
Establishment of a Coilia nasus Spermatogonial Stem Cell Line Capable of Spermatogenesis In Vitro

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
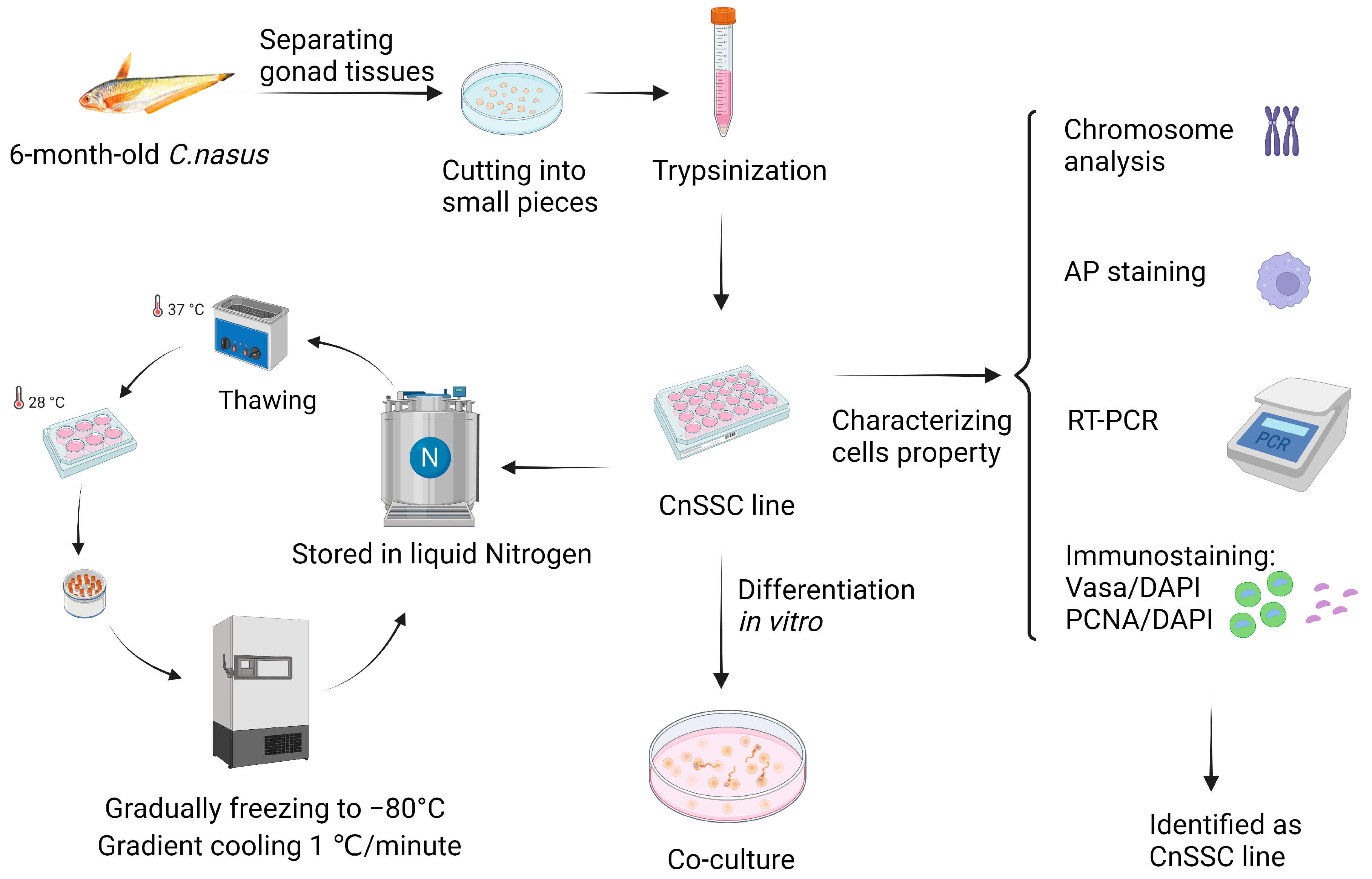
2.1. Fish, Primary Cell Culture, and Subculture
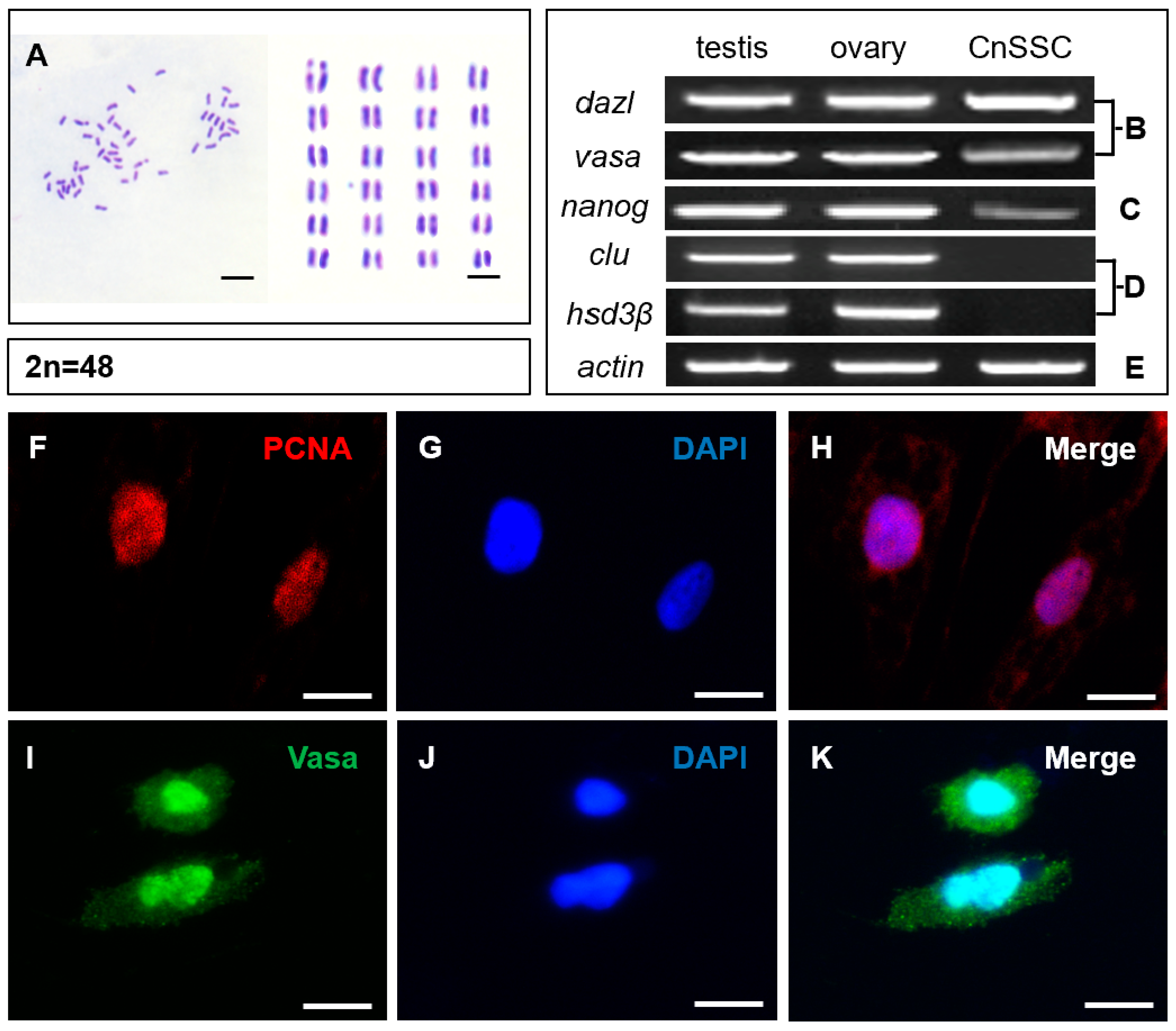
2.2. Chromosome Analysis
2.3. Gene Expression Analysis
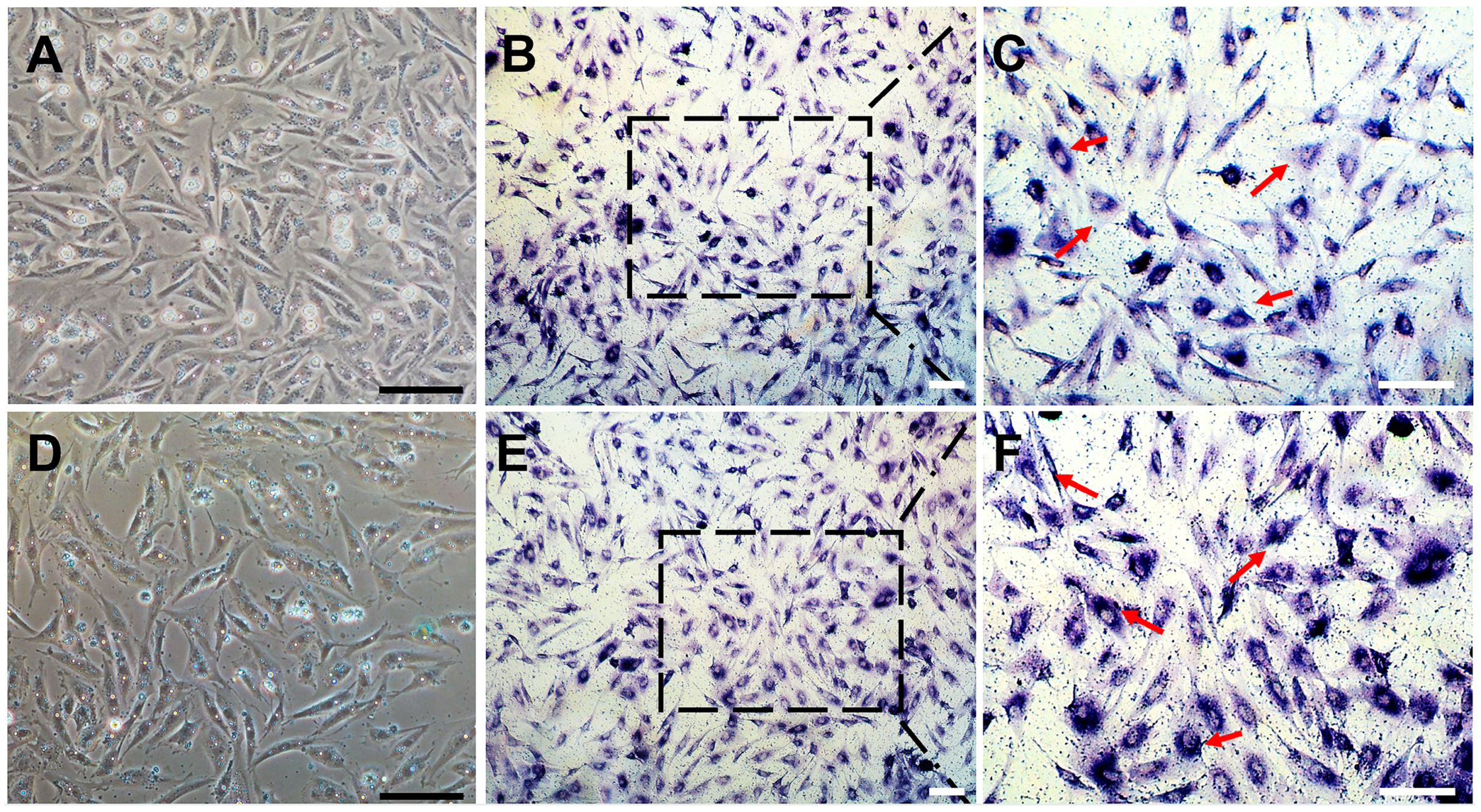
2.4. Cryopreservation, Thawing, and Alkaline Phosphatase (AP) Staining of CnSSC
2.5. Immunofluorescence Staining of CnSSC
2.6. Cell Transfection
2.7. Coculture of CnGSCs and CnSSCs
3. Results
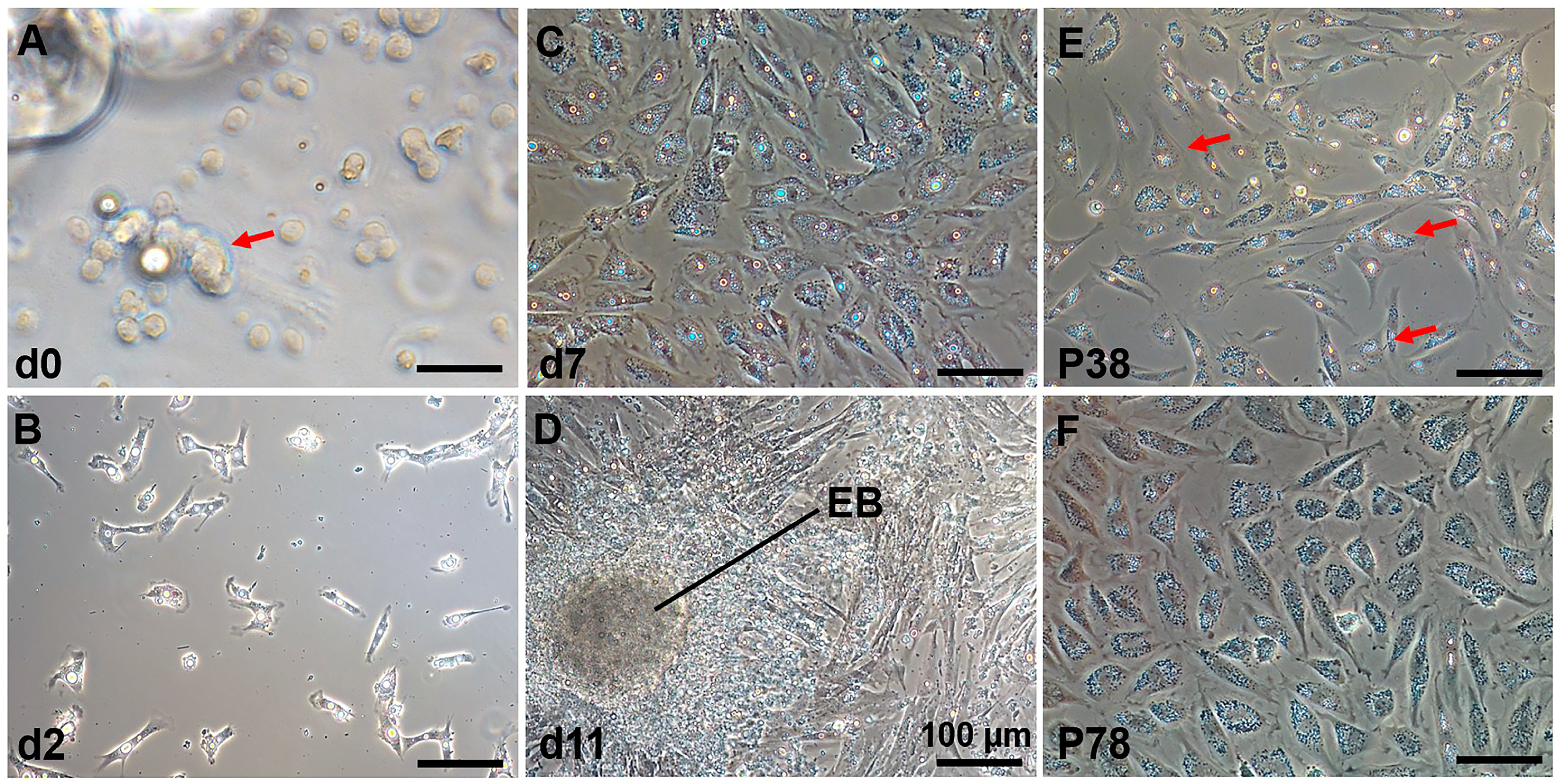
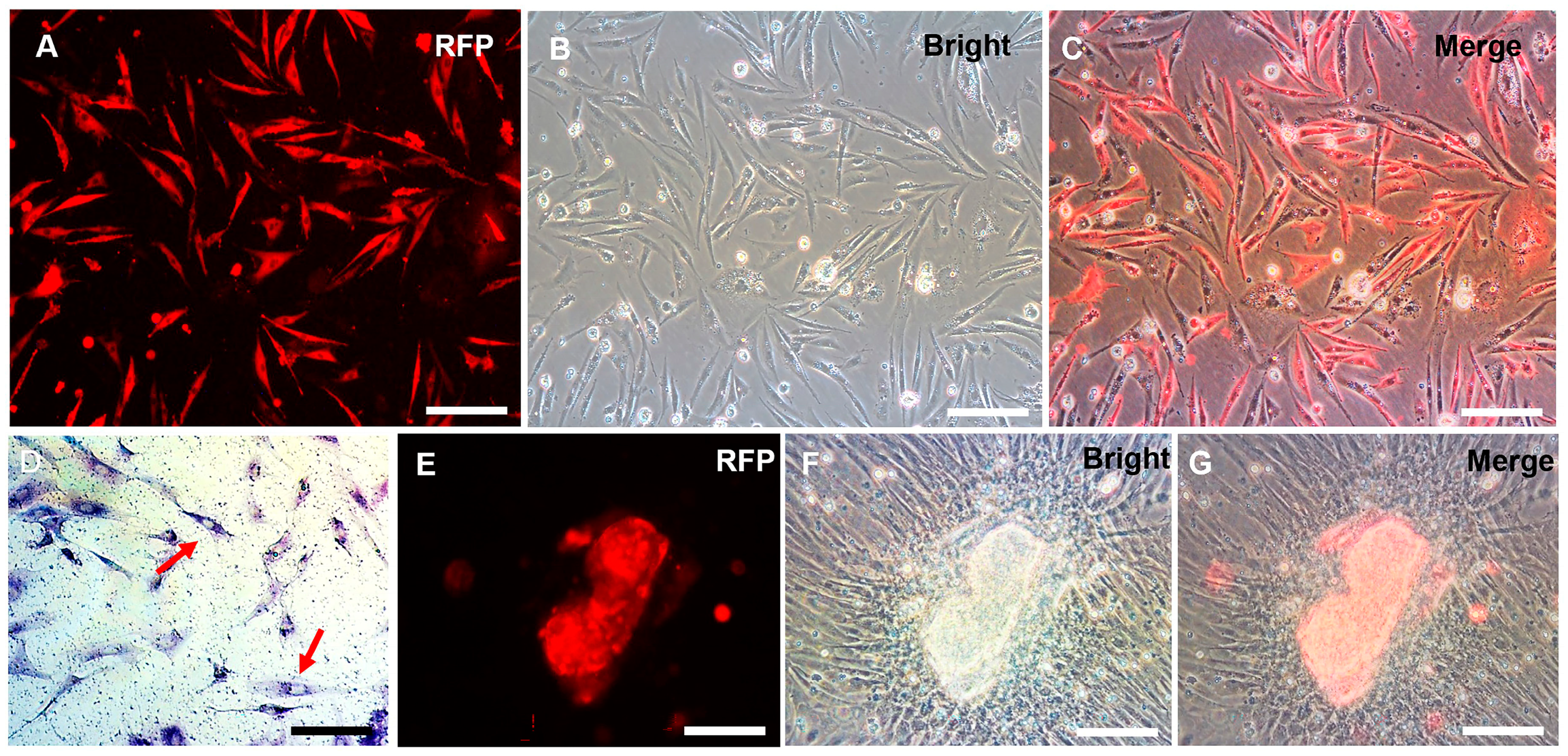
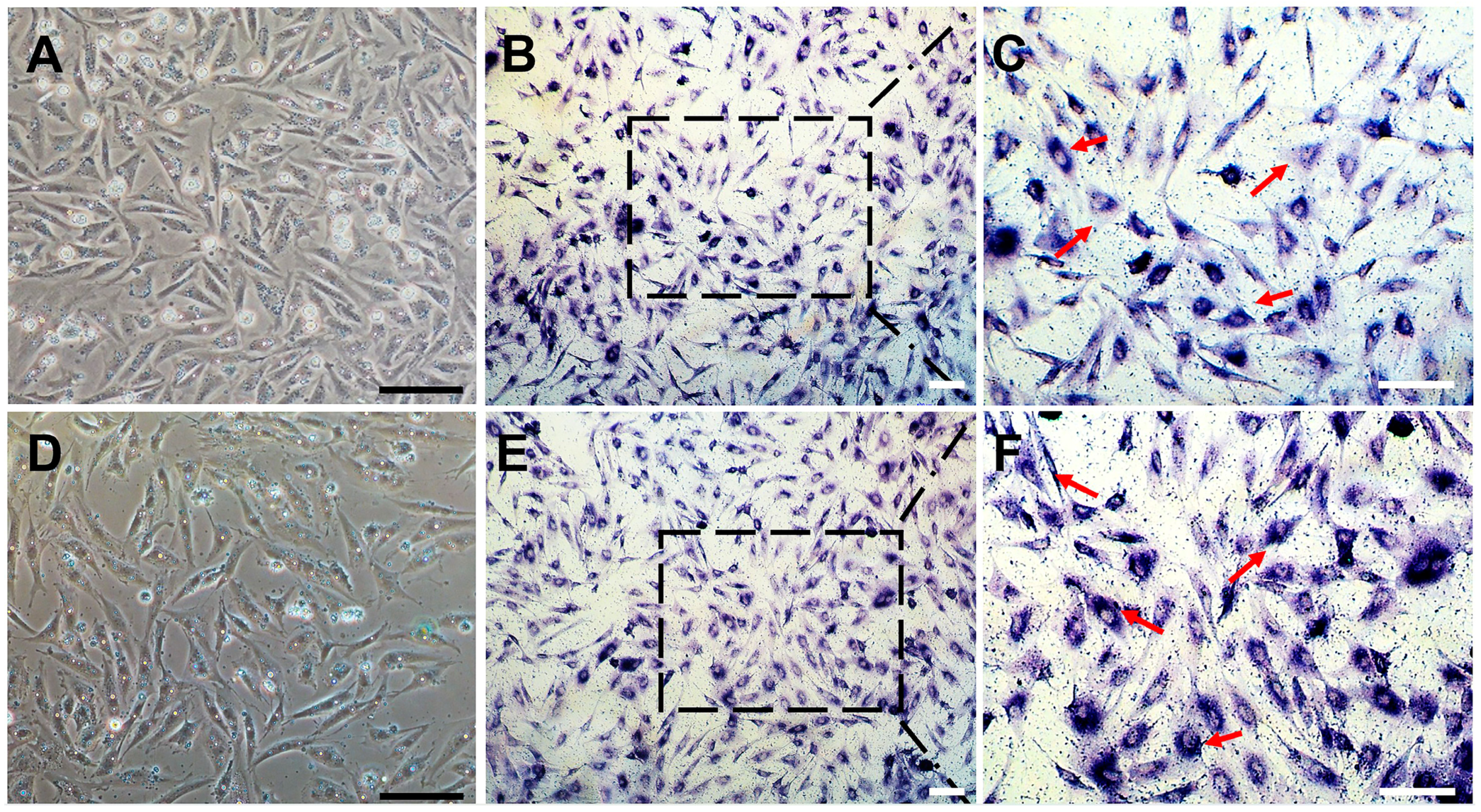
3.1. Establishment of a Spermatogonial Stem Cell Line
3.2. Chromosome Analysis and Characterization of Spermatogonial Stem Cell Properties
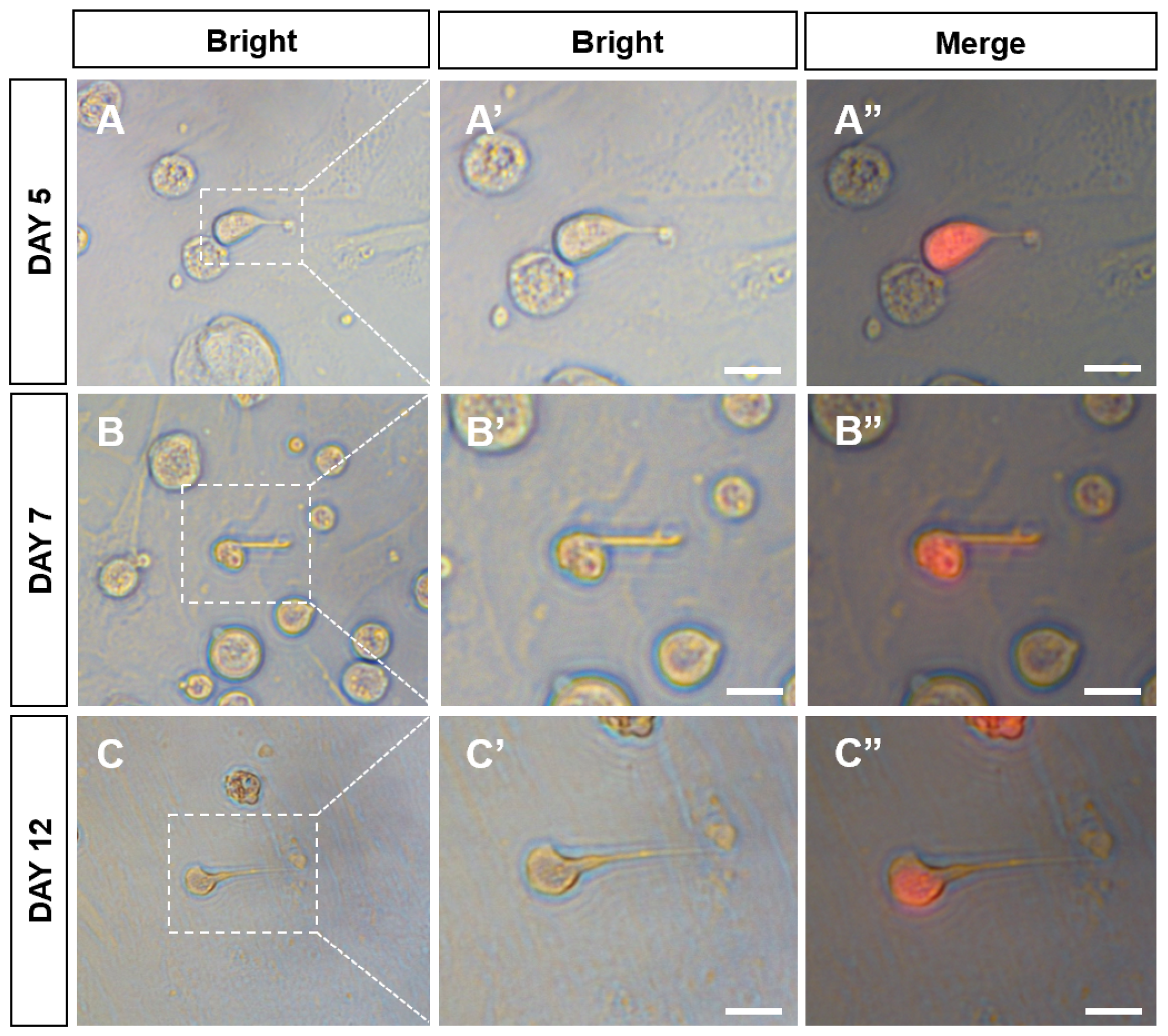
3.3. Sperm Production of CnSSCs In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kan, Y.; Zhong, Y.; Jawad, M.; Chen, X.; Liu, D.; Ren, M.; Xu, G.; Gui, L.; Li, M. Establishment of a Coilia nasus gonadal somatic cell line capable of sperm induction in vitro. Biology 2022, 11, 1049. [Google Scholar] [PubMed]
- Goswami, M.; Mishra, A. Bio-banking: An emerging approach for conservation of fish germplasm. Poult. Fish. Wildl. Sci. 2016, 4, 143. [Google Scholar] [CrossRef]
- Niu, J.; Wang, X.; Liu, P.; Liu, H.; Li, R.; Li, Z.; He, Y.; Qi, J. Effects of cryopreservation on sperm with cryodiluent in viviparous black rockfish (Sebastes schlegelii). Int. J. Mol. Sci. 2022, 23, 3392. [Google Scholar] [PubMed]
