
Current Status of Omics in Biological Quality Elements for Freshwater Biomonitoring



Abstract
Simple Summary
Abstract
1. Introduction
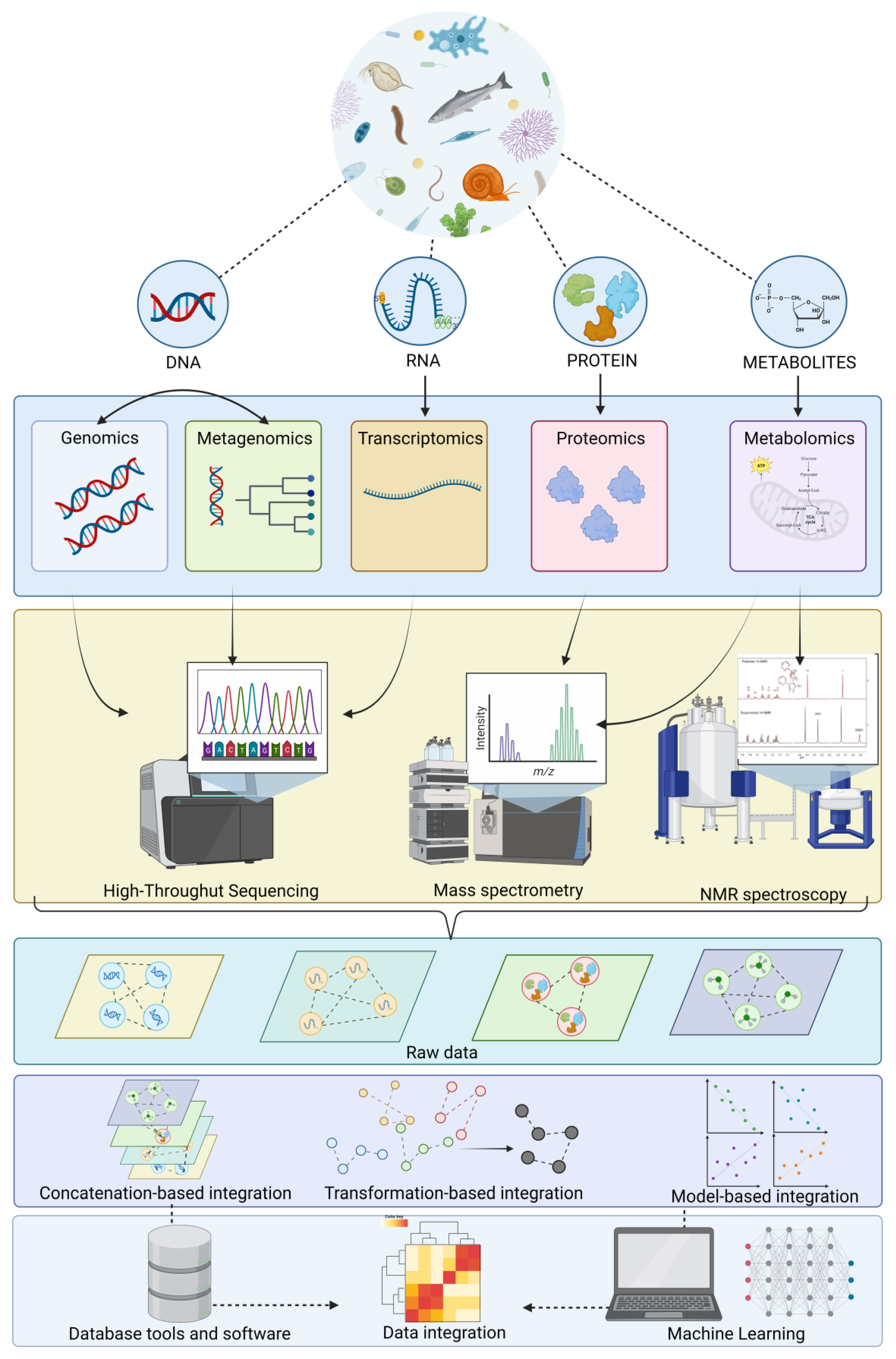
2. State-of-Art of Omics in Freshwater Biomonitoring
2.1. How Biological Quality Elements (BQEs) Can Be Used in Omics
2.2. Genomics
2.3. Metagenomics
2.4. Transcriptomics
2.5. Proteomics
2.6. Metabolomics
2.7. Multi-Omics
3. Use of Omics on Freshwater Biomonitoring: Perspectives and Challenges
3.1. Omics in Freshwater Biomonitoring: A Multifaceted Framework
3.2. Omics in a Scenario of More Severe Impacts
3.3. Omics Data and Informatics Tools
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keck, F.; Vasselon, V.; Tapolczai, K.; Rimet, F.; Bouchez, A. Freshwater Biomonitoring in the Information Age. Front. Ecol. Environ. 2017, 15, 266–274. [Google Scholar] [CrossRef]
- Maitland, V.C.; Robinson, C.V.; Porter, T.M.; Hajibabaei, M. Freshwater Diatom Biomonitoring through Benthic Kick-Net Metabarcoding. PLoS ONE 2020, 15, e0242143. [Google Scholar] [CrossRef]
- Persaud, S.F.; Cottenie, K.; Gleason, J.E. Ethanol EDNA Reveals Unique Community Composition of Aquatic Macroinvertebrates Compared to Bulk Tissue Metabarcoding in a Biomonitoring Sampling Scheme. Diversity 2021, 13, 34. [Google Scholar] [CrossRef]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA: For Biodiversity Research and Monitoring; Oxford University Press: Oxford, UK, 2018; ISBN 0198767226. [Google Scholar]
- López-Pedrouso, M.; Varela, Z.; Franco, D.; Fernández, J.A.; Aboal, J.R. Can Proteomics Contribute to Biomonitoring of Aquatic Pollution? A Critical Review. Environ. Pollut. 2020, 267, 115473. [Google Scholar] [CrossRef]
- Gibson, J.F.; Shokralla, S.; Curry, C.; Baird, D.J.; Monk, W.A.; King, I.; Hajibabaei, M. Large-Scale Biomonitoring of Remote and Threatened Ecosystems via High-Throughput Sequencing. PLoS ONE 2015, 10, e0138432. [Google Scholar] [CrossRef]
- Woodward, G.; Gray, C.; Baird, D.J. Biomonitoring for the 21st Century: New Perspectives in an Age of Globalisation and Emerging Environmental Threats. Limnetica 2013, 32, 159–174. [Google Scholar] [CrossRef]
- Singh, J.; Dutta, T.; Kim, K.-H.; Rawat, M.; Samddar, P.; Kumar, P. ‘Green’ Synthesis of Metals and Their Oxide Nanoparticles: Applications for Environmental Remediation. J. Nanobiotechnol. 2018, 16, 84. [Google Scholar] [CrossRef]
- Valentin, V.; Frédéric, R.; Isabelle, D.; Olivier, M.; Yorick, R.; Agnès, B. Assessing Pollution of Aquatic Environments with Diatoms’ DNA Metabarcoding: Experience and Developments from France Water Framework Directive Networks. Metabarcoding Metagenom. 2019, 3, e39646. [Google Scholar] [CrossRef]
- Sumudumali, R.G.I.; Jayawardana, J. A Review of Biological Monitoring of Aquatic Ecosystems Approaches: With Special Reference to Macroinvertebrates and Pesticide Pollution. Environ. Manag. 2021, 67, 263–276. [Google Scholar] [CrossRef]
- Wilkinson, J.L.; Boxall, A.B.A.; Kolpin, D.W.; Leung, K.M.Y.; Lai, R.W.S.; Galbán-Malagón, C.; Adell, A.D.; Mondon, J.; Metian, M.; Marchant, R.A. Pharmaceutical Pollution of the World’s Rivers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113947119. [Google Scholar] [CrossRef]
- Mehana, E.-S.E.; Khafaga, A.F.; Elblehi, S.S.; El-Hack, M.E.A.; Naiel, M.A.E.; Bin-Jumah, M.; Othman, S.I.; Allam, A.A. Biomonitoring of Heavy Metal Pollution Using Acanthocephalans Parasite in Ecosystem: An Updated Overview. Animals 2020, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Zolkefli, N.; Sharuddin, S.S.; Yusoff, M.Z.M.; Hassan, M.A.; Maeda, T.; Ramli, N. A Review of Current and Emerging Approaches for Water Pollution Monitoring. Water 2020, 12, 3417. [Google Scholar] [CrossRef]
- Scherer, C.; Weber, A.; Lambert, S.; Wagner, M. Interactions of Microplastics with Freshwater Biota. In Freshwater Microplastics; Springer: Cham, Switzerland, 2018; pp. 153–180. [Google Scholar]
- Santos, J.M.; Ferreira, M.T. Use of Aquatic Biota to Detect Ecological Changes in Freshwater: Current Status and Future Directions. Water 2020, 12, 1611. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, X.; Chen, Y.; Fu, R.; Du, X.; Chen, X.; Zhan, A. Zooplankton Biodiversity Monitoring in Polluted Freshwater Ecosystems: A Technical Review. Environ. Sci. Ecotechnol. 2020, 1, 100008. [Google Scholar] [CrossRef]
- Gatti, R.C. Freshwater Biodiversity: A Review of Local and Global Threats. Int. J. Environ. Stud. 2016, 73, 887–904. [Google Scholar] [CrossRef]
- Pawlowski, J.; Lejzerowicz, F.; Apotheloz-Perret-Gentil, L.; Visco, J.; Esling, P. Protist Metabarcoding and Environmental Biomonitoring: Time for Change. Eur. J. Protistol. 2016, 55, 12–25. [Google Scholar] [CrossRef]
- Fierro, P.; Valdovinos, C.; Vargas-Chacoff, L.; Bertrán, C.; Arismendi, I. Macroinvertebrates and Fishes as Bioindicators of Stream Water Pollution. In Water Quality; Intechopen: Rijeka, Croatia, 2017; pp. 23–38. [Google Scholar]
- Gál, B.; Szivák, I.; Heino, J.; Schmera, D. The Effect of Urbanization on Freshwater Macroinvertebrates–Knowledge Gaps and Future Research Directions. Ecol. Indic. 2019, 104, 357–364. [Google Scholar] [CrossRef]
- Pandey, L.K.; Bergey, E.A.; Lyu, J.; Park, J.; Choi, S.; Lee, H.; Depuydt, S.; Oh, Y.T.; Lee, S.M.; Han, T. The Use of Diatoms in Ecotoxicology and Bioassessment: Insights, Advances and Challenges. Water Res. 2017, 118, 39–58. [Google Scholar] [CrossRef]
- Cimarelli, L.; Singh, K.S.; Mai, N.T.N.; Dhar, B.C.; Brandi, A.; Brandi, L.; Spurio, R. Molecular Tools for the Selective Detection of Nine Diatom Species Biomarkers of Various Water Quality Levels. Int. J. Environ. Res. Public Health 2015, 12, 5485–5504. [Google Scholar] [CrossRef]
- Lainé, M.; Morin, S.; Tison-Rosebery, J. A Multicompartment Approach-Diatoms, Macrophytes, Benthic Macroinvertebrates and Fish-to Assess the Impact of Toxic Industrial Releases on a Small French River. PLoS ONE 2014, 9, e102358. [Google Scholar] [CrossRef]
