
The Effects of Doxapram Blocking the Response of Gram-Negative Bacterial Toxin (LPS) at Glutamatergic Synapses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
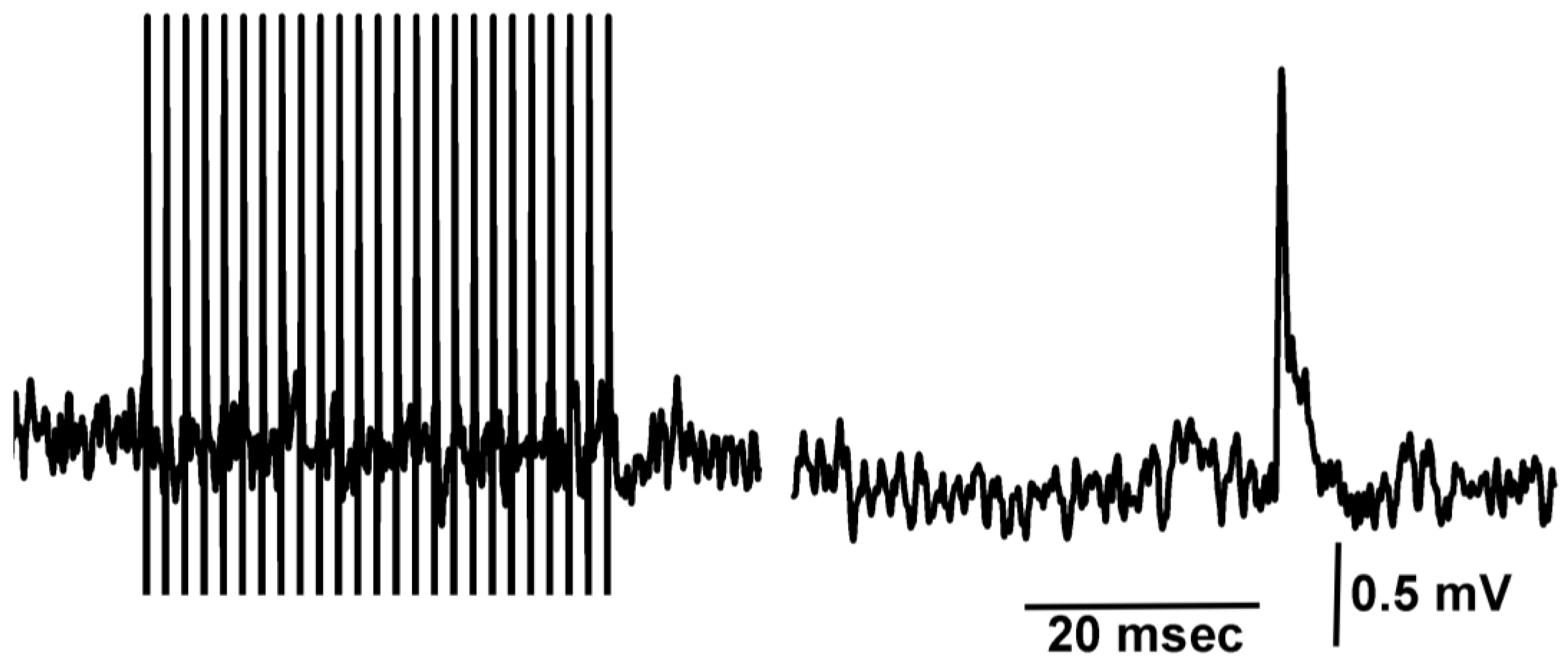
2.2. Neuromuscular Junction
2.3. Chemicals
2.4. Statistical Analysis
3. Results
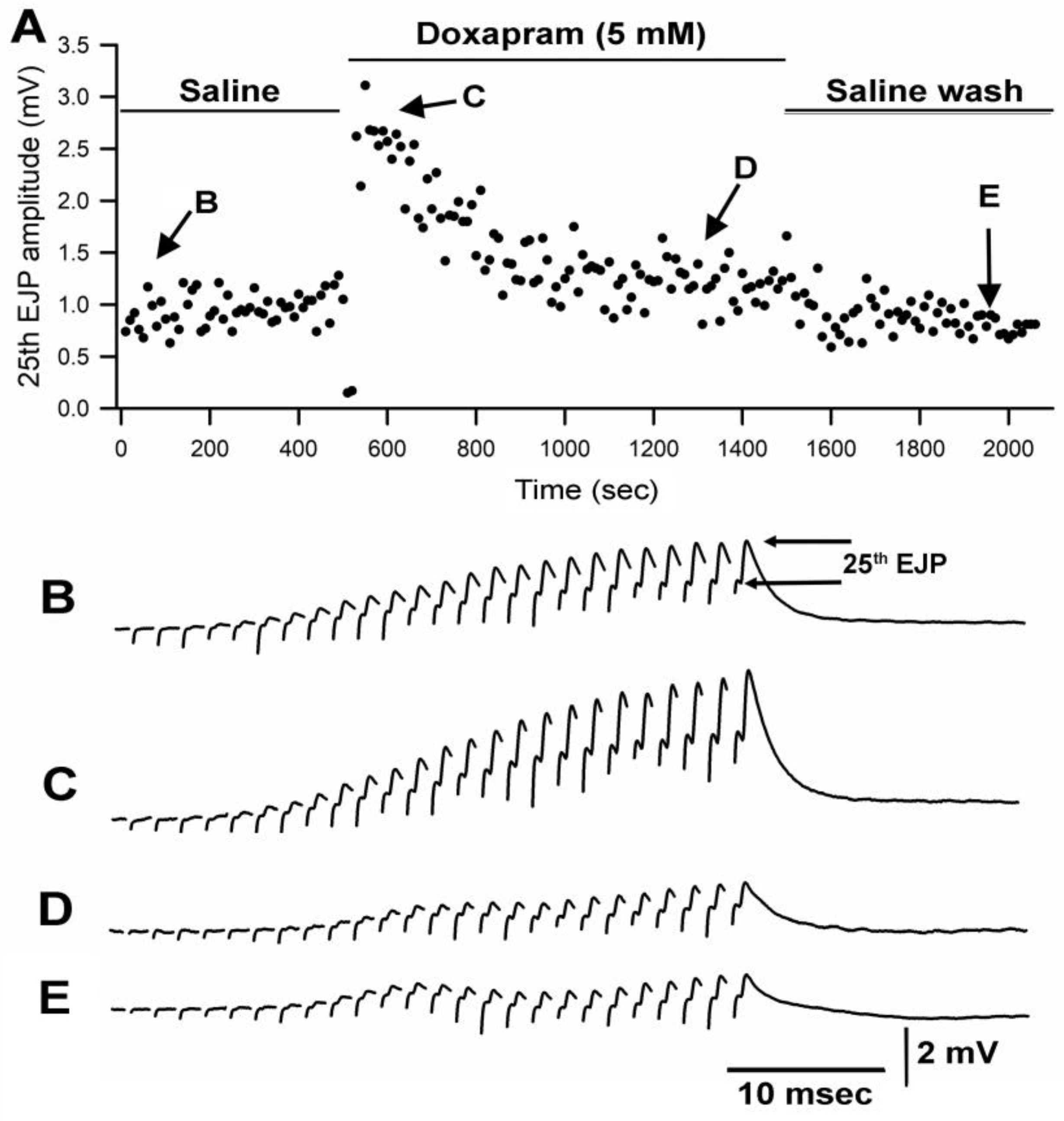
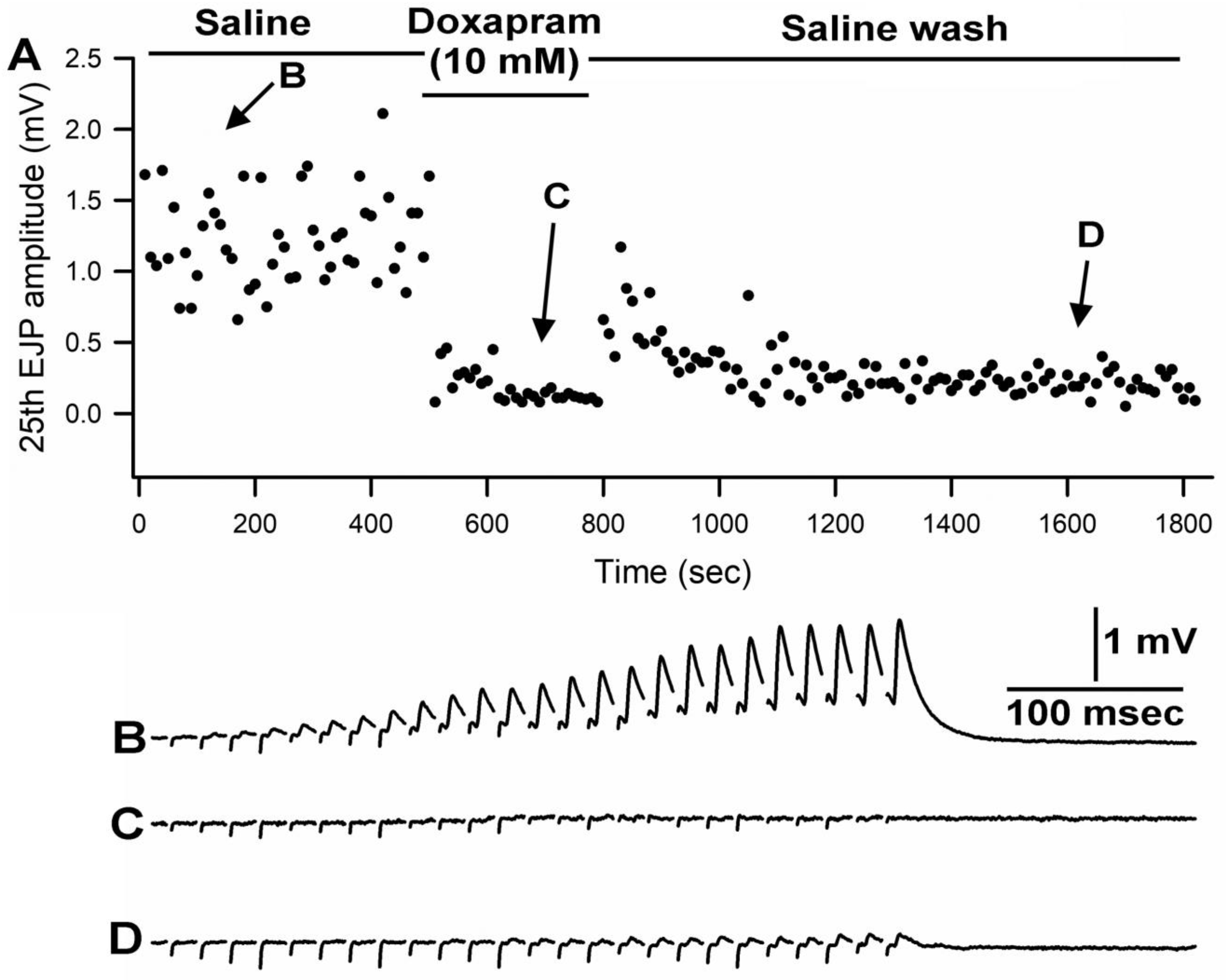
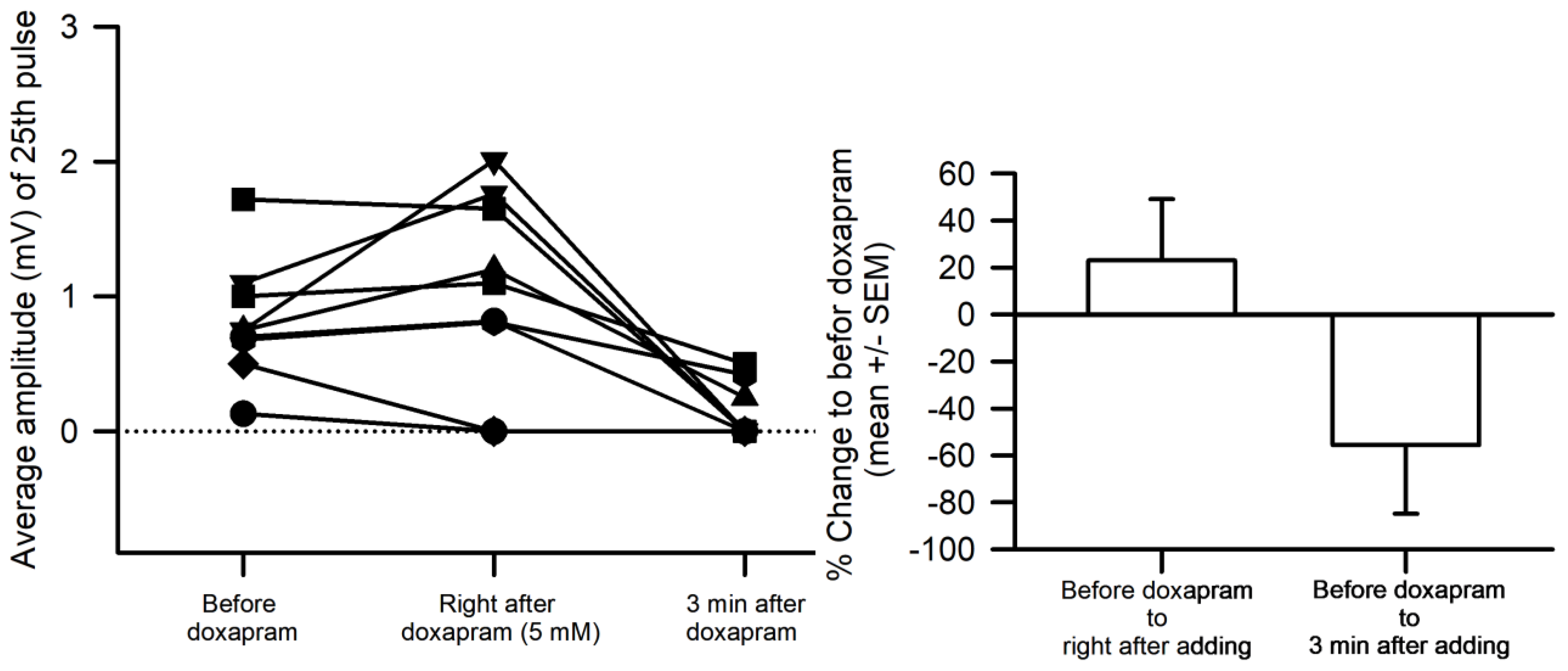
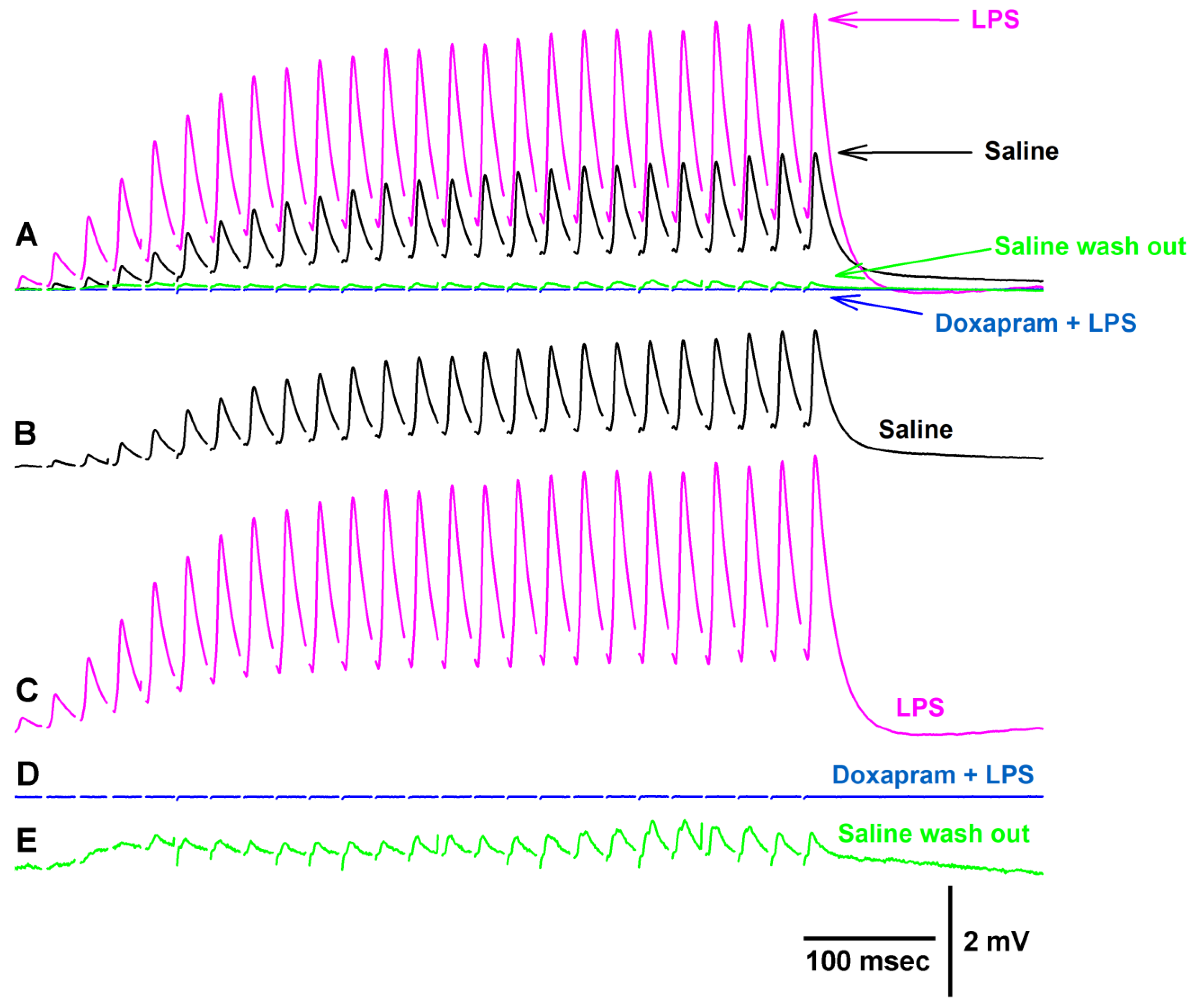
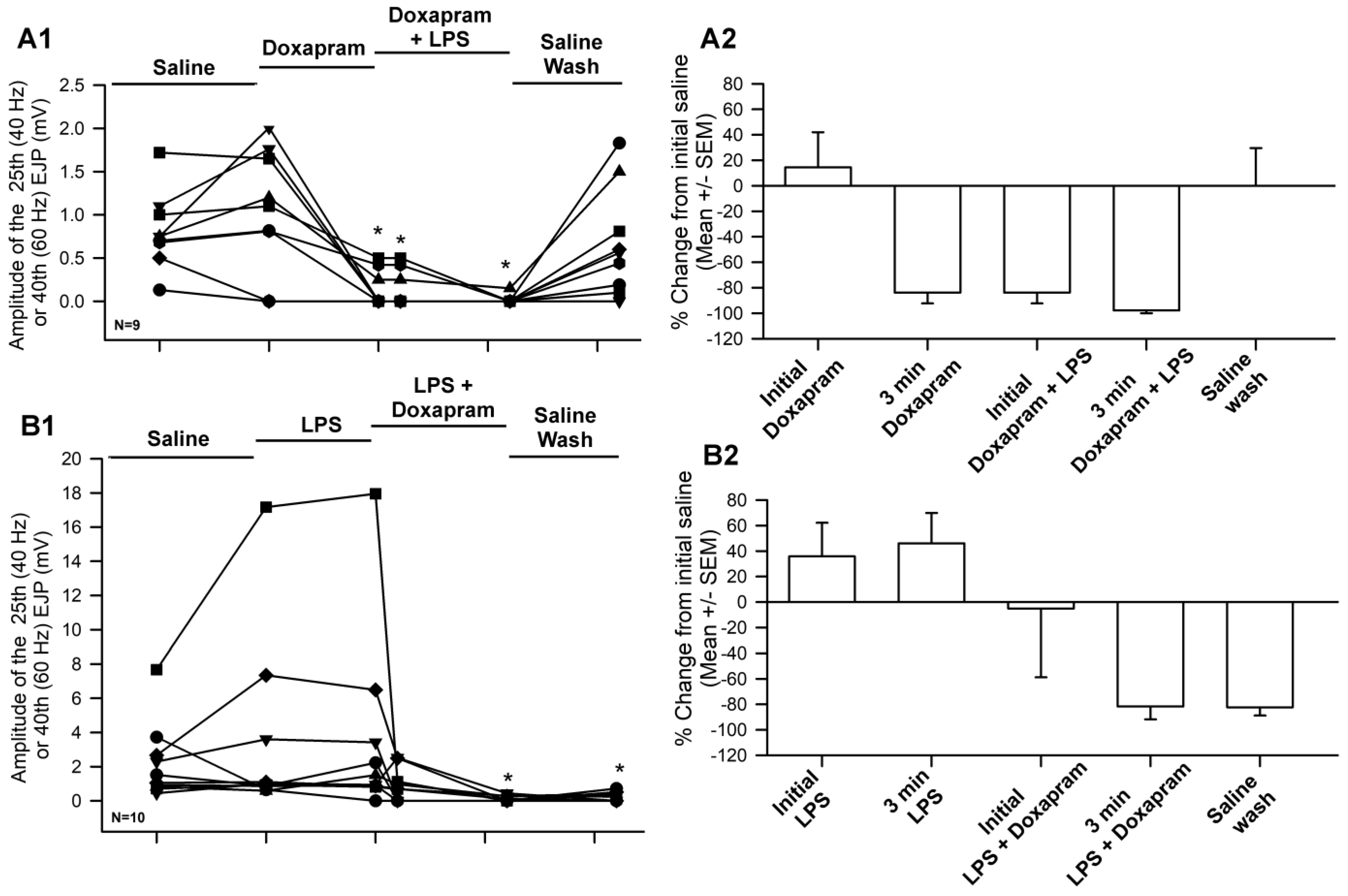
3.1. Effect of Doxapram Enhancing and Depressing Synaptic Transmission
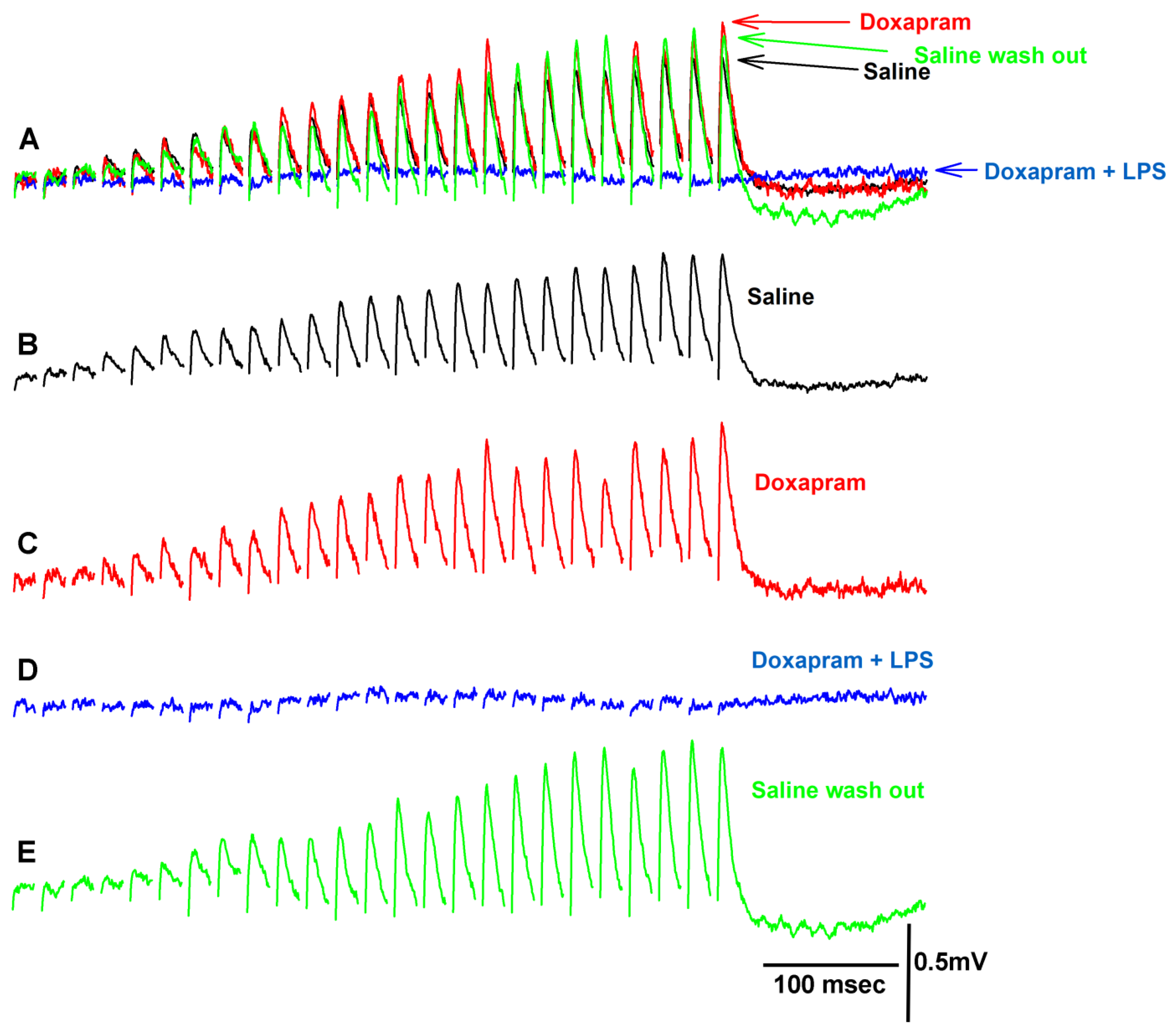