- Xu, Y.; Zhong, Z.; Zhang, Z.; Feng, Y.; Zhao, L.; Jiang, Y.; Wang, Y. Establishment and characterization of the gonadal cell lines derived from large yellow croaker (Larimichthys crocea) for gene expression studies. Aquaculture 2022, 546, 737300. [Google Scholar]
- Hong, Y.; Liu, T.; Zhao, H.; Xu, H.; Wang, W.; Liu, R.; Chen, T.; Deng, J.; Gui, J. Establishment of a normal Medaka fish spermatogonial cell line capable of sperm production in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 8011–8016. [Google Scholar]
- Kubota, H.; Brinster, R.L. Spermatogonial stem cells. Biol. Reprod. 2018, 99, 52–74. [Google Scholar] [PubMed]
- Nagano, M.C.; Yeh, J.R. The identity and fate decision control of spermatogonial stem cells: Where is the point of no return? Curr. Top. Dev. Biol. 2013, 102, 61–95. [Google Scholar] [PubMed]
- Valli, H.; Phillips, B.T.; Shetty, G.; Byrne, J.A.; Clark, A.T.; Meistrich, M.L.; Orwig, K.E. Germline stem cells: Toward the regeneration of spermatogenesis. Fertil. Steril. 2014, 101, 3–13. [Google Scholar]
- de Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar]
- Zhang, R.; Sun, J.; Zou, K. Advances in isolation methods for spermatogonial stem cells. Stem Cell Rev. Rep. 2016, 12, 15–25. [Google Scholar]
- Xie, X.; Nobrega, R.; Psenicka, M. Spermatogonial stem cells in fish: Characterization, isolation, enrichment, and recent advances of in vitro culture systems. Biomolecules 2020, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Ishikura, Y.; Ohta, H.; Sato, T.; Murase, Y.; Yabuta, Y.; Kojima, Y.; Yamashiro, C.; Nakamura, T.; Yamamoto, T.; Ogawa, T.; et al. In vitro reconstitution of the whole male germ-cell development from mouse pluripotent stem cells. Cell Stem Cell 2021, 28, 2167–2179.e9. [Google Scholar] [PubMed]
