
The Effect of Month of Harvesting and Temperature–Humidity Index on the Number and Quality of Oocytes and In Vitro Embryo Production in Holstein Cows and Heifers

 , , , , , ,
, , , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
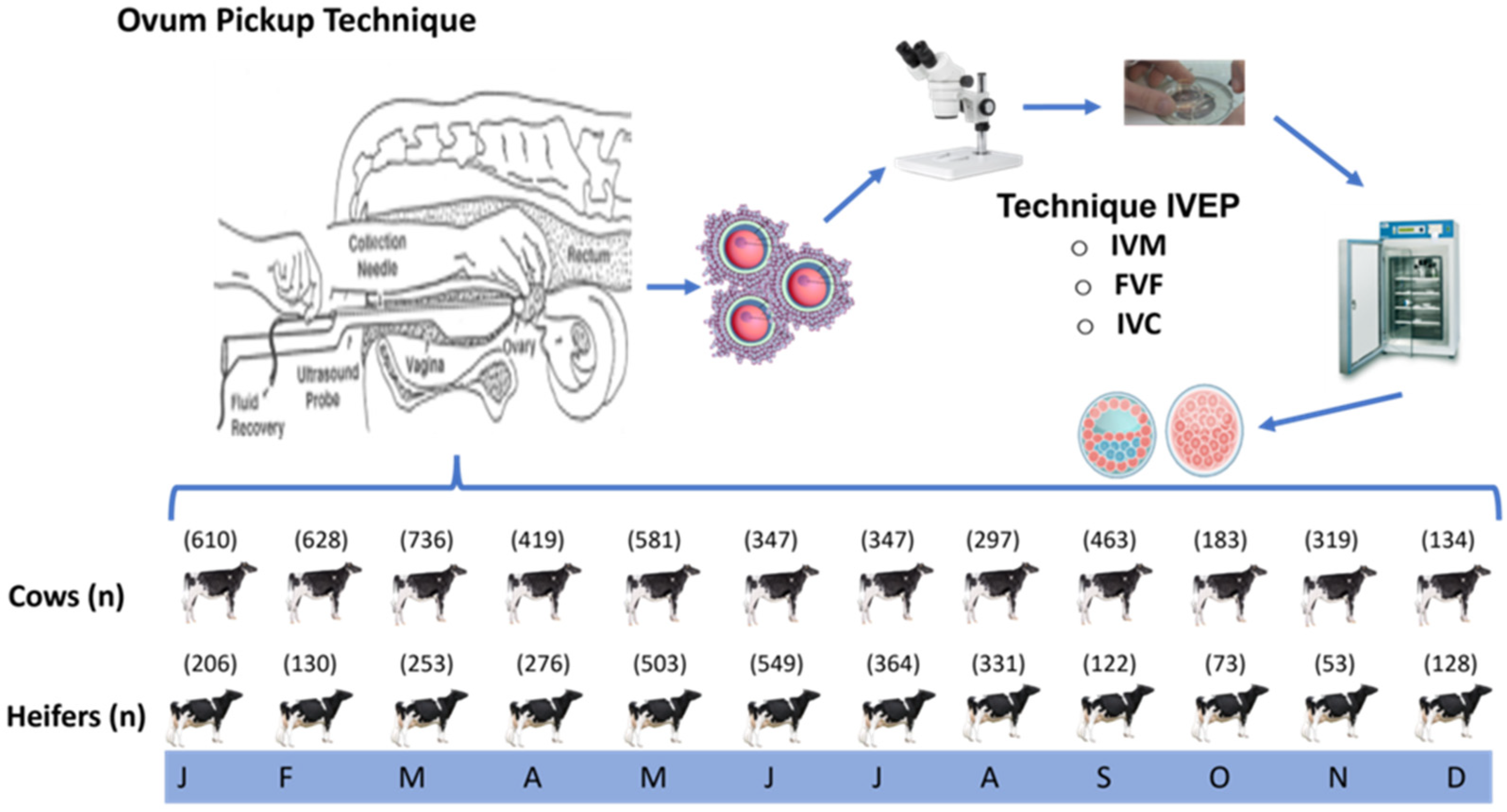
2. Materials and Methods
2.1. General
2.2. Location of the Study Area and Climatic Conditions
2.3. Meteorological Data and THI Calculation
2.4. Animals and Handling Conditions
2.5. Design of the Experiment
2.6. Statistical Analysis
3. Results
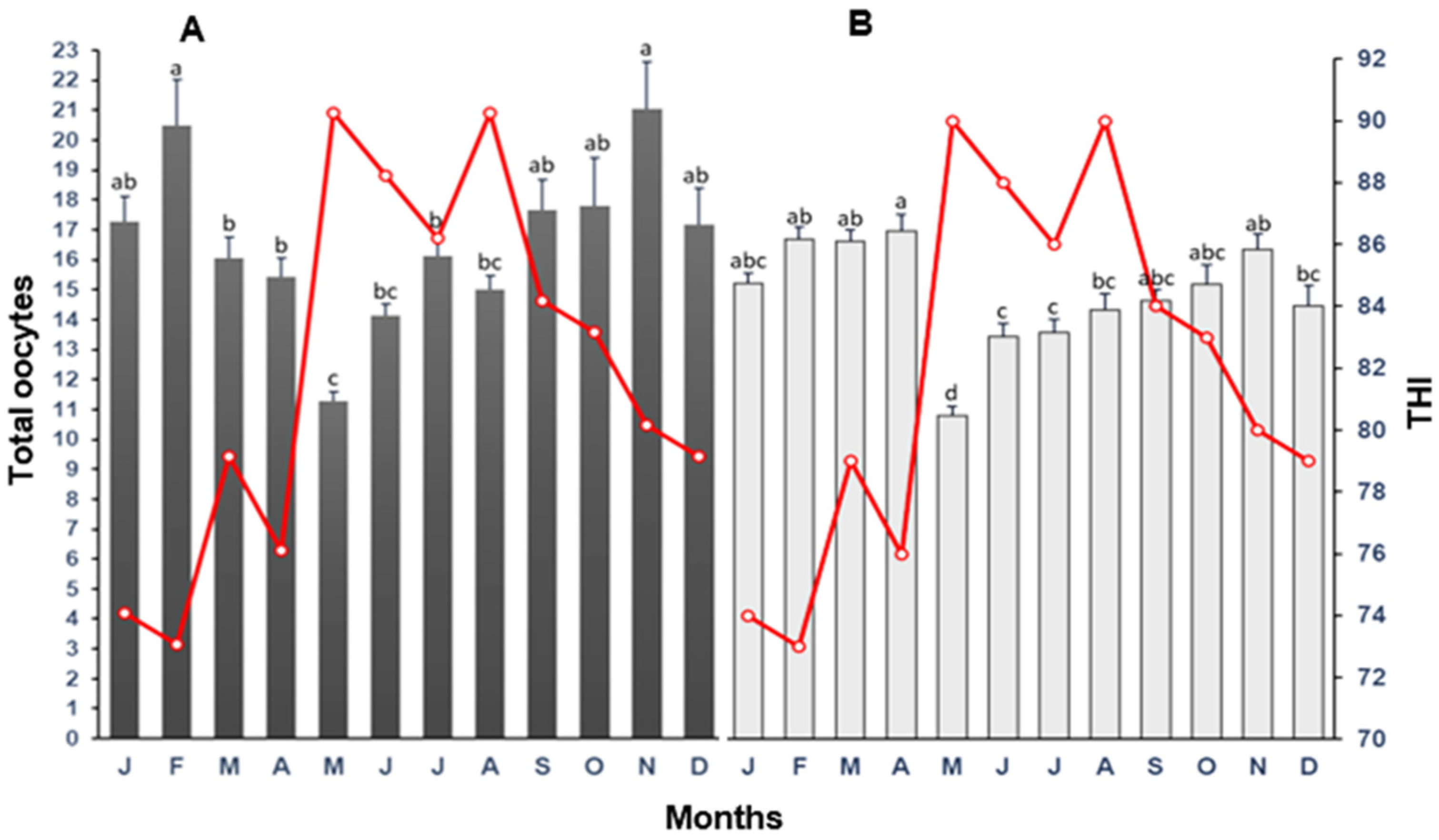
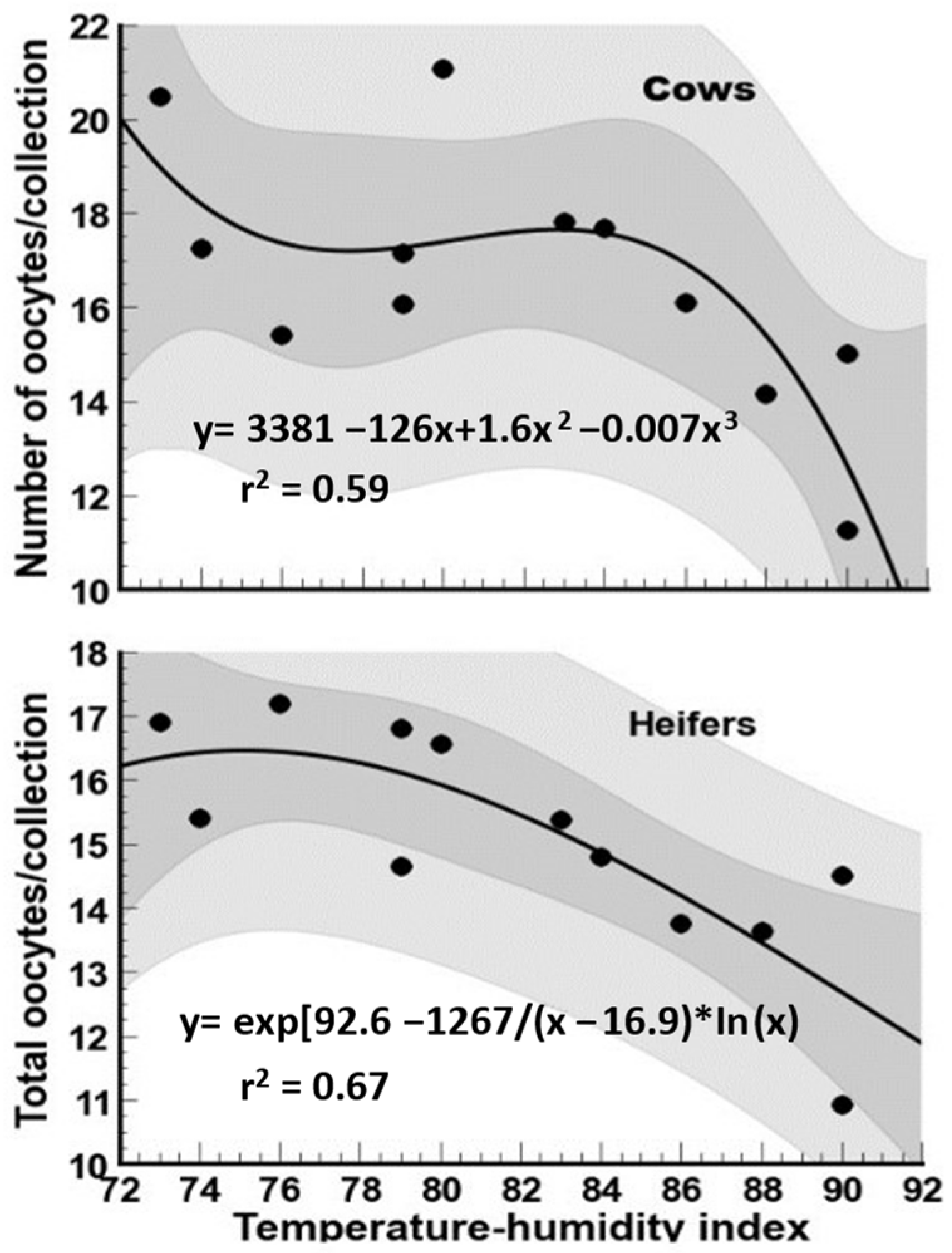
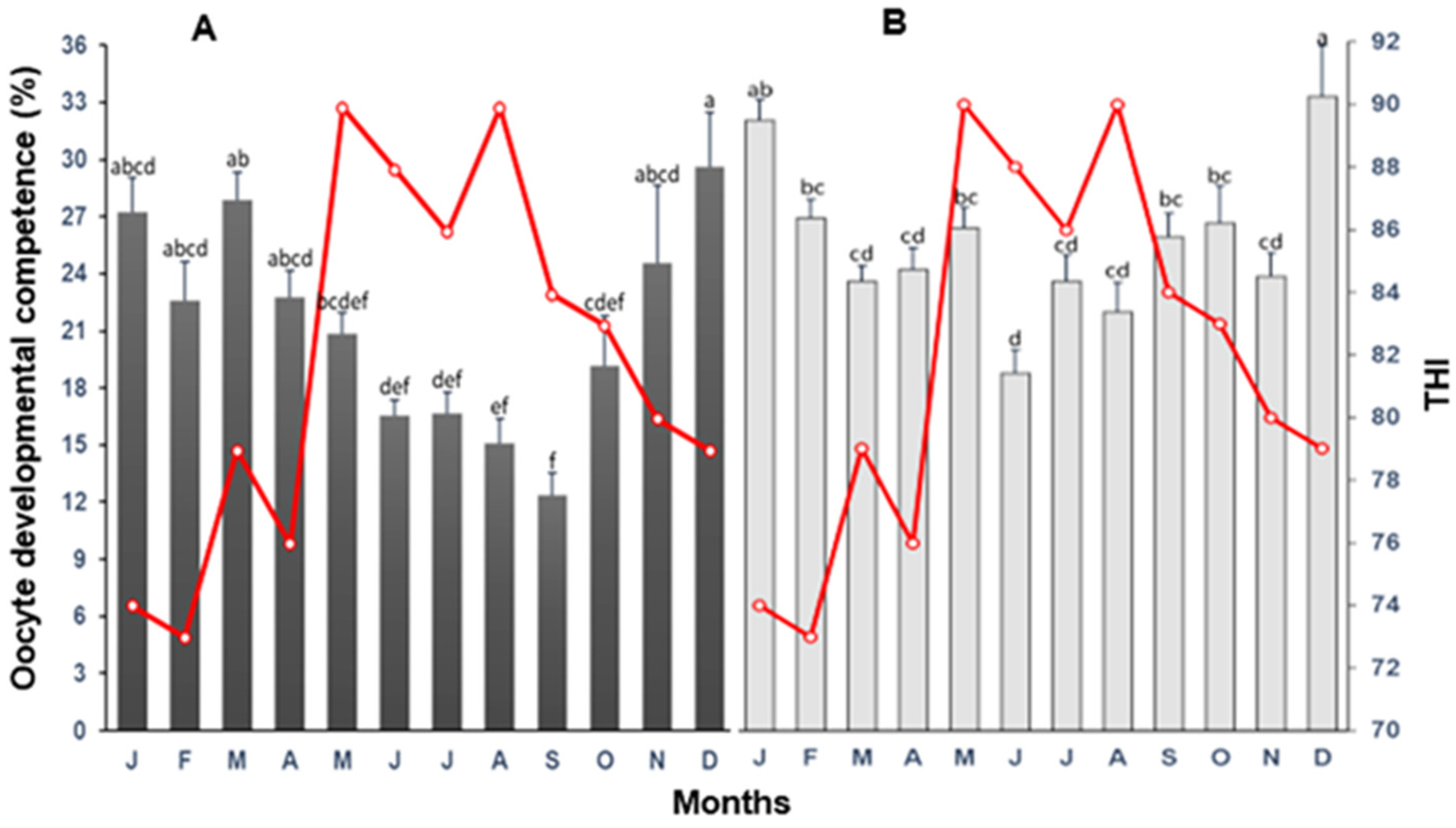
3.1. Total Oocytes
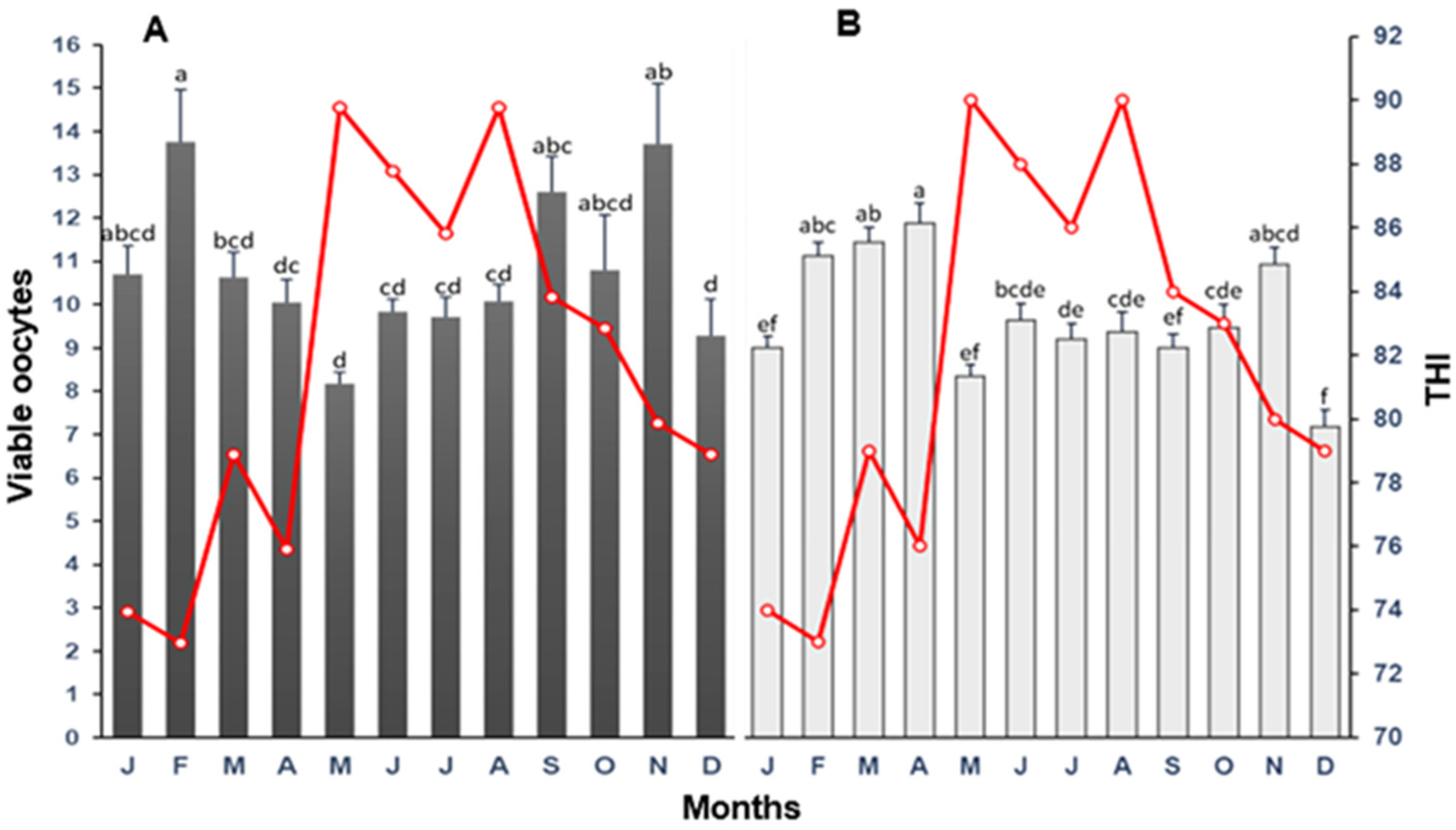
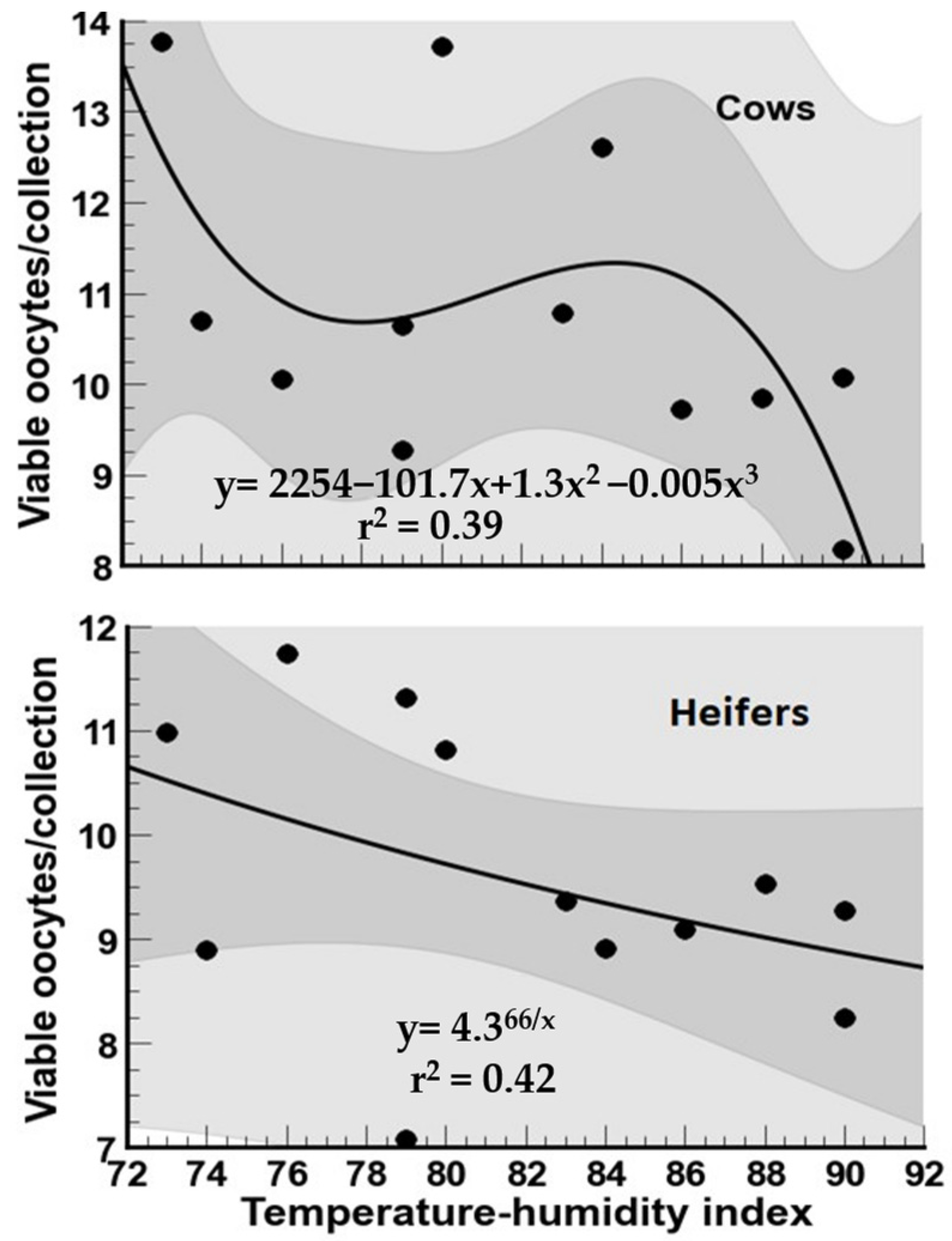
3.2. Viable Oocytes
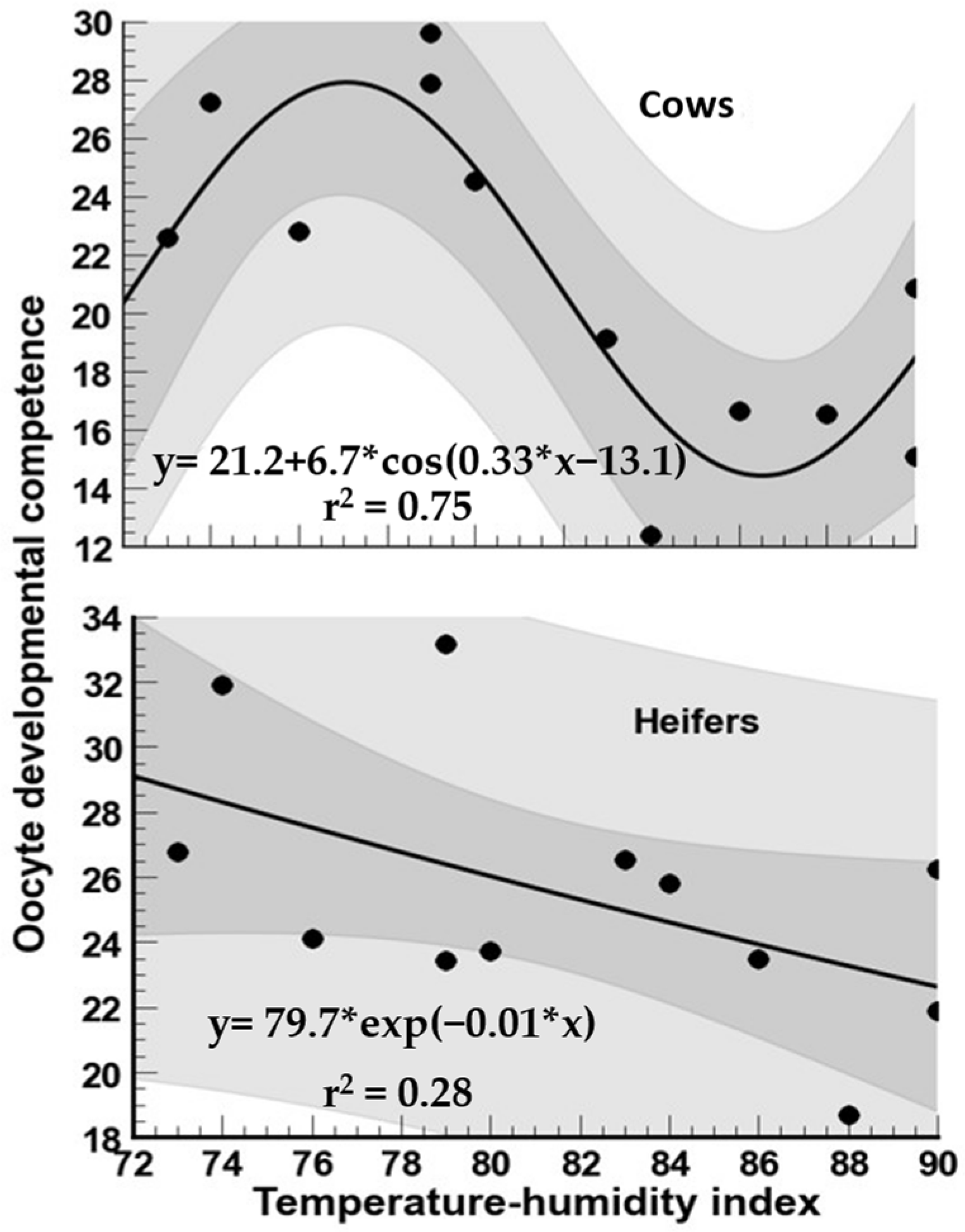
3.3. Oocyte Developmental Competence
3.4. Total Embryos
4. Discussion
4.1. Total Oocytes
4.2. Oocyte Viability
4.3. Oocyte Developmental Competence
4.4. Embryo Production
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sammad, A.; Umer, S.; Shi, R.; Zhu, H.; Zhao, X.; Wang, Y. Dairy cow reproduction under the influence of heat stress. J. Anim. Physiol. Anim. Nutr. 2020, 104, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, J.; Wang, F.; Xiao, J.; Wang, Y.; Yang, H.; Li, S.; Cao, Z. Heat stress on calves and heifers: A review. J. Anim. Sci. Biotechnol. 