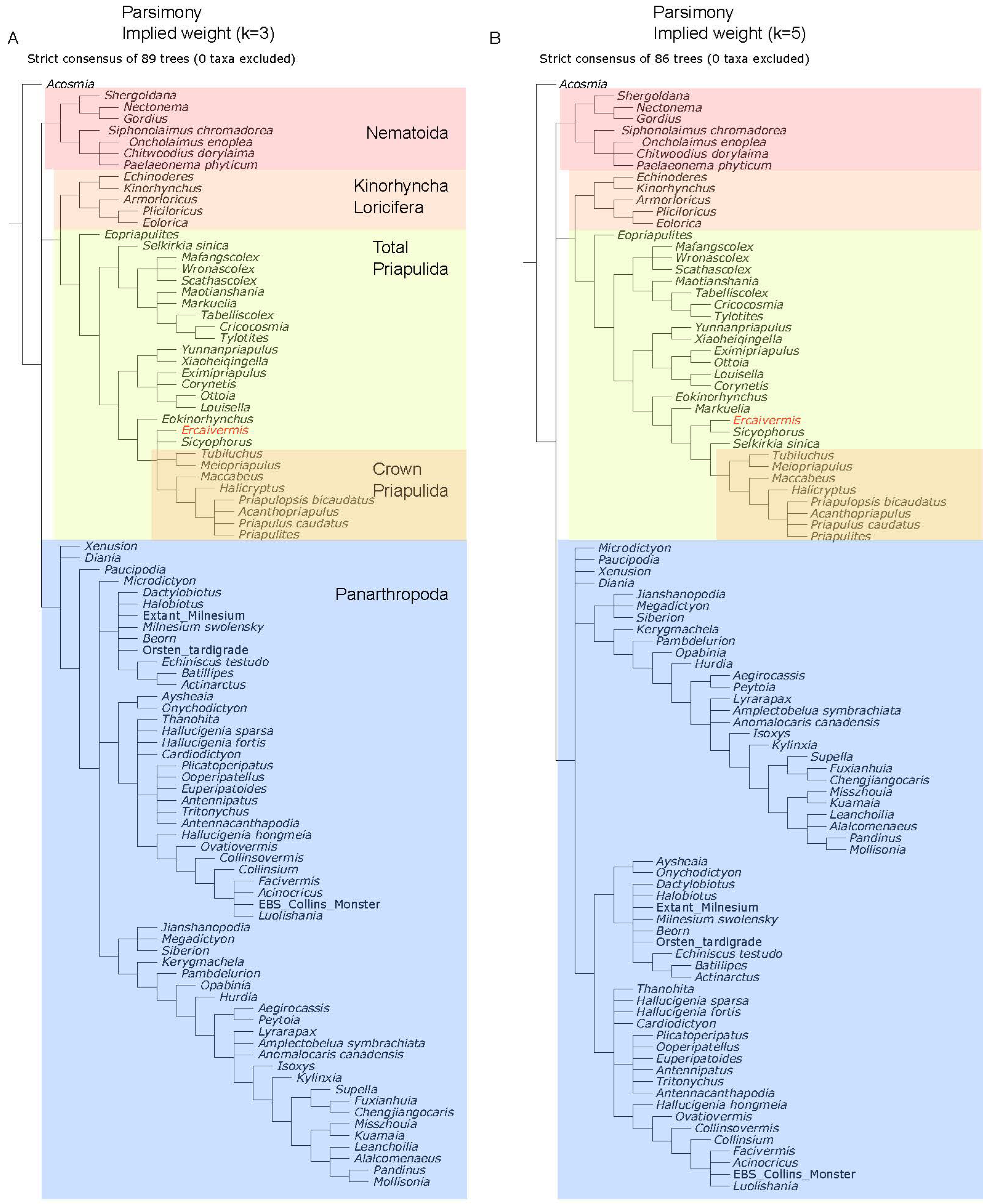

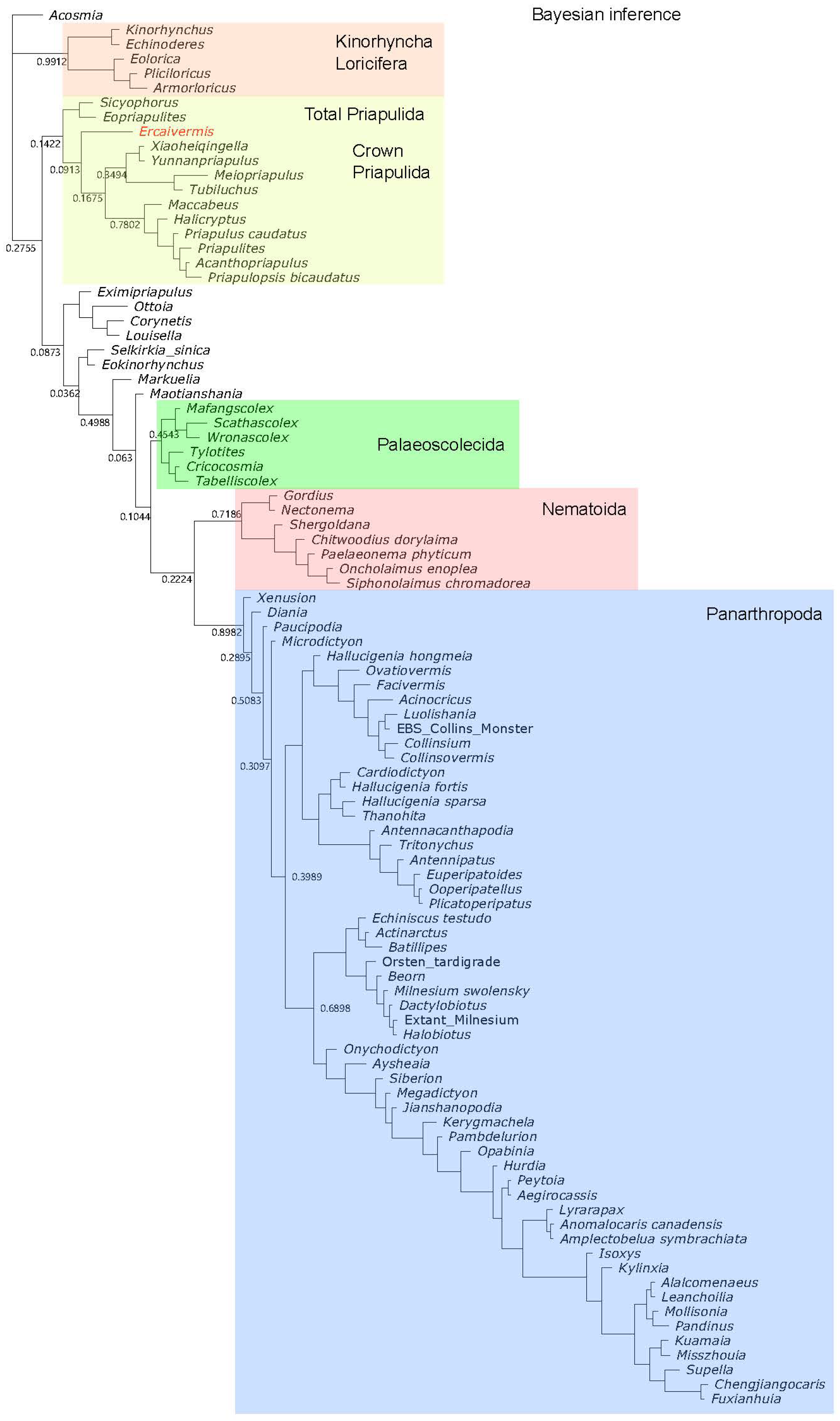
References
- Nielsen, C. Animal Evolution: Interrelationships of the Living Phyla, 3rd ed.; Oxford University Press: Oxford, UK, 2012; pp. 238–310. [Google Scholar]
- Brusca, R.C.; Moore, W.; Shuster, S.M. Invertebrates, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2016; pp. 639–910. [Google Scholar]
- Dong, X.P.; Donoghue, P.C.; Cunningham, J.A.; Liu, J.B.; Cheng, H. The anatomy, affinity, and phylogenetic significance of Markuelia. Evol. Dev. 2005, 7, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Harvey, T.H.; Dong, X.; Donoghue, P.C. Are palaeoscolecids ancestral ecdysozoans? Evol. Dev. 2010, 12, 177–200. [Google Scholar] [CrossRef]
- Wills, M.A.; Gerber, S.; Ruta, M.; Hughes, M. The disparity of priapulid, archaeopriapulid and palaeoscolecid worms in the light of new data. J. Evol. Biol. 2012, 25, 2056–2076. [Google Scholar] [CrossRef] [PubMed]
- Conway Morris, S.; Robison, R.A. Middle Cambrian priapulids and other soft-bodied fossils from Utah and Spain. Univ. Kans. Paleontol. Contrib. 1986, 117, 1–22. [Google Scholar]
- García-Bellido, D.C.; Paterson, J.R.; Edgecombe, G.D. Cambrian palaeoscolecids (Cycloneuralia) from Gondwana and reappraisal of species assigned to Palaeoscolex. Gondwana Res. 2013, 24, 780–795. [Google Scholar] [CrossRef]
- Conway Morris, S. Fossil priapulid worms. Spec. Pap. Palaeontol. 1977, 20, 1–95. [Google Scholar]
- Hou, X.G.; Siveter, D.J.; Siveter, D.J.; Aldridge, R.J.; Cong, P.Y.; Gabbott, S.E.; Ma, X.Y.; Purnell, M.A.; Williams, M. The Cambrian Fossils of Chengjiang, China: The Flowering of Early Animal Life, 2nd ed.; Wiley Blackwell: Hoboken, NJ, USA, 2017; pp. 98–270. [Google Scholar]
- Han, J. Introverta in Chengjiang Fauna; Northwest University: Xi’an, China, 2002; (In Chinese with English Abstract). [Google Scholar]
- Huang, D. Early Cambrian Worms from SW China: Morphology, Systematics, Lifestyles and Evolutionary Significance; Université Claude Bernard Lyon: Lyon, France, 2005. [Google Scholar]