- Smeti, E.; von Schiller, D.; Karaouzas, I.; Laschou, S.; Vardakas, L.; Sabater, S.; Tornés, E.; Monllor-Alcaraz, L.S.; Guillem-Argiles, N.; Martinez, E. Multiple Stressor Effects on Biodiversity and Ecosystem Functioning in a Mediterranean Temporary River. Sci. Total Environ. 2019, 647, 1179–1187. [Google Scholar] [CrossRef]
- Guariento, R.D.; Carneiro, L.S.; Caliman, A.; Bozelli, R.L.; Leal, J.J.F.; de Assis Esteves, F. Interactive Effects of Omnivorous Fish and Nutrient Loading on Net Productivity Regulation of Phytoplankton and Periphyton. Aquat. Biol. 2010, 10, 273–282. [Google Scholar] [CrossRef]
- Faquim, R.C.P.; Machado, K.B.; Teresa, F.B.; de Oliveira, P.H.F.; Granjeiro, G.F.; Vieira, L.C.G.; Nabout, J.C. Shortcuts for Biomonitoring Programs of Stream Ecosystems: Evaluating the Taxonomic, Numeric, and Cross-Taxa Congruence in Phytoplankton, Periphyton, Zooplankton, and Fish Assemblages. PLoS ONE 2021, 16, e0258342. [Google Scholar] [CrossRef]
- Tzafesta, E.; Zangaro, F.; Specchia, V.; Pinna, M. An Overview of DNA-Based Applications for the Assessment of Benthic Macroinvertebrates Biodiversity in Mediterranean Aquatic Ecosystems. Diversity 2021, 13, 112. [Google Scholar] [CrossRef]
- Brantschen, J.; Blackman, R.C.; Walser, J.C.; Altermatt, F. Environmental Dna Gives Comparable Results to Morphology-Based Indices of Macroinvertebrates in a Large-Scale Ecological Assessment. PLoS ONE 2021, 16, e0257510. [Google Scholar] [CrossRef]
- Uchida, N.; Kubota, K.; Aita, S.; Kazama, S. Aquatic Insect Community Structure Revealed by EDNA Metabarcoding Derives Indices for Environmental Assessment. PeerJ 2020, 8, e9176. [Google Scholar] [CrossRef]
- Fenoglio, S.; de Figueroa, J.M.T.; Doretto, A.; Falasco, E.; Bona, F. Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems. Water 2020, 12, 2934. [Google Scholar] [CrossRef]
- Fierro, P.; Valdovinos, C.; Arismendi, I.; Díaz, G.; Jara-Flores, A.; Habit, E.; Vargas-Chacoff, L. Examining the Influence of Human Stressors on Benthic Algae, Macroinvertebrate, and Fish Assemblages in Mediterranean Streams of Chile. Sci. Total Environ. 2019, 686, 26–37. [Google Scholar] [CrossRef]
- Zohary, T.; Flaim, G.; Sommer, U. Temperature and the Size of Freshwater Phytoplankton. Hydrobiologia 2021, 848, 143–155. [Google Scholar] [CrossRef]
- Ebner, J.N.; Ritz, D.; von Fumetti, S. Comparative Proteomics of Stenotopic Caddisfly Crunoecia Irrorata Identifies Acclimation Strategies to Warming. Mol. Ecol. 2019, 28, 4453–4469. [Google Scholar] [CrossRef]
- Bonacina, L.; Fasano, F.; Mezzanotte, V.; Fornaroli, R. Effects of Water Temperature on Freshwater Macroinvertebrates: A Systematic Review. Biol. Rev. 2023, 98, 191–221. [Google Scholar] [CrossRef] [PubMed]
- Mierzejewska, E.; Urbaniak, M. Molecular Methods as Potential Tools in Ecohydrological Studies on Emerging Contaminants in Freshwater Ecosystems. Water 2020, 12, 2962. [Google Scholar] [CrossRef]
- Mortimer, M.; Fang, W.; Zhou, X.; Vodovnik, M.; Guo, L.-H. Omics Approaches in Toxicological Studies. In Advances in Toxicology and Risk Assessment of Nanomaterials and Emerging Contaminants; Springer: Singapore, 2022; pp. 61–94. [Google Scholar]
- Cordier, T.; Lanzén, A.; Apothéloz-Perret-Gentil, L.; Stoeck, T.; Pawlowski, J. Embracing Environmental Genomics and Machine Learning for Routine Biomonitoring. Trends. Microbiol. 2019, 27, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Elbrecht, V.; Leese, F. Validation and Development of COI Metabarcoding Primers for Freshwater Macroinvertebrate Bioassessment. Front. Environ. Sci. 2017, 5, 11. [Google Scholar]
- Leese, F.; Bouchez, A.; Abarenkov, K.; Altermatt, F.; Borja, Á.; Bruce, K.; Ekrem, T.; Čiampor, F., Jr.; Čiamporová-Zaťovičová, Z.; Costa, F.O.; et al. Why We Need Sustainable Networks Bridging Countries, Disciplines, Cultures and Generations for Aquatic Biomonitoring 2.0: A Perspective Derived from the DNAqua-Net COST Action. Adv. Ecol. Res. 2018, 58, 63–99. [Google Scholar]
- Prabhakaran, K.; Nagarajan, R.; Franco, F.M.; Kumar, A.A. Biomonitoring of Malaysian Aquatic Environments: A Review of Status and Prospects. Ecohydrol. Hydrobiol. 2017, 17, 134–147. [Google Scholar] [CrossRef]
- Pomfret, S.M.; Brua, R.B.; Izral, N.M.; Yates, A.G. Metabolomics for Biomonitoring: An Evaluation of the Metabolome as an Indicator of Aquatic Ecosystem Health. Environ. Rev. 2020, 28, 89–98. [Google Scholar] [CrossRef]
- Garcia-Reyero, N.; Adelman, I.R.; Martinović, D.; Liu, L.; Denslow, N.D. Site-specific impacts on gene expression and behavior in fathead minnows (Pimephales promelas) exposed in situ to streams adjacent to sewage treatment plants. BMC Bioinform. 2009, 11, S11. [Google Scholar] [CrossRef]
- Archambault, J.M.; Cope, W.G.; Kwak, T.J. Burrowing, Byssus, and Biomarkers: Behavioral and Physiological Indicators of Sublethal Thermal Stress in Freshwater Mussels (Unionidae). Mar. Freshw. Behav. Physiol. 2013, 46, 229–250. [Google Scholar] [CrossRef]
- Hutchins, C.M.; Simon, D.F.; Zerges, W.; Wilkinson, K.J. Transcriptomic Signatures in Chlamydomonas reinhardtii as Cd Biomarkers in Metal Mixtures. Aquat. Toxicol. 2010, 100, 120–127. [Google Scholar] [CrossRef]
- Taylor, N.S.; Merrifield, R.; Williams, T.D.; Chipman, J.K.; Lead, J.R.; Viant, M.R. Molecular Toxicity of Cerium Oxide Nanoparticles to the Freshwater Alga Chlamydomonas reinhardtii Is Associated with Supra-Environmental Exposure Concentrations. Nanotoxicology 2016, 10, 32–41. [Google Scholar] [PubMed]
- Du, C.; Zhang, B.; He, Y.; Hu, C.; Ng, Q.X.; Zhang, H.; Ong, C.N. Biological Effect of Aqueous C60 Aggregates on Scenedesmus Obliquus Revealed by Transcriptomics and Non-Targeted Metabolomics. J. Hazard Mater. 2017, 324, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Kahlert, M.; Almeida, S.F.P.; Figueira, E. Assessing Cu Impacts on Freshwater Diatoms: Biochemical and Metabolomic Responses of Tabellaria flocculosa (Roth) Kützing. Sci. Total Environ. 2018, 625, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Bai, Y.; Chen, Z.; Mo, J.; Li, Q.; Sun, H.; Zhang, Q. Transcriptomic Analysis Suggests the Inhibition of DNA Damage Repair in Green Alga Raphidocelis subcapitata Exposed to Roxithromycin. Ecotoxicol. Environ. Saf. 2020, 201, 110737. [Google Scholar] [CrossRef]