2020, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.L.; Bogart, A.N.; Rispoli, L.A.; Saxton, M.A.; Schrick, F.N. Developmental competence of bovine embryos from heat-stressed ova. J. Dairy Sci. 2009, 92, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Alemu, W.T.; Pandey, O.H.; Wondim, S.D.; Gebremedhn, S.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Oxidative and endoplasmic reticulum stress defense mechanisms of bovine granulosa cells exposed to heat stress. Theriogenology 2018, 110, 130–141. [Google Scholar] [CrossRef]
- Amamou, H.; Beckers, Y.; Mahouachi, M.; Hammami, H. Thermotolerance indicators related to production and physiological responses to heat stress of holstein cows. J. Therm. Biol. 2019, 82, 90–98. [Google Scholar] [CrossRef]
- Polsky, L.B.; Augusto, M.L.; Madureira, E.L.; Drago, F.; Soriano, S.; Sica, F.; Vasconcelos, M.L.; Cerri, L.A.R. Association between ambient temperature and humidity, vaginal temperature, and automatic activity monitoring on induced estrus in lactating cows. J. Dairy Sci. 2017, 100, 8590–8601. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Li, R.; Wu, Y.; Zhang, D.; Xu, H.; Zhang, Y.; Qi, Z. Effect of seasonal thermal stress on oxidative status, immune response and stress hormones of lactating dairy cows. Anim. Nutr. 2021, 7, 216–223. [Google Scholar] [CrossRef]