- Xu, W.; Li, H.; Peng, L.; Pu, L.; Xiang, S.; Li, Y.; Tao, L.; Liu, W.; Liu, J.; Xiao, Y. Fish pluripotent stem-like cell line induced by small-molecule compounds from caudal fin and its developmental potentiality. Front. Cell Dev. Biol. 2022, 9, 3965. [Google Scholar]
- Chen, X.; Kan, Y.; Zhong, Y.; Jawad, M.; Wei, W.; Gu, K.; Gui, L.; Li, M. Generation of a normal long-term-cultured Chinese hook snout carp spermatogonial stem cell line capable of sperm production in vitro. Biology 2022, 11, 1069. [Google Scholar]
- Wang, M.; Xu, G.; Tang, Y.; Xu, P. Transcriptome analysis of the brain provides insights into the regulatory mechanism for Coilia nasus migration. BMC Genom. 2020, 21, 410. [Google Scholar]
- Huang, Y.F.; Rypel, A.L.; Murphy, B.R.; Xie, S.G. Declined fitness in larvae born from long-distance migrants of anadromous Coilia nasus in the lower reaches of the Yangtze River, China. Zool. Res. 2022, 43, 404–408. [Google Scholar] [CrossRef]
- Fang, D.A.; Yang, X.J.; Feng, X.; Zhou, Y.F.; Xu, D.P.; Zhang, M.Y.; Liu, K. FoxL2 combined with Cyp19a1a regulate the spawning upstream migration in Coilia nasus. Gene 2019, 710, 307–315. [Google Scholar] [CrossRef]
- Hong, Y.; Schartl, M. Isolation and differentiation of Medaka embryonic stem cells. Methods Mol. Biol. 2006, 329, 3–16. [Google Scholar]