- Peel, J.S. A corset-like fossil from the Cambrian Sirius Passet Lagerstätte of North Greenland and its implications for cycloneuralian evolution. J. Paleontol. 2010, 84, 332–340. [Google Scholar] [CrossRef]
- Harvey, T.H.P.; Butterfield, N.J. Exceptionally preserved Cambrian loriciferans and the early animal invasion of the meiobenthos. Nat. Ecol. Evol. 2017, 1, 22. [Google Scholar] [CrossRef]
- Zhang, H.; Xiao, S.; Liu, Y.; Yuan, X.; Wan, B.; Muscente, A.D.; Shao, T.; Gong, H.; Cao, G. Armored kinorhynch-like scalidophoran animals from the early Cambrian. Sci. Rep. 2015, 5, 16521. [Google Scholar] [CrossRef]
- Shao, T.Q.; Wang, Q.; Liu, Y.H.; Qin, J.C.; Zhang, Y.N.; Liu, M.J.; Shao, Y.; Zhao, J.Y.; Zhang, H.Q. A new scalidophoran animal from the Cambrian Fortunian Stage of South China and its implications for the origin and early evolution of Kinorhyncha. Precambrian Res. 2020, 349, 105616. [Google Scholar] [CrossRef]
- Maas, A.; Waloszek, D.; Haug, J.T.; Müller, K.J. A possible larval roundworm from the Cambrian ‘Orsten’ and its bearing on the phylogeny of Cycloneuralia. Mem. Assoc. Australas. Palaeontol. 2007, 34, 499–519. [Google Scholar]
- Huang, D.; Vannier, J.; Chen, J. Recent Priapulidae and their Early Cambrian ancestors: Comparisons and evolutionary significance. Geobios 2004, 37, 217–228. [Google Scholar] [CrossRef]
- Han, J.; Shu, D.; Zhang, Z.; Liu, J. The earliest-known ancestors of Recent Priapulomorpha from the Early Cambrian Chengjiang Lagerstätte. Chin. Sci. Bull. 2004, 49, 1860. [Google Scholar] [CrossRef]
- Han, J.; Hu, S. Response to “Discussion on the systematic position of the early Cambrian priapulomorph worms”. Chin. Sci. Bull. 2006, 51, 250–256. [Google Scholar] [CrossRef]
- Budd, G.E.; Jensen, S. A critical reappraisal of the fossil record of the bilaterian phyla. Bilogical Rev. 2000, 75, 253–295. [Google Scholar] [CrossRef]
- Wang, D.; Vannier, J.; Aria, C.; Sun, J.; Han, J. Tube-dwelling in early animals exemplified by Cambrian scalidophoran worms. BMC Biol. 2021, 19, 243. [Google Scholar] [CrossRef]
- Ma, X.Y.; Aldridge, R.J.; Siveter, D.J.; Siveter, D.J.; Hou, X.G.; Edgecombe, G.D. A new exceptionally preserved Cambrian priapulid from the Chengjiang Lagerstätte. J. Paleontol. 2014, 88, 371–384. [Google Scholar] [CrossRef]