- Rungruang, S.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H.; De Veth, M.J.; Collier, R.J. A dose-response evaluation of rumen-protected niacin in thermoneutral or heat-stressed lactating Holstein cows. J. Dairy Sci. 2014, 97, 5023–5034. [Google Scholar] [CrossRef]
- Becker, C.A.; Collier, R.J.; Stone, A.E. Invited review: Physiological and behavioral effects of heat stress in dairy cows. J. Dairy Sci. 2020, 103, 6751–6770. [Google Scholar] [CrossRef]
- Slimen, B.I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef]
- Nishisozu, T.; Singh, J.; Abe, A.; Okamura, K.; Dochi, O. Effects of the temperature-humidity index on conception rates in Holstein heifers and cows receiving in vitro-produced Japanese Black cattle embryos. J. Reprod. Dev. 2023, 69, 72–77. [Google Scholar] [CrossRef]
- Lopes, F.F.P.; Lima, R.S.; Risolia, P.H.B.; Ispada, J.; Assumpcao, M.E.O.A.; Visintin, J.A. Heat stress induced alteration in bovine oocytes: Funtional and cellular aspects. Anim. Reprod. 2012, 9, 395–403. [Google Scholar]
- Khan, A.; Khan, M.Z.; Umer, S.; Khan, I.M.; Xu, H.; Zhu, H.; Wang, Y. Cellular and Molecular Adaptation of Bovine Granulosa Cells and Oocytes under Heat Stress. Animals 2012, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, X.A.; Smith, J.F.; Bradford, B.J.; Harner, J.P.; Oddy, A. A comparison of the effects of 2 cattle-cooling systems on dairy cows in a desert environment. J. Dairy Sci. 2010, 93, 4955–4960. [Google Scholar] [CrossRef]