- Yong, W.K.; Sim, K.S.; Poong, S.W.; Wei, D.; Phang, S.M.; Lim, P.E. Interactive Effects of Warming and Copper Toxicity on a Tropical Freshwater Green Microalga Chloromonas augustae (Chlorophyceae). J. Appl. Phycol. 2021, 33, 67–77. [Google Scholar] [CrossRef]
- Sahlin, S. The Next-Generation of Aquatic Effect-Based Monitoring?: A Critical Review about the Application, Challenges and Barriers with Omics in Field. Master’s Thesis, Linnaeus University, Växjö, Sweden, 2019. [Google Scholar]
- Siciliano, A.; Gesuele, R. How Daphnia (Cladocera) Assays May Be Used as Bioindicators of Health Effects? J. Biodivers. Endanger. Species 2013, 1, 1–6. [Google Scholar] [CrossRef]
- Liang, X.; Martyniuk, C.J.; Simmons, D.B.D. Are We Forgetting the “Proteomics” in Multi-Omics Ecotoxicology? Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 36, 100751. [Google Scholar] [CrossRef]
- Olenici, A.; Baciu, C.; Blanco, S.; Morin, S. Naturally and Environmentally Driven Variations in Diatom Morphology: Implications for Diatom-Based Assessment of Water Quality. In Modern Trends in Diatom Identification; Springer: Cham, Switzerland, 2020; pp. 39–50. [Google Scholar]
- Ge, Y.; Xia, C.; Wang, J.; Zhang, X.; Ma, X.; Zhou, Q. The Efficacy of DNA Barcoding in the Classification, Genetic Differentiation, and Biodiversity Assessment of Benthic Macroinvertebrates. Ecol. Evol. 2021, 11, 5669–5681. [Google Scholar] [CrossRef]
- Churko, J.M.; Mantalas, G.L.; Snyder, M.P.; Wu, J.C. Overview of High Throughput Sequencing Technologies to Elucidate Molecular Pathways in Cardiovascular Diseases. Circ. Res. 2013, 112, 1613–1623. [Google Scholar] [CrossRef]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N. Environmental DNA Metabarcoding: Transforming How We Survey Animal and Plant Communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Colin, N.; Porte, C.; Fernandes, D.; Barata, C.; Padrós, F.; Carrassón, M.; Monroy, M.; Cano-Rocabayera, O.; de Sostoa, A.; Piña, B. Ecological Relevance of Biomarkers in Monitoring Studies of Macro-Invertebrates and Fish in Mediterranean Rivers. Sci. Total Environ. 2016, 540, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xia, P.; Wang, P.; Yang, J.; Baird, D.J. Omics Advances in Ecotoxicology. Environ. Sci. Technol. 2018, 52, 3842–3851. [Google Scholar] [CrossRef] [PubMed]
- Prat, O.; Degli-Esposti, D. New Challenges: Omics Technologies in Ecotoxicology. In Ecotoxicology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 181–208. [Google Scholar]
- Mauger, S.; Monard, C.; Thion, C.; Vandenkoornhuyse, P. Contribution of Single-Cell Omics to Microbial Ecology. Trends Ecol. Evol. 2021, 37, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Pandey, L.K.; Bergey, E.A. Exploring the Status of Motility, Lipid Bodies, Deformities and Size Reduction in Periphytic Diatom Community from Chronically Metal (Cu, Zn) Polluted Waterbodies as a Biomonitoring Tool. Sci. Total Environ. 2016, 550, 372–381. [Google Scholar]
- Arora, N.; Pienkos, P.T.; Pruthi, V.; Poluri, K.M.; Guarnieri, M.T. Leveraging Algal Omics to Reveal Potential Targets for Augmenting TAG Accumulation. Biotechnol. Adv. 2018, 36, 1274–1292. [Google Scholar] [CrossRef]
- Monroig, Ó.; Shu-Chien, A.C.; Kabeya, N.; Tocher, D.R.; Castro, L.F.C. Desaturases and Elongases Involved in Long-Chain Polyunsaturated Fatty Acid Biosynthesis in Aquatic Animals: From Genes to Functions. Prog. Lipid Res. 2022, 86, 101157. [Google Scholar] [CrossRef]
- Xie, D.; Chen, C.; Dong, Y.; You, C.; Wang, S.; Monroig, Ó.; Tocher, D.R.; Li, Y. Regulation of Long-Chain Polyunsaturated Fatty Acid Biosynthesis in Teleost Fish. Prog. Lipid Res. 2021, 82, 101095. [Google Scholar] [CrossRef]
- Castro, L.F.C.; Tocher, D.R.; Monroig, O. Long-Chain Polyunsaturated Fatty Acid Biosynthesis in Chordates: Insights into the Evolution of Fads and Elovl Gene Repertoire. Prog. Lipid Res. 2016, 62, 25–40. [Google Scholar] [CrossRef]
- Galachyants, Y.P.; Zakharova, Y.R.; Volokitina, N.A.; Morozov, A.A.; Likhoshway, Y.V.; Grachev, M.A. De Novo Transcriptome Assembly and Analysis of the Freshwater Araphid Diatom Fragilaria radians, Lake Baikal. Sci. Data 2019, 6, 183. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Bonaventura, G.D.; Hébraud, M.; Jaglic, Z. Critical Review on Biofilm Methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef]
- D’Alelio, D.; Russo, L.; Del Gaizo, G.; Caputi, L. Plankton under Pressure: How Water Conditions Alter the Phytoplankton–Zooplankton Link in Coastal Lagoons. Water 2022, 14, 974. [Google Scholar] [CrossRef]
- Delgado, M.L.; Ruzzante, D.E. Investigating Diadromy in Fishes and Its Loss in An-Omics Era. iScience 2020, 23, 101837. [Google Scholar] [CrossRef] [PubMed]
- Kienzler, A.; Connors, K.A.; Bonnell, M.; Barron, M.G.; Beasley, A.; Inglis, C.G.; Norberg-King, T.J.; Martin, T.; Sanderson, H.; Vallotton, N. Mode of Action Classifications in the EnviroTox Database: Development and Implementation of a Consensus MOA Classification. Environ. Toxicol. Chem. 2019, 38, 2294–2304. [Google Scholar] [CrossRef] [PubMed]
- Lyu, K.; Gu, L.; Wang, H.; Zhu, X.; Zhang, L.; Sun, Y.; Huang, Y.; Yang, Z. Transcriptomic Analysis Dissects the Mechanistic Insight into the Daphnia Clonal Variation in Tolerance to Toxic Microcystis. Limnol. Oceanogr. 2019, 64, 272–283. [Google Scholar] [CrossRef]
- Environmental Protection Agency. U.S. ECOTOX User Guide. ECOTOXicology Database System. 2022. Available online: https://nepis.epa.gov/Exe/ZyPDF.cgi/P10164D9.PDF?Dockey=P10164D9.PDF (accessed on 1 December 2022).