- Schmidt-Rhaesa, A. Priapulida. In Gastrotricha, Cycloneuralia and Gnathifera; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2013; Volume 1, pp. 147–180. [Google Scholar]
- Adrianov, A.V.; Malakhov, V.V. Symmetry of priapulids (Priapulida). 2. Symmetry of larva. J. Morphol. 2001, 247, 111–121. [Google Scholar] [CrossRef]
- Adrianov, A.V.; Malakhov, V.V. Symmetry of priapulids (Priapulida). 1. Symmetry of adults. J. Morphol. 2001, 247, 99–110. [Google Scholar] [CrossRef]
- Lemburg, C. Ultrastructure of the introvert and associated structures of the larvae of Halicryptus spinulosus (Priapulida). Zoomorphology 1995, 115, 11–29. [Google Scholar] [CrossRef]
- Maas, A. Gastrotricha, Cycloneuralia and Gnathifera: The fossil record. In Gastrotricha, Cycloneuralia and Gnathifera; Schmidt-Rhaesa, A., Ed.; Nematomorpha; De Gruyter: Berlin, Germany, 2013; Volume 1, pp. 29–123. [Google Scholar]
- Cheng, J.L. Restudy of the Gourd-Shaped Priapulids from the Cambrian Chengjiang Biota and Their Phylogeny; Yunnan University: Kunming, China, 2019. [Google Scholar]
- Liu, Y.H.; Xiao, S.H.; Shao, T.Q.; Broce, J.; Zhang, H.Q. The oldest known priapulid-like scalidophoran animal and its implications for the early evolution of cycloneuralians and ecdysozoans. Evol. Dev. 2014, 16, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Edgecombe, G.D.; Shi, X.; Hou, X.; Ma, X. Ancestral morphology of Ecdysozoa constrained by an early Cambrian stem group ecdysozoan. BMC Evol. Biol. 2020, 20, 156. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Howard, R.J.; Edgecombe, G.D.; Hou, X.; Ma, X. Tabelliscolex (Cricocosmiidae: Palaeoscolecidomorpha) from the early Cambrian Chengjiang Biota and the evolution of seriation in Ecdysozoa. J. Geol. Soc. 2021, 179, jgs2021-060. [Google Scholar] [CrossRef]
- Bang-Berthelsen, I.H.; Schmidt-Rhaease, A.; Møbjerg, K. Loricifera. In Handbook of Zoology-Gastrotricha, Cycloneuralia and Gnathifera; Schmidt-Rhaesa, A., Ed.; Nematomorpha; De Gruyter: Berlin, Germany, 2013; Volume 1, pp. 349–370. [Google Scholar]