3.2. The LPS Can Dampen the Effect of Doxapram
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steiner, H. Peptidoglycan recognition proteins: On and off switches for innate immunity. Immunol. Rev. 2004, 198, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, S.D.; Kidd, J.F.; Law, R.J.; Franks, C.J.; Sattelle, D.B. Structure and function of two-pore-domain K+ channels: Contributions from genetic model organisms. Trends Pharmacol. Sci. 2005, 26, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Model. Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Yoshida, H. Drosophila as a model organism. Adv. Exp. Med. Biol. 2018, 1076, 1–10. [Google Scholar] [PubMed]
- Estévez-Lao, T.Y.; Sigle, L.T.; Gomez, S.N.; Hillyer, J.F. Nitric oxide produced by periostial hemocytes modulates the bacterial infection-induced reduction of the mosquito heart rate. J. Exp. Biol. 2020, 223 Pt 15, jeb225821. [Google Scholar] [CrossRef]
- Coscia, M.; Giacomelli, S.; Oreste, U. Toll-like receptors: An overview from invertebrates to vertebrates. Invert. Surv. J. 2011, 8, 210–226. [Google Scholar]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Loker, E.S.; Adema, C.M.; Zhang, S.M.; Kepler, T.B. Invertebrate immune systems—Not homogeneous, not simple, not well understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef]
- Kleino, A.; Silverman, N. The Drosophila IMD pathway in the activation of the humoral immune response. Dev. Com. Immunol. 2014, 42, 25–35. [Google Scholar] [CrossRef]
- Gerth, A.; Grosche, J.; Nieber, K.; Hauschildt, S. Intracellular LPS inhibits the activity of potassium channels and fails to activate NFkappaB in human macrophages. J. Cell Physiol. 2005, 202, 442–452. [Google Scholar] [CrossRef]
- Cooper, R.L.; Krall, R.M. Hyperpolarization Induced by Lipopolysaccharides but Not by Chloroform Is