- Barron, M.G.; Lilavois, C.R.; Martin, T.M. MOAtox: A Comprehensive Mode of Action and Acute Aquatic Toxicity Database for Predictive Model Development. Aquat. Toxicol. 2015, 161, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Karpowicz, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I.; Dzialowski, A.R. Zooplankton Community Responses to Oxygen Stress. Water 2020, 12, 706. [Google Scholar] [CrossRef]
- Gamboa, M. Hemocyanin and Hexamerins Expression in Response to Hypoxia in Stoneflies (Plecoptera, Insecta). Arch. Insect. Biochem. Physiol. 2020, 105, e21743. [Google Scholar] [CrossRef]
- Maberly, S.C.; Gontero, B.; Puppo, C.; Villain, A.; Severi, I.; Giordano, M. Inorganic Carbon Uptake in a Freshwater Diatom, Asterionella formosa (Bacillariophyceae): From Ecology to Genomics. Phycologia 2021, 60, 427–438. [Google Scholar] [CrossRef]
- Weigand, H.; Weiss, M.; Cai, H.; Li, Y.; Yu, L.; Zhang, C.; Leese, F. Fishing in Troubled Waters: Revealing Genomic Signatures of Local Adaptation in Response to Freshwater Pollutants in Two Macroinvertebrates. Sci. Total Environ. 2018, 633, 875–891. [Google Scholar] [CrossRef]
- Hotaling, S.; Kelley, J.L.; Frandsen, P.B. Aquatic Insects Are Dramatically Underrepresented in Genomic Research. Insects 2020, 11, 601. [Google Scholar] [CrossRef]
- Begić, V.; Perić, M.S.; Hančić, S.; Štargl, M.; Svoboda, M.; Korać, P.; Radanović, I. Estimating Quality of Archive Urban Stream Macroinvertebrate Samples for Genomic, Transcriptomic and Proteomic Assessment. Ecol. Indic. 2021, 125, 107509. [Google Scholar] [CrossRef]
- Bista, I.; Carvalho, G.R.; Tang, M.; Walsh, K.; Zhou, X.; Hajibabaei, M.; Shokralla, S.; Seymour, M.; Bradley, D.; Liu, S. Performance of Amplicon and Shotgun Sequencing for Accurate Biomass Estimation in Invertebrate Community Samples. Mol. Ecol. Resour. 2018, 18, 1020–1034. [Google Scholar] [CrossRef]
- Smith, M.W.; Herfort, L.; Rivers, A.R.; Simon, H.M. Genomic Signatures for Sedimentary Microbial Utilization of Phytoplankton Detritus in a Fast-Flowing Estuary. Front. Microbiol. 2019, 10, 2475. [Google Scholar] [CrossRef]
- Porter, T.M.; Hajibabaei, M. Scaling up: A Guide to High-throughput Genomic Approaches for Biodiversity Analysis. Mol. Ecol. 2018, 27, 313–338. [Google Scholar] [CrossRef]
- Binelli, A.; Torre, C.D.; Magni, S.; Parolini, M. Does Zebra Mussel (Dreissena polymorpha) Represent the Freshwater Counterpart of Mytilus in Ecotoxicological Studies? A Critical Review. Environ. Pollut. 2015, 196, 386–403. [Google Scholar] [CrossRef]
- Guan, J.Y.; Zhang, Z.Y.; Cao, Y.R.; Xu, X.D.; Storey, K.B.; Yu, D.N.; Zhang, J.Y. The Complete Mitochondrial Genome of Choroterpes (Euthralus) yixingensis (Ephemeroptera: Leptophlebiidae) and its Mitochondrial Protein-Coding Gene Expression under Imidacloprid Stress. Gene 2021, 800, 145833. [Google Scholar] [CrossRef]
- Ma, X.; Dai, W.; Kang, J.; Yang, L.; He, S. Comprehensive Transcriptome Analysis of Six Catfish Species from an Altitude Gradient Reveals Adaptive Evolution in Tibetan Fishes. G3 Genes Genomes Genet. 2016, 6, 141–148. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, H.; Dong, Z.; Yang, X.; Lin, L.; Chen, J.; Yuan, M. Comparative Transcriptomic Analysis of Fireflies (Coleoptera: Lampyridae) to Explore the Molecular Adaptations to Fresh Water. Mol. Ecol. 2020, 29, 2676–2691. [Google Scholar] [CrossRef]
- Jeong, J.E.; Patnaik, B.B.; Kang, S.W.; Hwang, H.-J.; Park, S.Y.; Wang, T.H.; Park, E.B.; Lee, J.B.; Nam, M.-M.; Jo, Y.H. Characterization of Physa acuta Expressed Sequence Tags and Transcript Mining Following Cadmium Exposure. Genes Genom. 2015, 37, 1017–1025. [Google Scholar] [CrossRef]
- Qiao, F.; Lei, K.; Li, Z.; Wei, Z.; Liu, Q.; Yang, L.; He, J.; An, L.; Qi, H.; Cui, S. Transcriptomic Responses of the Freshwater Snail (Parafossarulus striatulus) Following Dietary Exposure to Cyanobacteria. Sci. Total Environ. 2018, 624, 153–161. [Google Scholar] [CrossRef]
- Miserazzi, A.; Perrigault, M.; Sow, M.; Gelber, C.; Ciret, P.; Lomenech, A.M.; Dalens, J.M.; Weber, C.; Floch, S.L.; Lacroix, C. Proteome Changes in Muscles, Ganglia, and Gills in Corbicula fluminea Clams Exposed to Crude Oil: Relationship with Behavioural Disturbances. Aquat. Toxicol. 2020, 223, 105482. [Google Scholar] [CrossRef] [PubMed]
- Shakya, M.; Silvester, E.; Rees, G.; Rajapaksha, K.H.; Faou, P.; Holland, A. Changes to the Amino Acid Profile and Proteome of the Tropical Freshwater Microalga Chlorella sp. in Response to Copper Stress. Ecotoxicol. Environ. Saf. 2022, 233, 113336. [Google Scholar] [CrossRef]
- Muhseen, Z.T.; Xiong, Q.; Chen, Z.; Ge, F. Proteomics Studies on Stress Responses in Diatoms. Proteomics 2015, 15, 3943–3953. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.R.; Pestana, J.L.T.; Soares, A.M.V.M.; Devreese, B.; Lemos, M.F.L. Chironomus riparius Proteome Responses to Spinosad Exposure. Toxics 2020, 8, 117. [Google Scholar] [CrossRef]
- Trotter, B.; Wilde, M.V.; Brehm, J.; Dafni, E.; Aliu, A.; Arnold, G.J.; Fröhlich, T.; Laforsch, C. Long-Term Exposure of Daphnia magna to Polystyrene Microplastic (PS-MP) Leads to Alterations of the Proteome, Morphology and Life-History. Sci. Total Environ. 2021, 795, 148822. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, D.; Almunia, C.; Cogne, Y.; Pible, O.; Degli-Esposti, D.; Salvador, A.; Cristobal, S.; Sheehan, D.; Chaumot, A.; Geffard, O. Ecotoxicoproteomics: A Decade of Progress in Our Understanding of Anthropogenic Impact on the Environment. J. Proteom. 2019, 198, 66–77. [Google Scholar] [CrossRef]
- Wagner, N.D.; Simpson, A.J.; Simpson, M.J. Metabolomic Responses to Sublethal Contaminant Exposure in Neonate and Adult Daphnia Magna. Environ. Toxicol. Chem. 2017, 36, 938–946. [Google Scholar] [CrossRef]
- Baratange, C.; Paris-Palacios, S.; Bonnard, I.; Delahaut, L.; Grandjean, D.; Wortham, L.; Sayen, S.; Gallorini, A.; Michel, J.; Renault, D. Metabolic, Cellular and Defense Responses to Single and Co-Exposure to Carbamazepine and Methylmercury in Dreissena polymorpha. Environ. Pollut. 2022, 300, 118933. [Google Scholar] [CrossRef]