- Sun, W.G.; Hou, X.G. Early Cambrian worms from Chengjiang, Yunnan, China: Maotianshania gen. nov. Acta Palaeontol. Sin. 1987, 26, 299–307. [Google Scholar]
- Goloboff, P.A. Analyzing large data sets in reasonable times solutions for composite optima. Cladistics 1999, 24, 774–786. [Google Scholar] [CrossRef]
- Nixon, K.C. The Parsimony Ratchet, a New Method for Rapid Parsimony Analysis. Cladistics 1999, 15, 407–414. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Carpenter, J.M.; Arias, J.S.; Esquivel, D.R.M. Weighting against homoplasy improves phylogenetic analysis of morphological data sets. Cladistics 2008, 24, 758–773. [Google Scholar] [CrossRef]
- Lewis, P.O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 2001, 50, 913–925. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.; Xie, D.; Baele, G.; Suchard, M. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences (with discussion). Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Smith, M.R.; Dhungana, A. Discussion on ‘Tabelliscolex (Cricocosmiidae: Palaeoscolecidomorpha) from the early Cambrian Chengjiang Biota and the evolution of seriation in Ecdysozoa’ by Shi et al. 2021 (JGS, jgs2021-060). J. Geol. Soc. 2022, 179, jgs2021-111. [Google Scholar] [CrossRef]
- Neuhaus, B. Kinorhyncha (=Echinodera). In Handbook of Zoology-Gastrotricha, Cycloneuralia and Gnathifera; Schmidt-Rhaesa, A., Ed.; Nematomorpha; De Gruyter: Berlin, Germany, 2013; Volume 1, pp. 181–323. [Google Scholar]
- Rothe, B.H.; Schmidt-Rhaesa, A. Structure of the nervous system in Tubiluchus troglodytes (Priapulida). Invertebr. Biol. 2010, 129, 39–58. [Google Scholar] [CrossRef]
- Herranz, M.; Pardos, F.; Boyle, M.J. Comparative morphology of serotonergic-like immunoreactive elements in the central nervous system of kinorhynchs (Kinorhyncha, Cyclorhagida). J. Morphol. 2013, 274, 258–274. [Google Scholar] [CrossRef]
- Neuhaus, B. Ultrastructure of alimentary canal and body cavity, ground pattern, and phylogenetic relationships of the Kinorhyncha. Microfanua Mar. 1994, 9, 61–156. [Google Scholar]
- Vannier, J. Gut contents as direct indicators for trophic relationships in the Cambrian marine ecosystem. PLoS ONE 2012, 7, e52200. [Google Scholar] [CrossRef]