Inhibited by Doxapram, an Inhibitor of Two-P-Domain K+ Channel (K2P). Int. J. Mol. Sci. 2022, 23, 15787. [Google Scholar] [CrossRef] [PubMed]
- Lorenzon, S.; de Guarrini, S.; Smith, V.; Ferrero, E. Effects of LPS injection on circulating haemocytes in crustaceans in vivo. Fish Shellfish Immunol. 1999, 9, 31–50. [Google Scholar] [CrossRef]
- Andrä, J.; Garidel, P.; Majerle, A.; Jerala, R.; Ridge, R.; Paus, E.; Novitsky, T.; Koch, M.H.J.; Brandendurg, K. Biophysical characterization of the interaction of Limulus polyphemus endotoxin neutralizing protein with lipopolysaccharide. Eur. J. Biochem. 2004, 271, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Novitsky, T.J. Limulus amebocyte lysate (LAL) detection of endotoxin in human blood. J. Endotoxin Res. 1994, 1, 253–263. [Google Scholar] [CrossRef]
- US Department Health and Human Services. Guideline on the Validation of the Limulus Amebocyte Lysate Test as an End-Product Endotoxin Test for Human and Animal Parenteral Drugs, Biological Products, and Medical Devices. 1987. Available online: http://citeseerx.ist.psu.edu/viewdoc/download;jsessionid=3A346C6757AD03CB41B6DA54067C9648?doi=10.1.1.178.656&rep=rep1&type=pdf (accessed on 26 December 2019).