- Yong, W.-K.; Sim, K.-S.; Poong, S.-W.; Wei, D.; Phang, S.-M.; Lim, P.-E. Interactive Effects of Temperature and Copper Toxicity on Photosynthetic Efficiency and Metabolic Plasticity in Scenedesmus quadricauda (Chlorophyceae). J. Appl. Phycol. 2018, 30, 3029–3041. [Google Scholar] [CrossRef]
- Jeong, K.-S.; Jeong, K.-Y.; Hong, Y.-S.; Kim, D.-K.; Oh, H.-J.; Chang, K.-H. Application of Nuclear Magnetic Resonance for Analyzing Metabolic Characteristics of Winter Diatom Blooms. J. Plankton. Res. 2020, 42, 31–39. [Google Scholar] [CrossRef]
- Previšić, A.; Rožman, M.; Mor, J.R.; Acuña, V.; Serra-Compte, A.; Petrović, M.; Sabater, S. Aquatic Macroinvertebrates under Stress: Bioaccumulation of Emerging Contaminants and Metabolomics Implications. Sci. Total Environ. 2019, 704, 143322. [Google Scholar] [CrossRef]
- Peng, J.; Guo, J.; Lei, Y.; Mo, J.; Sun, H.; Song, J. Integrative Analyses of Transcriptomics and Metabolomics in Raphidocelis subcapitata Treated with Clarithromycin. Chemosphere 2021, 266, 128933. [Google Scholar] [CrossRef]
- Spänig, S.; Eick, L.; Nuy, J.K.; Beisser, D.; Ip, M.; Heider, D.; Boenigk, J. A Multi-Omics Study on Quantifying Antimicrobial Resistance in European Freshwater Lakes. Environ. Int. 2021, 157, 106821. [Google Scholar] [CrossRef]
- Brockmeier, E.K.; Hodges, G.; Hutchinson, T.H.; Butler, E.; Hecker, M.; Tollefsen, K.E.; Garcia-Reyero, N.; Kille, P.; Becker, D.; Chipman, K. The Role of Omics in the Application of Adverse Outcome Pathways for Chemical Risk Assessment. Toxicol. Sci. 2017, 158, 252–262. [Google Scholar] [CrossRef]
- Dumas, T.; Courant, F.; Fenet, H.; Gomez, E. Environmental Metabolomics Promises and Achievements in the Field of Aquatic Ecotoxicology: Viewed through the Pharmaceutical Lens. Metabolites 2022, 12, 186. [Google Scholar] [CrossRef]
- Cosio, C. Inorganic Mercury and Methyl-Mercury Uptake and Effects in the Aquatic Plant Elodea nuttallii: A Review of Multi-Omic Data in the Field and in Controlled Conditions. Appl. Sci. 2020, 10, 1817. [Google Scholar] [CrossRef]
- Kirse, A.; Bourlat, S.J.; Langen, K.; Zapke, B.; Zizka, V.M.A. Comparison of Destructive and Nondestructive DNA Extraction Methods for the Metabarcoding of Arthropod Bulk Samples. Mol. Ecol. Resour. 2022, 23, 92–105. [Google Scholar] [CrossRef]
- Zoppi, J.; Guillaume, J.-F.; Neunlist, M.; Chaffron, S. MiBiOmics: An Interactive Web Application for Multi-Omics Data Exploration and Integration. BMC Bioinform. 2021, 22, 6. [Google Scholar] [CrossRef]
- Piña, B.; Barata, C. A Genomic and Ecotoxicological Perspective of DNA Array Studies in Aquatic Environmental Risk Assessment. Aquat. Toxicol. 2011, 105, 40–49. [Google Scholar] [CrossRef]
- Blackman, R.C.; Brantschen, J.; Walser, J.; Wüthrich, R.; Altermatt, F. Monitoring Invasive Alien Macroinvertebrate Species with Environmental DNA. River Res. Appl. 2022, 38, 1400–1412. [Google Scholar] [CrossRef]
- Mock, T.; Medlin, L.K. Genomics and Genetics of Diatoms. Adv. Bot. Res. 2012, 64, 245–284. [Google Scholar]
- Ardila-Garcia, A.M.; Gregory, T.R. An Exploration of Genome Size Diversity in Dragonflies and Damselflies (Insecta: Odonata). J. Zool. 2009, 278, 163–173. [Google Scholar] [CrossRef]
- Steinberg, C.E.W.; Stürzenbaum, S.R.; Menzel, R. Genes and Environment—Striking the Fine Balance between Sophisticated Biomonitoring and True Functional Environmental Genomics. Sci. Total Environ. 2008, 400, 142–161. [Google Scholar] [CrossRef] [PubMed]
- Becks, L.; Ellner, S.P.; Jones, L.E.; Hairston, N.G. The Functional Genomics of an Eco-Evolutionary Feedback Loop: Linking Gene Expression, Trait Evolution, and Community Dynamics. Ecol. Lett. 2012, 15, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Gomes-dos-Santos, A.; Lopes-Lima, M.; Machado, A.M.; Marcos Ramos, A.; Usié, A.; Bolotov, I.N.; Vikhrev, I.V.; Breton, S.; Castro, L.F.C.; da Fonseca, R.R.; et al. The Crown Pearl: A Draft Genome Assembly of the European Freshwater Pearl Mussel Margaritifera margaritifera (Linnaeus, 1758). DNA Res. 2021, 28, dsab002. [Google Scholar] [CrossRef]
- Hussin, N.; Azmir, I.A.; Esa, Y.; Ahmad, A.; Salleh, F.M.; Jahari, P.N.S.; Munian, K.; Gan, H.M. Characterization of the First Mitogenomes of the Smallest Fish in the World, Paedocypris progenetica, from Peat Swamp of Peninsular Malaysia, Selangor, and Perak. Genom. Inform. 2022, 20, e12. [Google Scholar] [CrossRef]
- Cheng, P.; Huang, Y.; Du, H.; Li, C.; Lv, Y.; Ruan, R.; Ye, H.; Bian, C.; You, X.; Xu, J.; et al. Draft Genome and Complete Hox-Cluster Characterization of the Sterlet (Acipenser ruthenus). Front. Genet. 2019, 10, 776. [Google Scholar] [CrossRef]
- Di Genova, A.; Nardocci, G.; Maldonado-Agurto, R.; Hodar, C.; Valdivieso, C.; Morales, P.; Gajardo, F.; Marina, R.; Gutiérrez, R.A.; Orellana, A.; et al. Genome Sequencing and Transcriptomic Analysis of the Andean Killifish Orestias ascotanensis Reveals Adaptation to High-Altitude Aquatic Life. Genomics 2022, 114, 305–315. [Google Scholar] [CrossRef]
- Alindonosi, A.; Baeshen, M.; Elsharawy, N. Prospects for Diatoms Identification Using Metagenomics: A Review. Appl. Ecol. Environ. Res. 2021, 19, 4281–4298. [Google Scholar] [CrossRef]
- Edge, T.A.; Baird, D.J.; Bilodeau, G.; Gagné, N.; Greer, C.; Konkin, D.; Newton, G.; Séguin, A.; Beaudette, L.; Bilkhu, S. The Ecobiomics Project: Advancing Metagenomics Assessment of Soil Health and Freshwater Quality in Canada. Sci. Total Environ. 2020, 710, 135906. [Google Scholar] [CrossRef]
- Vakati, V.; Dodsworth, S. Non-Destructive Genome Skimming for Aquatic Copepods. Conserv. Genet. Resour. 2020, 12, 515–520. [Google Scholar] [CrossRef]
- Tan, M.H.; Gan, H.M.; Bracken-Grissom, H.; Chan, T.Y.; Grandjean, F.; Austin, C.M. More from Less: Genome Skimming for Nuclear Markers for Animal Phylogenomics, a Case Study Using Decapod Crustaceans. J. Crustacean Biol. 2021, 41, ruab009. [Google Scholar] [CrossRef]