- De Aguiar, H.L.; Hyde, A.H.; Pedrosa, H.G.; Denicol, C.A. Heat stress of impairs in vitro development of preantral follicles of cattle. Anim. Reprod. Sci. 2020, 213, 106277. [Google Scholar] [CrossRef] [PubMed]
- Mellado, M.; Sepulveda, E.; Meza-Herrera, C.; Veliz, F.; Arevalo, J.; Mellado, J.; De Santiago, A. Effects of heat stress on reproductive efficiency of high yielding Holstein cows in a hot-arid environment. Rev. Colomb. Cienc. Pec. 2013, 26, 193–200. [Google Scholar]
- Ferreira, R.M.; Ayres, H.; Chiaratti, M.R.; Ferraz, M.L.; Araújo, A.B.; Rodrigues, C.A.; Watanabe, Y.F.; Vireque, A.A.; Joaquim, D.C.; Smith, L.C.; et al. The low fertility of repeat-breeder cows during summer heat stress is related to a low oocyte competence to develop into blastocysts. J. Dairy Sci. 2011, 94, 2383–2392. [Google Scholar] [CrossRef] [PubMed]
- Abdelatty, A.M.; Iwaniuk, M.E.; Potts, S.B.; Gad, A. Influence of maternal nutrition and heat stress on bovine oocyte and embryo development. Int. J. Vet. Sci. Med. 2018, 6, S1–S5. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Exploitation of genetic and physiological determinants of embryonic resistance to elevated temperature to improve embryonic survival in dairy cattle during heat stress. Theriogenology 2007, 68, S242–S249. [Google Scholar] [CrossRef]