- Yi, M.; Hong, N.; Hong, Y. Derivation and characterization of haploid embryonic stem cell cultures in Medaka fish. Nat. Protoc. 2010, 5, 1418–1430. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Hong, N.; Hong, Y. Generation of Medaka fish haploid embryonic stem cells. Science 2009, 326, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, P.; Xia, J.; Guo, J.; Shi, Y.; Zhong, Y.; Li, M. Evolutionarily conserved boule and dazl identify germ cells of Coilia nasus. Aquac. Fish. 2021, 8, 244–251. [Google Scholar] [CrossRef]
- Hong, Y.; Winkler, C.; Schartl, M. Pluripotency and differentiation of embryonic stem cell lines from the Medaka fish (Oryzias latipes). Mech. Dev. 1996, 60, 33–44. [Google Scholar] [CrossRef]
- Zhao, H.; Hong, N.; Lu, W.; Zeng, H.; Song, J.; Hong, Y. Fusion gene vectors allowing for simultaneous drug selection, cell labeling, and reporter assay in vitro and in vivo. Anal. Chem. 2012, 84, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, L.; Fan, Z.; Hong, Y.; Zhao, Y.; Zhou, L.; Wang, D. Identification, prokaryote expression of Medaka gdnfa/b and their biological activity in a spermatogonial cell line. Stem Cells Dev. 2017, 26, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Tao, Y.; Liu, M.; Wu, X.; Yang, Y.; Wang, T.; Meng, Z.; Xu, H.; Liu, X. Establishment of a spermatogonial stem cell line with potential of meiosis in a Hermaphroditic fish, Epinephelus coioides. Cells 2022, 11, 2868. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Zhuang, P.; Song, C.; Zhao, F.; Zhou, L.; Geng, Z.; Hao, C.; Zhang, T. Karyotype analysis of Coilia nasus. Mar. Fish. 2020, 42, 005. [Google Scholar]
- Jin, Y.H.; Davie, A.; Migaud, H. Expression pattern of nanos, piwil, dnd, vasa and pum genes during ontogenic development in Nile tilapia Oreochromis niloticus. Gene 2019, 688, 62–70. [Google Scholar] [CrossRef]