- Buchner, D.; Haase, P.; Leese, F. Wet Grinding of Invertebrate Bulk Samples—A Scalable and Cost-Efficient Protocol for Metabarcoding and Metagenomics. Metabarcoding Metagenom. 2021, 5, e67533. [Google Scholar]
- Mutz, K.O.; Heilkenbrinker, A.; Lönne, M.; Walter, J.G.; Stahl, F. Transcriptome Analysis Using Next-Generation Sequencing. Curr. Opin. Biotechnol. 2013, 24, 22–30. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Smith, A.M.; Ellison, A.M.; Ballif, B.A. Proteomic Characterization of the Major Arthropod Associates of the Carnivorous Pitcher Plant Sarracenia purpurea. Proteomics 2011, 11, 2354–2358. [Google Scholar] [CrossRef]
- Larras, F.; Regier, N.; Planchon, S.; Poté, J.; Renaut, J.; Cosio, C. Physiological and Proteomic Changes Suggest an Important Role of Cell Walls in the High Tolerance to Metals of Elodea nuttallii. J. Hazard Mater. 2013, 263, 575–583. [Google Scholar] [CrossRef]
- Büchel, C.; Wilhelm, C.; Wagner, V.; Mittag, M. Functional Proteomics of Light-Harvesting Complex Proteins under Varying Light-Conditions in Diatoms. J. Plant. Physiol. 2017, 217, 38–43. [Google Scholar] [CrossRef]
- Laetz, C.A.; Baldwin, D.H.; Hebert, V.R.; Stark, J.D.; Scholz, N.L. Elevated Temperatures Increase the Toxicity of Pesticide Mixtures to Juvenile Coho salmon. Aquat. Toxicol. 2014, 146, 38–44. [Google Scholar] [CrossRef]
- Gandar, A.; Laffaille, P.; Marty-Gasset, N.; Viala, D.; Molette, C.; Jean, S. Proteome Response of Fish under Multiple Stress Exposure: Effects of Pesticide Mixtures and Temperature Increase. Aquat. Toxicol. 2017, 184, 61–77. [Google Scholar] [CrossRef]
- Cappello, T. Environmental Metabolomics in Aquatic Pollution and Toxicology. J Aquat. Pollut. Toxicol. 2018, 2, 22. [Google Scholar]
- Taylor, N.S.; Gavin, A.; Viant, M.R. Metabolomics Discovers Early-Response Metabolic Biomarkers That Can Predict Chronic Reproductive Fitness in Individual Daphnia Magna. Metabolites 2018, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Vallejo, M.; García, A.; Barbas, C. Method Validation Strategies Involved in Non-Targeted Metabolomics. J. Chromatogr. A 2014, 1353, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Izral, N.M.; Brua, R.B.; Culp, J.M.; Yates, A.G. Crayfish Tissue Metabolomes Effectively Distinguish Impacts of Wastewater and Agriculture in Aquatic Ecosystems. Sci. Total Environ. 2021, 760, 143322. [Google Scholar] [CrossRef] [PubMed]
- Roszkowska, A.; Yu, M.; Bessonneau, V.; Ings, J.; McMaster, M.; Smith, R.; Bragg, L.; Servos, M.; Pawliszyn, J. In Vivo Solid-Phase Microextraction Sampling Combined with Metabolomics and Toxicological Studies for the Non-Lethal Monitoring of the Exposome in Fish Tissue. Environ. Pollut. 2019, 249, 109–115. [Google Scholar] [CrossRef]
- Beale, D.J.; Karpe, A.V.; Ahmed, W.; Cook, S.; Morrison, P.D.; Staley, C.; Sadowsky, M.J.; Palombo, E.A. A Community Multi-Omics Approach towards the Assessment of Surface Water Quality in an Urban River System. Int. J. Environ. Res. Public Health 2017, 14, 303. [Google Scholar] [CrossRef]
- Shah, R.M.; Stephenson, S.; Crosswell, J.; Gorman, D.; Hillyer, K.E.; Palombo, E.A.; Jones, O.A.H.; Cook, S.; Bodrossy, L.; van de Kamp, J. Omics-Based Ecosurveillance Uncovers the Influence of Estuarine Macrophytes on Sediment Microbial Function and Metabolic Redundancy in a Tropical Ecosystem. Sci. Total Environ. 2022, 809, 151175. [Google Scholar] [CrossRef]
- Zizka, V.M.A.; Leese, F.; Peinert, B.; Geiger, M.F. DNA Metabarcoding from Sample Fixative as a Quick and Voucher-Preserving Biodiversity Assessment Method. Genome 2019, 62, 122–136. [Google Scholar] [CrossRef]
- Rivera, S.F.; Vasselon, V.; Mary, N.; Monnier, O.; Rimet, F.; Bouchez, A. Exploring the Capacity of Aquatic Biofilms to Act as Environmental DNA Samplers: Test on Macroinvertebrate Communities in Rivers. Sci. Total Environ. 2021, 763, 144208. [Google Scholar] [CrossRef]
- Fernández, S.; Rodríguez, S.; Martínez, J.L.; Borrell, Y.J.; Ardura, A.; García-Vázquez, E. Evaluating Freshwater Macroinvertebrates from EDNA Metabarcoding: A River Nalón Case Study. PLoS ONE 2018, 13, e0201741. [Google Scholar] [CrossRef]
- Apothéloz-Perret-Gentil, L.; Bouchez, A.; Cordier, T.; Cordonier, A.; Guéguen, J.; Rimet, F.; Vasselon, V.; Pawlowski, J. Monitoring the Ecological Status of Rivers with Diatom EDNA Metabarcoding: A Comparison of Taxonomic Markers and Analytical Approaches for the Inference of a Molecular Diatom Index. In Proceedings of the Molecular Ecology; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2021; Volume 30, pp. 2959–2968. [Google Scholar]
- Bae, M.-J.; Ham, S.-N.; Lee, Y.-K.; Kim, E.-J. Evaluation of Benthic Macroinvertebrate Diversity in a Stream of Abandoned Mine Land Based on Environmental DNA (EDNA) Approach. Korean J. Ecol. Environ. 2021, 54, 221–228. [Google Scholar] [CrossRef]
- Weitemier, K.; Penaluna, B.E.; Hauck, L.L.; Longway, L.J.; Garcia, T.; Cronn, R. Estimating the Genetic Diversity of Pacific Salmon and Trout Using Multigene EDNA Metabarcoding. Mol. Ecol. 2021, 30, 4970–4990. [Google Scholar] [CrossRef] [PubMed]
- Hanžek, N.; Udovič, M.G.; Kajan, K.; Borics, G.; Várbíró, G.; Stoeck, T.; Žutinić, P.; Orlić, S.; Stanković, I. Assessing Ecological Status in Karstic Lakes through the Integration of Phytoplankton Functional Groups, Morphological Approach and Environmental DNA Metabarcoding. Ecol. Indic. 2021, 131, 108166. [Google Scholar] [CrossRef]
- Deiner, K.; Fronhofer, E.A.; Mächler, E.; Walser, J.C.; Altermatt, F. Environmental DNA Reveals That Rivers Are Conveyer Belts of Biodiversity Information. Nat. Commun. 2016, 7, 12544. [Google Scholar] [CrossRef] [PubMed]
- Poulíčková, A.; Dvořák, P.; Mazalová, P.; Hašler, P. Epipelic Microphototrophs: An Overlooked Assemblage in Lake Ecosystems. Freshw. Sci. 2014, 33, 513–523. [Google Scholar] [CrossRef]
- Beale, D.J.; Karpe, A.V.; Ahmed, W. Beyond Metabolomics: A Review of Multi-Omics-Based Approaches. In Microbial Metabolomics; Springer: Cham, Switzerland, 2016; pp. 289–312. [Google Scholar]