- Bényei, B.; Gáspárdy, A.; Barros, C.W.C. Changes in embryo production results and ovarian recrudescence during the acclimatisation to the semiarid tropics of embryo donor Holstein–Friesian cows raised in a temperate climate. Anim. Reprod. Sci. 2001, 68, 57–68. [Google Scholar] [CrossRef]
- Fernandez-Novo, A.; Pérez-Garnelo, S.S.; Villagrá, A.; Pérez-Villalobos, N.; Astiz, S. The effect of stress on reproduction and reproductive technologies in beef cattle—A review. Animals 2020, 10, 2096. [Google Scholar] [CrossRef]
- Payton, R.R.; Romar, R.; Coy, P.; Saxton, A.M.; Lawrence, J.L.; Edwards, J.L. Susceptibility of bovine germinal vesicle-stage oocytes from antral follicles to direct effects of heat stress in vitro. Biol. Reprod. 2004, 71, 1303–1308. [Google Scholar] [CrossRef]
- Ingraham, R.H.; Gillette, D.D.; Wagner, W.D. Relationship of temperature and humidity to conception rate of Holstein cows in subtropical climate. J. Dairy Sci. 1974, 57, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Pavani, K.; Carvalhais, I.; Faheem, M.; Chaveiro, A.; Reis, F.V.; da Silva, F.M. Reproductive performance of Holstein dairy cows grazing in dry-summer subtropical climatic conditions: Effect of heat stress and heat shock on meiotic competence and in vitro fertilization. Asian Australas J. Anim. Sci. 2015, 28, 334. [Google Scholar] [CrossRef]