- Wernström, J.V.; Slater, B.J.; Sørensen, M.V.; Crampton, D.; Altenburger, A. Geometric morphometrics of macro- and meiofaunal priapulid pharyngeal teeth provides a proxy for studying Cambrian “tooth taxa”. Zoomorphology 2023. [Google Scholar] [CrossRef]
- Huang, D.; Chen, J.; Zhu, M.; Zhao, F. The burrow dwelling behavior and locomotion of palaeoscolecidian worms: New fossil evidence from the Cambrian Chengjiang fauna. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 398, 154–164. [Google Scholar] [CrossRef]
- Zhang, X.-G.; Hou, X.-G.; Bergström, J.A.N. Early Cambrian priapulid worms buried with their lined burrows. Geol. Mag. 2006, 143, 743–748. [Google Scholar] [CrossRef]
- Vannier, J.; Calandra, I.; Gaillard, C.; Żylińska, A. Priapulid worms: Pioneer horizontal burrowers at the Precambrian-Cambrian boundary. Geology 2010, 38, 711–714. [Google Scholar] [CrossRef]
- Kesidis, G.; Slater, B.J.; Jensen, S.; Budd, G.E. Caught in the act: Priapulid burrowers in early Cambrian substrates. Proc. R. Soc. B 2019, 286, 20182505. [Google Scholar] [CrossRef] [PubMed]
- Wennberg, S.A.; Janssen, R.; Budd, G.E. Hatching and earliest larval stages of the priapulid worm Priapulus caudatus. Invertebr. Biol. 2009, 128, 157–171. [Google Scholar] [CrossRef]
- Sørensen, M.V.; Rho, H.S.; Min, W.-G.; Kim, D. A new recording of the rare priapulid Meiopriapulus fijiensis, with comparative notes on juvenile and adult morphology. Zool. Anz.—J. Comp. Zool. 2012, 251, 364–371. [Google Scholar] [CrossRef]
- Storch, V.; Higgins, R.P. Scanning and transmission electron microscopic observations on the larva of Halicryptus spinulosus (priapulida). J. Morphol. 1991, 210, 175–194. [Google Scholar] [CrossRef]
- Higgins, R.P.; Stroch, V. Evidence for direct development in Meiopriapulus fijiensis (Priapulida). Trans. Am. Microsc. Soc. 1991, 110, 37–46. [Google Scholar] [CrossRef]
- Por, F.D.; Bromley, H.J. Morphology and anatomy of Maccabeus tentaculatus (Priapulida: Seticoronaria). J. Zool. 1974, 173, 173–197. [Google Scholar] [CrossRef]