- Li, M.; Guan, G.; Hong, N.; Hong, Y. Multiple regulatory regions control the transcription of Medaka germ gene vasa. Biochimie 2013, 95, 850–857. [Google Scholar] [CrossRef]
- Yoshizaki, G.; Sakatani, S.; Tominaga, H.; Takeuchi, T. Cloning and characterization of a vasa-like gene in rainbow trout and its expression in the germ cell lineage. Mol. Reprod. Dev. 2000, 55, 364–371. [Google Scholar] [CrossRef]
- Sánchez-Sánchez, A.V.; Camp, E.; Mullor, J.L. Fishing pluripotency mechanisms in vivo. Int. J. Biol. Sci. 2011, 7, 410. [Google Scholar] [CrossRef]
- Sun, B.; Gui, L.; Liu, R.; Hong, Y.; Li, M. Medaka oct4 is essential for gastrulation, central nervous system development and angiogenesis. Gene 2020, 733, 144270. [Google Scholar] [CrossRef]
- Patra, S.K.; Vemulawada, C.; Soren, M.M.; Sundaray, J.K.; Panda, M.K.; Barman, H.K. Molecular characterization and expression patterns of Nanog gene validating its involvement in the embryonic development and maintenance of spermatogonial stem cells of farmed carp, Labeo rohita. J. Anim. Sci. Biotechnol. 2018, 9, 45. [Google Scholar] [CrossRef]
- Xu, H.; Zhu, X.; Li, W.; Tang, Z.; Zhao, Y.; Wu, X. Isolation and in vitro culture of ovarian stem cells in Chinese soft-shell turtle (Pelodiscus sinensis). J. Cell Biochem. 2018, 119, 7667–7677. [Google Scholar] [CrossRef]
- Riaz, M.A.S.A.; Borgers, M.; Konrad, L. Clusterin signals via ApoER2/VLDLR and induces meiosis of male germ cells. Am. J. Transl. Res. 2017, 9, 1266–1276. [Google Scholar]