- Gotelli, N.J.; Ellison, A.M.; Ballif, B.A. Environmental Proteomics, Biodiversity Statistics and Food-Web Structure. Trends Ecol. Evol. 2012, 27, 436–442. [Google Scholar] [CrossRef]
- Markert, B.A.; Breure, A.M.; Zechmeister, H.G. Definitions, Strategies and Principles for Bioindication/Biomonitoring of the Environment. In Trace Metals and other Contaminants in the Environment; Elsevier: Amsterdam, The Netherlands, 2003; Volume 6, pp. 3–39. [Google Scholar]
- Le, Q.A.V.; Sekhon, S.S.; Lee, L.; Ko, J.H.; Min, J. Daphnia in Water Quality Biomonitoring—“Omic” Approaches. Toxicol. Environ. Health Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef]
- Ebner, J.N. Trends in the Application of “Omics” to Ecotoxicology and Stress Ecology. Genes 2021, 12, 1481. [Google Scholar] [CrossRef]
- Jamers, A.; Blust, R.; Coen, W. De Omics in Algae: Paving the Way for a Systems Biological Understanding of Algal Stress Phenomena? Aquat. Toxicol. 2009, 92, 114–121. [Google Scholar] [CrossRef]
- O’Callaghan, I.; Harrison, S.; Fitzpatrick, D.; Sullivan, T. The Freshwater Isopod Asellus aquaticus as a Model Biomonitor of Environmental Pollution: A Review. Chemosphere 2019, 235, 498–509. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An Emerging Tool in Conservation for Monitoring Past and Present Biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Pauls, S.U.; Alp, M.; Bálint, M.; Bernabò, P.; Čiampor, F., Jr.; Čiamporová-Zaťovičová, Z.; Finn, D.S.; Kohout, J.; Leese, F.; Lencioni, V.; et al. Integrating Molecular Tools into Freshwater Ecology: Developments and Opportunities. Freshw. Biol. 2014, 59, 1559–1576. [Google Scholar] [CrossRef]
- Ouyang, S.; Li, Y.; Zheng, T.; Wu, K.; Wang, X.; Zhou, Q. Ecotoxicity of Natural Nanocolloids in Aquatic Environment. Water 2022, 14, 2971. [Google Scholar] [CrossRef]
- Reitsema, R.E.; Meire, P.; Schoelynck, J. The Future of Freshwater Macrophytes in a Changing World: Dissolved Organic Carbon Quantity and Quality and Its Interactions with Macrophytes. Front. Plant Sci. 2018, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Chessman, B.C. Dissolved-Oxygen, Current and Temperature Preferences of Stream Invertebrates Estimated from Field Distributions: Application to Assemblage Responses to Drought. Hydrobiologia 2018, 809, 141–153. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Halliday, B.T.; Matthews, T.G.; Iervasi, D.; Dodemaide, D.T.; Pickett, P.J.; Linn, M.M.; Burns, A.; Bail, I.; Lester, R.E. Potential for Water-Resource Infrastructure to Act as Refuge Habitat. Ecol. Eng. 2015, 84, 136–148. [Google Scholar] [CrossRef]
- Sagrario, G.; Los Ángeles, M.D.; Balseiro, E.; Ituarte, R.; Spivak, E. Macrophytes as Refuge or Risky Area for Zooplankton: A Balance Set by Littoral Predacious Macroinvertebrates. Freshw. Biol. 2009, 54, 1042–1053. [Google Scholar] [CrossRef]
- Misteli, B.; Pannard, A.; Labat, F.; Fosso, L.K.; Baso, N.C.; Harpenslager, S.F.; Motitsoe, S.N.; Thiebaut, G.; Piscart, C. How Invasive Macrophytes Affect Macroinvertebrate Assemblages and Sampling Efficiency: Results from a Multinational Survey. Limnologica 2022, 96, 125998. [Google Scholar] [CrossRef]
- Wijewardene, L.; Wu, N.; Fohrer, N.; Riis, T. Epiphytic Biofilms in Freshwater and Interactions with Macrophytes: Current Understanding and Future Directions. Aquat. Bot. 2022, 176, 103467. [Google Scholar] [CrossRef]
- Tlili, A. Effects of Engineered Nanoparticles on Plant Litter Decomposition in Streams. In The Ecology of Plant Litter Decomposition in Stream Ecosystems; Springer: Cham, Switzerland, 2021; pp. 411–430. [Google Scholar]
- Pascoal, C.; Fernandes, I.; Seena, S.; Danger, M.; Ferreira, V.; Cássio, F. Linking Microbial Decomposer Diversity to Plant Litter Decomposition and Associated Processes in Streams. In The Ecology of Plant Litter Decomposition in Stream Ecosystems; Springer: Cham, Switzerland, 2021; pp. 163–192. [Google Scholar]
- Nakano, T.; Shirakawa, H.; Yeo, G.; Devlin, R.H.; Soga, T. Metabolome Profiling of Growth Hormone Transgenic Coho Salmon by Capillary Electrophoresis Time-of-Flight Mass Spectrometry. In Oceanography Challenges to Future Earth; Springer: Cham, Switzerland, 2019; pp. 223–234. [Google Scholar]
- Dunham, R.A.; Su, B. Genetically Engineered Fish: Potential Impacts on Aquaculture, Biodiversity, and the Environment. In GMOs; Springer: Cham, Switzerland, 2020; pp. 241–275. [Google Scholar]
- Beltran, J.F.; Zamorano, M.; Belén, L.H.; Risopatron, J.; Valdebenito, I.; Norambuena, J.-A.; Farias, J.A.; Farias, J.G. A Differential Proteomic Study Reveals the Downregulation of Several Plasma Membrane Ca2+-Binding Proteins in the Sperm of Atlantic Salmon (Salmo salar) Following Cold Storage. Aquaculture 2021, 545, 737211. [Google Scholar] [CrossRef]
- Luo, M.; Lu, G.; Yin, H.; Wang, L.; Atuganile, M.; Dong, Z. Fish Pigmentation and Coloration: Molecular Mechanisms and Aquaculture Perspectives. Rev. Aquac. 2021, 13, 2395–2412. [Google Scholar] [CrossRef]
- Arao, T.; Kato, Y.; Nong, Q.D.; Yamamoto, H.; Watanabe, H.; Matsuura, T.; Tatarazako, N.; Tani, K.; Okamoto, A.; Matsumoto, T. Production of Genome-Edited Daphnia for Heavy Metal Detection by Fluorescence. Sci. Rep. 2020, 10, 21490. [Google Scholar] [CrossRef] [PubMed]
- Trapp, J.; Armengaud, J.; Pible, O.; Gaillard, J.C.; Abbaci, K.; Habtoul, Y.; Chaumot, A.; Geffard, O. Proteomic Investigation of Male Gammarus fossarum, a Freshwater Crustacean, in Response to Endocrine Disruptors. J. Proteome Res. 2015, 14, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, J.D.; Gismondi, E. Behavioural and Biochemical Alterations in Gammarids as Induced by Chronic Metallic Exposures (Cd, Cu and Pb): Implications for Freshwater Biomonitoring. Chemosphere 2020, 257, 127253. [Google Scholar] [CrossRef] [PubMed]