- Ballinger-Boone, C.; Anyagaligbo, O.; Bernard, J.; Bierbower, S.M.; Dupont-Versteegden, E.E.; Ghoweri, A.; Greenhalgh, A.; Harrison, D.; Istas, O.; McNabb, M.; et al. The effects of bacterial endotoxin (LPS) on cardiac and synaptic function in various animal models: Larval Drosophila, crawfish, crab, and rodent. Int. J. Zool. Res. 2020, 16, 33–62. [Google Scholar] [CrossRef]
- Greenhalgh, A.; Istas, O.; Cooper, R.L. Bacterial endotoxin lipopolysaccharide enhances synaptic transmission at low-output glutamatergic synapses. Neurosci. Res. 2021, 170, 59–65. [Google Scholar] [CrossRef]
- Plant, L.D.; Goldstein, S.A.N. Two-Pore Domain Potassium Channels. In Handbook of Ion Channels, 1st ed.; Zheng, J., Trudeau, M.C., Eds.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9780429193965. [Google Scholar]
- Vacassenno, R.M.; Haddad, C.N.; Cooper, R.L. The effects on resting membrane potential and synaptic transmission by Doxapram (blocker of K2P channels) at the Drosophila neuromuscular junction. Comp. Biochem. Physiol. C 2023, 263, 109497. [Google Scholar] [CrossRef]
- Lorenzon, S.; Pasqual, P.; Ferrero, E.A. Different bacterial lipopolysaccharides as toxicants and stressors in the shrimp Palaemon elegans. Fish Shellfish Immunol. 2002, 13, 27–45. [Google Scholar] [CrossRef]
- Pien, B.C.; Sundaram, P.; Raoof, N.; Costa, S.F.; Mirrett, S.; Woods, C.W.; Reller, L.B.; Weinstein, M.P. The clinical and prognostic importance of positive blood cultures in adults. Am. J. Med. 2010, 123, 819–828. [Google Scholar] [CrossRef]
- Carl, M.A.; Ndao, I.M.; Springman, A.C.; Manning, S.D.; Johnson, J.R.; Johnston, B.D.; Burnham, C.A.; Weinstock, E.S.; Weinstock, G.M.; Wylie, T.N.; et al. Sepsis from the gut: The enteric habitat of bacteria that cause late-onset neonatal bloodstream infections. Clin. Infect. Dis. 2014, 58, 1211–1218. [Google Scholar] [CrossRef]
- Kim, S.B.; Jeon, Y.D.; Kim, J.H.; Kim, J.K.; Ann, H.W.; Choi, H.; Kim, M.H.; Song, J.E.; Ahn, J.Y.; Jeong, S.J.; et al. Risk factors for mortality in patients with Serratia marcescens bacteremia. Yonsei Med. J. 2015, 56, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Joyner, J.; Wanless, D.; Sinigalliano, C.D.; Lipp, E.K. Use of quantitative real-time PCR for direct detection of serratia marcescens in marine and other aquatic environments. Appl. Environ. Microbiol. 2014, 80, 1679–1683. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ríos, M.; Lapesa-Lázaro, S.; Larumbe-Arricibita, J.; Alonso-Gutiérrez, F.; Galindo-Parrila, F.J.; Martín-Torrijos, L.; Diéguez-Uribeondo, J. Resistance to Crayfish Plague: Assessing the Response of Native Iberian Populations of the White-Clawed Freshwater Crayfish. J. Fungi 2022, 8, 342. [Google Scholar] [CrossRef]
- Cooper, A.S.; Cooper, R.L. Historical view and physiology demonstration at the NMJ of the crayfish opener muscle. J. Vis. Exp. 2009, 33, e1595. [Google Scholar] [CrossRef]
- Sparks, G.; Cooper, R.L. 5-HT offsets homeostasis of synaptic transmission during short-term facilitation. J. Appl. Physiol. 2004, 96, 1681–1690. [Google Scholar] [CrossRef]
- Parnas, I.; Reinhold, R.; Fine, J. Synaptic transmission in the crayfish: Increased release of transmitter substance by bacterial endotoxin. Science 1971, 171, 1153–1155. [Google Scholar] [CrossRef] [PubMed]
- Elliott, E.R.; Taul, A.C.; Abul-Khoudoud, M.O.; Hensley, N.; Cooper, R.L. Effect of doxapram, bacterial endotoxin and pH on heart rate: Larval Drosophila model. Appl. Biosci. 2023, accepted. [Google Scholar]
- Cotton, J.F. TASK-1 (KCNK3) and TASK-3 (KCNK9) tandem pore potassium channel antagonists stimulate breathing in isoflurane-anesthetized rats. Anesth. Analg. 2013, 116, 810–816. [Google Scholar] [CrossRef]
- Kim, D. Physiology and pharmacology of two-pore domain potassium channels. Curr. Pharm. Des. 2005, 11, 2717–2736. [Google Scholar] [CrossRef]