- Nakamoto, M.; Fukasawa, M.; Tanaka, S.; Shimamori, K.; Suzuki, A.; Matsuda, M.; Kobayashi, T.; Nagahama, Y.; Shibata, N. Expression of 3beta-hydroxysteroid dehydrogenase (hsd3b), star and ad4bp/sf-1 during gonadal development in medaka (Oryzias latipes). Gen. Comp. Endocrinol. 2012, 176, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Gui, J.; Hong, Y. Differential expression of vasa RNA and protein during spermatogenesis and oogenesis in the gibel carp (Carassius auratus gibelio), a bisexually and gynogenetically reproducing vertebrate. Dev. Dyn. 2005, 233, 872–882. [Google Scholar] [CrossRef]
- Ye, D.; Liu, T.; Li, Y.; Wang, Y.; Hu, W.; Zhu, Z.; Sun, Y. Identification of fish spermatogenic cells through high-throughput immunofluorescence against testis with an antibody set. Front. Endocrinol. 2023, 14, 1044318. [Google Scholar] [CrossRef]
- Zhang, F.; Hao, Y.; Li, X.; Li, Y.; Ye, D.; Zhang, R.; Wang, X.; He, M.; Wang, H.; Zhu, Z.; et al. Surrogate production of genome-edited sperm from a different subfamily by spermatogonial stem cell transplantation. Sci. China Life Sci. 2022, 65, 969–987. [Google Scholar] [CrossRef] [PubMed]
- de Siqueira-Silva, D.H.; Saito, T.; Dos Santos-Silva, A.P.; da Silva Costa, R.; Psenicka, M.; Yasui, G.S. Biotechnology applied to fish reproduction: Tools for conservation. Fish. Physiol. Biochem. 2018, 44, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Tao, B.; Wang, Y.; Hu, W.; Sun, Y. Isolation and characterization of germline stem cells in protogynous hermaphroditic Monopterus albus. Int. J. Mol. Sci. 2022, 23, 5861. [Google Scholar] [CrossRef]
- de Rooij, D.G. The nature and dynamics of spermatogonial stem cells. Development 2017, 144, 3022–3030. [Google Scholar] [CrossRef]