- Ratchamak, R.; Ratsiri, T.; Chumchai, R.; Boonkum, W.; Chankitisakul, V. Relationship of the temperature-humidity index (THI) with ovarian responses and embryo production in superovulated Thai-Holstein crossbreds under tropical climate conditions. Vet. Sci. 2021, 8, 270. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Gallego, H.Z.; Calderon-Leyva, G.; Angel-Garcia, O.; Guillen-Muñoz, J.M.; Leyva, C.; Mellado, M.; Pedroso, R.; Pessoa, L.G.; Esparza, C.; Morales, J.L. Effect of Season on Quantity and Competence of Oocytes Recovered Transvaginally from Holstein Cows for In vitro Fertilization. Indian J. Anim. Res. 2021, 1, 4. [Google Scholar] [CrossRef]
- FASS. Guide for the Care and Use of Agricultural Animals. In Agricultural Research and Teaching, 3rd ed.; Federation Animal Science Society: Champaing, IL, USA, 2010; p. 177. [Google Scholar]
- NAM-National Academy of Medicine. Co-Produced by the National Academy of Medicine–Mexico and the Association for Assessment and Accreditation of Laboratory Animal Care International. In Guide for the Care and Use of Laboratory Animals, 1st ed.; Harlan: Mexico City, Mexico, 2010. [Google Scholar]
- INIFAP-Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. Red Nacional de Estaciones Agrometeorológicas Automatizadas 2018. Available online: https://clima.inifap.gob.mx/lnmysr/Estaciones (accessed on 21 March 2021).