- Bierbower, S.M.; Cooper, R.L. The effects of acute carbon dioxide on behavior and physiology in Procambarus clarkii. J. Exp. Zool. 2010, 313A, 484–497. [Google Scholar] [CrossRef]
- Goldstein, S.A.; Price, L.A.; Rosenthal, D.N.; Pausch, M.H. ORK1, a potassium-selective leak channel with two pore domains cloned from Drosophila melanogaster by expression in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1996, 93, 13256–13261. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.A.; Wang, K.W.; Ilan, N.; Pausch, M.H. Sequence and function of the two P domain potassium channels: Implications of an emerging superfamily. J. Mol. Med. 1998, 76, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Belvin, M.P.; Anderson, K.V. A conserved signaling pathway: The Drosophila Toll-dorsal pathway. Ann. Rev. Cell Dev. Biol. 1996, 12, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996, 86, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.W.; White, T.D. Some commercial preparations of Escherichia coli bacterial endotoxin lipopolysaccaride (LPS) are contaminated with biologically active substances. J. Neurochem. 2001, 78, 1183–1184. [Google Scholar] [CrossRef]
- Hirschfeld, M.; Ma, Y.; Weis, J.H.; Vogel, S.N.; Weis, J.J. Cutting edge: Repurification of lipopolysaccharide eliminates signaling through both human and murine toll-like receptor 2. J. Immunol. 2000, 165, 618–622. [Google Scholar] [CrossRef]
- Ochoa-Cortes, F.; Ramos-Lomas, T.; Miranda-Morales, M.; Spreadbury, I.; Ibeakanma, C.; Barajas-Lopez, C.; Vanner, S. Bacterial cell products signal to mouse colonic nociceptive dorsal root ganglia neurons. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G723–G732. [Google Scholar] [CrossRef]
- Linhartova, I.; Bumba, L.; Masin, J.; Basler, M.; Osicka, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef]
- Gray, M.; Szabo, G.; Otero, A.S.; Gray, L.; Hewlett, E. Distinct mechanisms for K+ efflux, intoxication and hemolysis by Bordetella pertussis AC toxin. J. Biol. Chem. 1998, 273, 18260–18267. [Google Scholar] [CrossRef] [PubMed]
- Hertle, R. Serratia type pore forming toxins. Curr. Protein Pept. Sci. 2000, 1, 75–89. [Google Scholar] [CrossRef]
- Flint, R.B.; Simons, S.H.P.; Andriessen, P.; Liem, K.D.; Degraeuwe, P.L.J.; Reiss, I.K.M.; Heine, R.T.; Engbers, A.G.J.; Koch, B.C.P.; de Groot, R.; et al. The bioavailability and maturing clearance of doxapram in preterm infants. Pediatr. Res. 2021, 89, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.C.; Klemm, W.R. Clinical evaluation of the analeptic, doxapram, in dogs and cats. J. Am. Vet. Med. Assoc. 1967, 150, 516–525. [Google Scholar] [PubMed]
- Karklus, A.A.; Sladky, K.K.; Johnson, S.M. Respiratory and antinociceptive effects of dexmedetomidine and doxapram in ball pythons (Python regius). Am. J. Vet. Res. 2021, 82, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L. Clinical evaluation of doxapram hydrochloride, a respiratory stimulant. J. Okla. State Med. Assoc. 1973, 66, 481–487. [Google Scholar] [PubMed]
- Yost, C.S. A new look at the respiratory stimulant doxapram. CNS Drug Rev. 2006, 12, 236–249. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brock, K.E.; Cooper, R.L. The Effects of Doxapram Blocking the Response of Gram-Negative Bacterial Toxin (LPS) at Glutamatergic Synapses. Biology 2023, 12, 1046. https://doi.org/10.3390/biology12081046
Brock KE, Cooper RL. The Effects of Doxapram Blocking the Response of Gram-Negative Bacterial Toxin (LPS) at Glutamatergic Synapses. Biology. 2023; 12(8):1046. https://doi.org/10.3390/biology12081046
Chicago/Turabian StyleBrock, Kaitlyn E., and Robin L. Cooper. 2023. "The Effects of Doxapram Blocking the Response of Gram-Negative Bacterial Toxin (LPS) at Glutamatergic Synapses" Biology 12, no. 8: 1046. https://doi.org/10.3390/biology12081046
APA StyleBrock, K. E., & Cooper, R. L. (2023). The Effects of Doxapram Blocking the Response of Gram-Negative Bacterial Toxin (LPS) at Glutamatergic Synapses. Biology, 12(8), 1046. https://doi.org/10.3390/biology12081046