- Sato, T.; Katagiri, K.; Yokonishi, T.; Kubota, Y.; Inoue, K.; Ogonuki, N.; Matoba, S.; Ogura, A.; Ogawa, T. In vitro production of fertile sperm from murine spermatogonial stem cell lines. Nat. Commun. 2011, 2, 472. [Google Scholar] [CrossRef] [PubMed]
- Bashiri, Z.; Amiri, I.; Gholipourmalekabadi, M.; Falak, R.; Asgari, H.; Maki, C.B.; Moghaddaszadeh, A.; Koruji, M. Artificial testis: A testicular tissue extracellular matrix as a potential bio-ink for 3D printing. Biomater. Sci. 2021, 9, 3465–3484. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.W.; Mo, C.Y.; Dong, M.D.; Jia, K.T.; Liu, W.; Yi, M.S. Production of functional sperm from in vitro-cultured premeiotic spermatogonia in a marine fish. Zool. Res. 2022, 43, 537–551. [Google Scholar] [CrossRef]
- Xie, H.; Wang, X.; Jin, M.; Li, L.; Zhu, J.; Kang, Y.; Chen, Z.; Sun, Y.; Zhao, C. Cilia regulate meiotic recombination in zebrafish. J. Mol. Cell Biol. 2022, 14, mjac049. [Google Scholar] [CrossRef] [PubMed]
- Rankin, S. Complex elaboration: Making sense of meiotic cohesin dynamics. FEBS J. 2015, 282, 2426–2443. [Google Scholar] [CrossRef]
- Yoon, S.W.; Lee, M.S.; Xaver, M.; Zhang, L.; Hong, S.G.; Kong, Y.J.; Cho, H.R.; Kleckner, N.; Kim, K.P. Meiotic prophase roles of Rec8 in crossover recombination and chromosome structure. Nucleic Acids Res. 2016, 44, 9296–9314. [Google Scholar] [CrossRef]
- Nayak, R.; Franek, R.; Sindelka, R.; Psenicka, M. Enhancement of zebrafish sperm production via a large body-sized surrogate with germ cell transplantation. Commun. Biol. 2023, 6, 412. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| Name | Forward Primer | Reverse Primer |