- Schmidt-Rhaesa, A. The Evolution of Organ Systems; Oxford University Press: Oxford, UK, 2007; pp. 54–73. [Google Scholar]
- Schram, F.R. Pseudocoelomates and a Nemertine from the Illinois Pennsylvanian. J. Paleontol. 1973, 47, 985–989. [Google Scholar]
- Liu, Y.H.; Qin, J.C.; Wang, Q.; Maas, A.; Duan, B.C.; Zhang, Y.N.; Zhang, H.; Shao, T.Q.; Zhang, H.Q.; Zhang, X.G. New armoured scalidophorans (Ecdysozoa, Cycloneuralia) from the Cambrian Fortunian Zhangjiagou Lagerstätte, South China. Pap. Palaeontol. 2018, 5, 241–260. [Google Scholar] [CrossRef]
- Yang, J.; Smith, M.R.; Zhang, X.-G.; Yang, X.-Y. Introvert and pharynx of Mafangscolex, a Cambrian palaeoscolecid. Geol. Mag. 2020, 157, 2044–2050. [Google Scholar] [CrossRef]
- Budd, G.E.; Mann, R.P. The dynamics of stem and crown groups. Sci. Adv. 2020, 6, eaaz1626. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Kubota, S.; Li, G.; Ou, Q.; Wang, X.; Yao, X.; Shu, D.; Li, Y.; Uesugi, K.; Hoshino, M.; et al. Divergent evolution of medusozoan symmetric patterns: Evidence from the microanatomy of Cambrian tetramerous cubozoans from South China. Gondwana Res. 2016, 31, 150–163. [Google Scholar] [CrossRef]
- McMenamin, M.A.S. Dynamic Paleontology: Using Quantification and Other Tools to Decipher the History of Life; Springer: Cham, Switzerland, 2016. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Vannier, J.; Sun, J.; Yu, C.; Han, J. A New Chengjiang Worm Sheds Light on the Radiation and Disparity in Early Priapulida. Biology 2023, 12, 1242. https://doi.org/10.3390/biology12091242
Wang D, Vannier J, Sun J, Yu C, Han J. A New Chengjiang Worm Sheds Light on the Radiation and Disparity in Early Priapulida. Biology. 2023; 12(9):1242. https://doi.org/10.3390/biology12091242
Chicago/Turabian StyleWang, Deng, Jean Vannier, Jie Sun, Chiyang Yu, and Jian Han. 2023. "A New Chengjiang Worm Sheds Light on the Radiation and Disparity in Early Priapulida" Biology 12, no. 9: 1242. https://doi.org/10.3390/biology12091242
APA StyleWang, D., Vannier, J., Sun, J., Yu, C., & Han, J. (2023). A New Chengjiang Worm Sheds Light on the Radiation and Disparity in Early Priapulida. Biology, 12(9), 1242. https://doi.org/10.3390/biology12091242