- Mader, T.L.; Davis, M.S.; Brown-Brandl, T. Environmental factors influencing heat stress in feedlot cattle. J. Anim. Sci. 2006, 84, 712–719. [Google Scholar] [CrossRef]
- Armstrong, D. Heat stress interaction with shade and cooling. J. Dairy Sci. 1994, 77, 2044–2050. [Google Scholar] [CrossRef]
- López, G.; Brizuela, A.B.; Rondán, G.; Lissaso, C.M.; Alejandra, C.; Kemerer, A.C.; de los Santos, M. Determinación del índice de temperatura y humedad (ITH) para vacas lecheras, en el departamento Nogoyá, entre ríos. Rev. Cient. Agropecu. 2016, 20, 57–65. [Google Scholar]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Influence of Body Condition Score on Relationships Between Metabolic Status and Oxidative Stress in Periparturient Dairy Cows. J. Dairy Sci. 2005, 88, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Avendaño-Reyes, L.; Correa-Calderón, A.; Macías-Cruz, U.; García-Casillas, A.C.; Mellado, M.; Robinson, P.H.; Hernández-Rivera, J.A. Impacts on two dairy breeds of adding a third (night) cooling event under extreme ambient heat. Int. J. Biometeorol. 2021, 65, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- NRC-National Research Council. Nutrient Requirements of Dairy Cattle: Seventh Revised Edition; The National Academies Press: Washington, DC, USA, 2001. [CrossRef]
- Solís, C.A.; Guerra, R.; Sandoya, G.; De Armas, R. Efecto de sincronización de la onda folicular y de la frecuencia de aspiración de folículos en novillas de la raza Brahman. Red. Vet. 2012, 13, 1–16. [Google Scholar]
- De Loos, F.; Van, V.C.; Van, M.P.; Kruip, T.A. Morphology of Immature Bovine Oocytes. Gamete Res. 1989, 24, 197–204. [Google Scholar] [CrossRef]
- Paula-Lopes, F.F.; Hansen, P.J. Heat shock-induced apoptosis in preimplantation bovine embryos is a developmentally regulated phenomenon. Biol. Reprod. 2002, 66, 1169–1177. [Google Scholar] [CrossRef]
- Bó, G.A.; Mappletoft, R.J. Evaluation and classification of bovine embryos. Anim. Reprod. 2013, 10, 344–348. [Google Scholar]
- Roth, Z. Physiology and endocrinology symposium: Cellular and molecular mechanisms of heat stress related to bovine ovarian function. J. Anim. Sci. 2015, 93, 2034–2044. [Google Scholar] [CrossRef] [PubMed]
- Rust, J.M.; Visser, D.S.; Venter, J.E.; Boshoff, M.P.; Foss, S.; Greyling, J.P. The effect of season on aspects of in vitro embryo production in sub-fertile beef cows. S. Afr. J. Anim. Sci. 2009, 39, 275–279. [Google Scholar]
- Roth, Z. Heat Stress, the Follicle, and Its Enclosed Oocyte: Mechanisms and Potential Strategies to Improve Fertility in Dairy Cows. Reprod. Domest. Anim. 2008, 43, 238–244. [Google Scholar] [CrossRef]
- Wolfenson, D.; Roth, Z. Impact of heat stress on cow reproduction and fertility. Anim. Front. 2019, 9, 32–38. [Google Scholar] [CrossRef]
- Roth, Z. Heat stress reduces maturation and developmental capacity in bovine oocytes. Reprod. Fertil. Dev. 2021, 33, 66–75. [Google Scholar] [CrossRef]
- Gendelman, M.; Aroyo, A.; Yavin, S.; Roth, Z. Seasonal effects on gene expression, cleavage timing, and developmental competence of bovine preimplantation embryos. Reproduction 2010, 140, 73–82. [Google Scholar] [CrossRef]
- Roth, Z. Influence of heat stress on reproduction in dairy cows—Physiological and practical aspects. J. Anim. Sci. 2020, 98, S80–S87. [Google Scholar] [CrossRef]
- Wolfenson, D.; Thatcher, W.W.; Badinga, L.J.; Savio, D.R.; Meidan, B.J.; Lew, R.; Braw-Tal, R.; Berman, A. Effect of heat stress on follicular development during the estrous cycle in lactating dairy cattle. Biol. Reprod. 1995, 52, 1106–1113. [Google Scholar] [CrossRef]
- Roth, Z. Reproductive physiology and endocrinology responses of cows exposed to environmental heat stress—Experiences from the past and lessons for the present. Theriogenology 2020, 155, 150–156. [Google Scholar] [CrossRef]
- Wang, J.Z.; Sui, H.S.; Miao, D.Q.; Liu, N.; Zhou, P.; Ge, L.; Tan, J.H. Effects of heat stress during in vitro maturation on cytoplasmic versus nuclear components of mouse oocytes. Reproduction 2009, 137, 181–189. [Google Scholar] [CrossRef]
- Ju, J.C.; Jiang, S.; Tseng, J.K.; Parks, J.E.; Yang, X. Heat shock reduces developmental competence and alters spindle configuration of bovine oocytes. Theriogenology 2005, 64, 1677–1689. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z.; Hansen, P.J. Disruption of nuclear maturation and rearrangement of cytoskeletal elements in bovine oocytes exposed to heat shock during maturation. Reproduction 2005, 129, 235–244. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, Q.Y. Evaluation of oocyte quality: Morphological, celular and molecular predictors. Reprod. Fertil. Dev. 2007, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Reproductive physiology of the heat-stressed dairy cow: Implications for fertility and assisted reproduction. Anim. Reprod. 2019, 16, 497–507. [Google Scholar] [CrossRef]