| dazl | ACCTGAGGGCAAAATGACACC | CGTGAGCTCCTCTCTTTCATGATGG |
| vasa | ACGCCATCTTCAATCAGTTCCAGACC | CTATTCCCATTCGTCGTCATCTCCGC |
| nanog | ATGGCGGACTGGAAAGTACCAGTAAG | CACAATCTGCAATGCACACAAACATTCAG |
| clu | TCTCTGCTCTGTGTCTTATC | AACTTCTTGTGGTCCTCTC |
| hsd3β | GTGGTGGTGGTAGCGAAGT | GCCTCCGACAGCATACAGT |
| dmc1 | TGTCACCAACCAGATGACGG | TTGGCATCCGTGATTCCTCC |
| rec8 | CCGAGTCTGCCTAAACCACG | CTTTCTCCTTAAGAGTGATG |
| β-actin | TTCAACAGCCCTGCCATGTAC | CCTCCAATCCAGACAGAGTATT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, K.; Zhang, Y.; Zhong, Y.; Kan, Y.; Jawad, M.; Gui, L.; Ren, M.; Xu, G.; Liu, D.; Li, M. Establishment of a Coilia nasus Spermatogonial Stem Cell Line Capable of Spermatogenesis In Vitro. Biology 2023, 12, 1175. https://doi.org/10.3390/biology12091175
Gu K, Zhang Y, Zhong Y, Kan Y, Jawad M, Gui L, Ren M, Xu G, Liu D, Li M. Establishment of a Coilia nasus Spermatogonial Stem Cell Line Capable of Spermatogenesis In Vitro. Biology. 2023; 12(9):1175. https://doi.org/10.3390/biology12091175
Chicago/Turabian StyleGu, Kaiyan, Ya Zhang, Ying Zhong, Yuting Kan, Muhammad Jawad, Lang Gui, Mingchun Ren, Gangchun Xu, Dong Liu, and Mingyou Li. 2023. "Establishment of a Coilia nasus Spermatogonial Stem Cell Line Capable of Spermatogenesis In Vitro" Biology 12, no. 9: 1175. https://doi.org/10.3390/biology12091175
APA StyleGu, K., Zhang, Y., Zhong, Y., Kan, Y., Jawad, M., Gui, L., Ren, M., Xu, G., Liu, D., & Li, M. (2023). Establishment of a Coilia nasus Spermatogonial Stem Cell Line Capable of Spermatogenesis In Vitro. Biology, 12(9), 1175. https://doi.org/10.3390/biology12091175