- Sartori, R.; Sartor-Bergfelt, R.; Mertens, S.A.; Guenther, J.N.; Parrish, J.J.; Wiltbank, M.C. Fertilization and early embryonic development in heifers and lactating cows in summer and lactating and dry cows in winter. J. Dairy Sci. 2002, 85, 2803–2812. [Google Scholar] [CrossRef] [PubMed]
- Al-Katanani, Y.M.; Paula-Lopes, F.F.; Hansen, P.J. Effect of Season and Exposure to Heat Stress on Oocyte Competence in Holstein Cows. J. Dairy Sci. 2002, 85, 390–396. [Google Scholar] [CrossRef]
- Pontes, J.H.F.; Silva, K.C.F.; Basso, A.C.; Rigo, A.G.; Ferreira, C.R.; Santos, G.M.G.; Sanches, B.V.; Porcionato, J.P.F.; Vieira, P.H.S.; Faifer, F.S.; et al. Large-scale in vitro embryo production and pregnancy rates from Bos taurus, Bos indicus, and indicus-taurus dairy cows using sexed sperm. Theriogenology 2010, 74, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z. Symposium review: Reduction in oocyte developmental competence by stress is associated with alterations in mitochondrial function. J. Dairy Sci. 2018, 101, 3642–3654. [Google Scholar] [CrossRef]
- Lopez-Gatius, F. Is fertility declining in dairy cattle? A retrospective study in northeastern Spain. Theriogenology 2003, 60, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Sakatani, M. Effects of heat stress on bovine preimplantation embryos produced in vitro. J. Reprod. Dev. 2017, 63, 347–352. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Chiaratti, M.R.; Macabelli, C.H.; Rodrigues, C.A.; Ferraz, M.L.; Watanabe, Y.F.; Smith, L.C.; Meirelles, F.V.; Baruselli, P.S. The Infertility of Repeat-Breeder Cows During Summer Is Associated with Decreased Mitochondrial DNA and Increased Expression of Mitochondrial and Apoptotic Genes in Oocytes. Biol. Reprod. 2016, 94, 66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Months | Average (°C) | Maximun | Minimun | Average (THI) | Average Relative Humidity |
|---|---|---|---|---|---|
| January | 14.65 (±2.13) | 29 | 1 | 74 | 34 |
| February | 14.85 (±2.77) | 28 | 2 | 73 | 35 |
| March | 20.79 (±1.94) | 35 | 7 | 79 | 27 |
| April | 21.80 (±1.55) | 30 | 5 | 76 | 19 |
| May | 27.42 (±1.56 | 42 | 10 | 90 | 24 |
| June | 29.30 (±1.34) | 39 | 17 | 88 | 31 |
| July | 28.88 (±1.24) | 39 | 20 | 86 | 40 |
| August | 27.94 (±1.27) | 38 | 16 | 90 | 42 |
| September | 24.81 (±1.48) | 37 | 15 | 84 | 42 |
| Octuber | 22.03 (±1.55) | 33 | 11 | 83 | 49 |
| November | 17.83 (±1.86) | 34 | 2 | 80 | 57 |
| December | 11.93 (±1.58) | 27 | −5 | 79 | 56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Cruz, J.L.; Calderon-Leyva, G.; Angel-García, O.; Guillen-Muñoz, J.M.; Santos-Jimenez, Z.; Mellado, M.; Pessoa, L.G.; Guerrero-Gallego, H.Z. The Effect of Month of Harvesting and Temperature–Humidity Index on the Number and Quality of Oocytes and In Vitro Embryo Production in Holstein Cows and Heifers. Biology 2023, 12, 1174. https://doi.org/10.3390/biology12091174
Morales-Cruz JL, Calderon-Leyva G, Angel-García O, Guillen-Muñoz JM, Santos-Jimenez Z, Mellado M, Pessoa LG, Guerrero-Gallego HZ. The Effect of Month of Harvesting and Temperature–Humidity Index on the Number and Quality of Oocytes and In Vitro Embryo Production in Holstein Cows and Heifers. Biology. 2023; 12(9):1174. https://doi.org/10.3390/biology12091174
Chicago/Turabian StyleMorales-Cruz, Juan L., Guadalupe Calderon-Leyva, Oscar Angel-García, Juan M. Guillen-Muñoz, Zurisaday Santos-Jimenez, Miguel Mellado, Luiz Gustavo Pessoa, and Hugo Z. Guerrero-Gallego. 2023. "The Effect of Month of Harvesting and Temperature–Humidity Index on the Number and Quality of Oocytes and In Vitro Embryo Production in Holstein Cows and Heifers" Biology 12, no. 9: 1174. https://doi.org/10.3390/biology12091174
APA StyleMorales-Cruz, J. L., Calderon-Leyva, G., Angel-García, O., Guillen-Muñoz, J. M., Santos-Jimenez, Z., Mellado, M., Pessoa, L. G., & Guerrero-Gallego, H. Z. (2023). The Effect of Month of Harvesting and Temperature–Humidity Index on the Number and Quality of Oocytes and In Vitro Embryo Production in Holstein Cows and Heifers. Biology, 12(9), 1174. https://doi.org/10.3390/biology12091174