- Sagova-Mareckova, M.; Boenigk, J.; Bouchez, A.; Cermakova, K.; Chonova, T.; Cordier, T.; Eisendle, U.; Elersek, T.; Fazi, S.; Fleituch, T. Expanding Ecological Assessment by Integrating Microorganisms into Routine Freshwater Biomonitoring. Water Res. 2021, 191, 116767. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.R.; Hawkins, C.P. Effects of Total Dissolved Solids on Growth and Mortality Predict Distributions of Stream Macroinvertebrates. Freshw. Biol. 2017, 62, 779–791. [Google Scholar] [CrossRef]
- El-Sheekh, M.; El-Dalatony, M.M.; Thakur, N.; Zheng, Y.; Salama, E.-S. Role of Microalgae and Cyanobacteria in Wastewater Treatment: Genetic Engineering and Omics Approaches. Int. J. Environ. Sci. Technol. 2022, 19, 2173–2194. [Google Scholar] [CrossRef]
- Liu, Y.; Lv, J.; Feng, J.; Liu, Q.; Nan, F.; Xie, S. Treatment of Real Aquaculture Wastewater from a Fishery Utilizing Phytoremediation with Microalgae. J. Chem. Technol. Biotechnol. 2019, 94, 900–910. [Google Scholar] [CrossRef]
- Agarwal, P.; Rani, R. Strategic Management of Contaminated Water Bodies: Omics, Genome-Editing and Other Recent Advances in Phytoremediation. Environ. Technol. Innov. 2022, 27, 102463. [Google Scholar] [CrossRef]
- Marella, T.K.; López-Pacheco, I.Y.; Parra-Saldívar, R.; Dixit, S.; Tiwari, A. Wealth from Waste: Diatoms as Tools for Phycoremediation of Wastewater and for Obtaining Value from the Biomass. Sci. Total Environ. 2020, 724, 137960. [Google Scholar] [CrossRef]
- Mali, P.; Esvelt, K.M.; Church, G.M. Cas9 as a Versatile Tool for Engineering Biology. Nat. Methods 2013, 10, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Durán-Vinet, B.; Araya-Castro, K.; Chao, T.C.; Wood, S.A.; Gallardo, V.; Godoy, K.; Abanto, M. Potential Applications of CRISPR/Cas for next-Generation Biomonitoring of Harmful Algae Blooms: A Review. Harmful Algae 2021, 103, 102027. [Google Scholar] [CrossRef] [PubMed]
- Schenekar, T. The Current State of EDNA Research in Freshwater Ecosystems: Are We Shifting from the Developmental Phase to Standard Application in Biomonitoring? Hydrobiologia 2022, 850, 1263–1282. [Google Scholar] [CrossRef]
- Sun, D.; Guo, Z.; Liu, Y.; Zhang, Y. Progress and Prospects of CRISPR/Cas Systems in Insects and Other Arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.C. Risks and Consequences of Hazard Agents to Human Health. In Toxic Chemical and Biological Agents; Springer: Dordrecht, The Netherlands, 2020; pp. 129–141. [Google Scholar]
- Beale, D.J.; Sinclair, G.; Shah, R.; Paten, A.; Kumar, A.; Long, S.M.; Vardy, S.; Jones, O.A.H. A Review of Omics-Based PFAS Exposure Studies Reveals Common Biochemical Response Pathways. Sci. Total Environ. 2022, 845, 157255. [Google Scholar] [CrossRef] [PubMed]
- Blackman, R.C.; Altermatt, F.; Foulquier, A.; Lefébure, T.; Gauthier, M.; Bouchez, A.; Stubbington, R.; Weigand, A.M.; Leese, F.; Datry, T. Unlocking Our Understanding of Intermittent Rivers and Ephemeral Streams with Genomic Tools. Front. Ecol. Environ. 2021, 19, 574–583. [Google Scholar] [CrossRef]
- Zanne, A.E.; Abarenkov, K.; Afkhami, M.E.; Aguilar-Trigueros, C.A.; Bates, S.; Bhatnagar, J.M.; Busby, P.E.; Christian, N.; Cornwell, W.K.; Crowther, T.W. Fungal Functional Ecology: Bringing a Trait-based Approach to Plant-associated Fungi. Biol. Rev. 2020, 95, 409–433. [Google Scholar] [CrossRef]
- Biderre-Petit, C.; Charvy, J.; Bronner, G.; Chauvet, M.; Debroas, D.; Gardon, H.; Hennequin, C.; Jouan-Dufournel, I.; Moné, A.; Monjot, A. FreshOmics: A Manually Curated and Standardized–Omics Database for Investigating Freshwater Microbiomes. Mol. Ecol. Resour. 2022, 23, 222–232. [Google Scholar] [CrossRef]
- Ravindran, S.P.; Lüneburg, J.; Gottschlich, L.; Tams, V.; Cordellier, M. Daphnia Stressor Database: Taking Advantage of a Decade of Daphnia ‘-Omics’ Data for Gene Annotation. Sci. Rep. 2019, 9, 11135. [Google Scholar]
- Zhou, P.; Wang, L.; Liu, H.; Li, C.; Li, Z.; Wang, J.; Tan, X. CyanoOmicsDB: An Integrated Omics Database for Functional Genomic Analysis of Cyanobacteria. Nucleic Acids Res. 2022, 50, D758–D764. [Google Scholar]
- Clark, D.R.; Ferguson, R.M.W.; Harris, D.N.; Nicholass, K.J.M.; Prentice, H.J.; Randall, K.C.; Randell, L.; Warren, S.L.; Dumbrell, A.J. Streams of Data from Drops of Water: 21st Century Molecular Microbial Ecology. Wiley Interdiscip. Rev. Water 2018, 5, e1280. [Google Scholar] [CrossRef]
- Yang, L.; Xu, Z.; Zeng, H.; Sun, N.; Wu, B.; Wang, C.; Bo, J.; Li, L.; Dong, Y.; He, S. FishDB: An Integrated Functional Genomics Database for Fishes. BMC Genom. 2020, 21, 801. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.E.; Kang, S.W.; Hwang, H.-J.; Park, S.Y.; Patnaik, B.B.; Kim, C.; Kim, S.; Nam, M.-M.; Lee, J.B.; Wang, T.H.; et al. Expressed Sequence Tag Analysis and Annotation of Genetic Information from the Freshwater Clam, Pisidium (Neopisidium) coreanum Endemic to Korea. Genes Genom. 2015, 37, 1041–1049. [Google Scholar] [CrossRef]
- Zhang, Y. Freshwater Biodiversity Conservation in China: Progress in the Yangtze River Basin. Aquat. Conserv. 2022, 32, 1565–1570. [Google Scholar]
- Nam, S.-E.; Bae, D.-Y.; Ki, J.-S.; Ahn, C.-Y.; Rhee, J.-S. The Importance of Multi-Omics Approaches for the Health Assessment of Freshwater Ecosystems. Mol. Cell Toxicol. 2022, 19, 3–11. [Google Scholar] [CrossRef]
- Gubelit, Y.I.; Grossart, H.P. New Methods, New Concepts: What Can Be Applied to Freshwater Periphyton? Front. Microbiol. 2020, 11, 1275. [Google Scholar] [CrossRef]
- Tcherkez, G.; Schaeufele, R.; Nogues, S.; Piel, C.; Boom, A.; Lanigan, G.; Barbaroux, C.; Mata, C.; Elhani, S.; Hemming, D. On the 13C/12C Isotopic Signal of Day and Night Respiration at the Mesocosm Level. Plant Cell Environ. 2010, 33, 900–913. [Google Scholar] [CrossRef]
- Cortés-Guzmán, D.; Alcocer, J.; Planas, D. Autotrophs Are Important Contributors to Benthic Macroinvertebrate Food Webs in Two Tropical First-order Forest Streams. Freshw. Biol. 2022, 67, 941–953. [Google Scholar] [CrossRef]
- McLean, T.I. “Eco-Omics”: A Review of the Application of Genomics, Transcriptomics, and Proteomics for the Study of the Ecology of Harmful Algae. Microb. Ecol. 2013, 65, 901–915. [Google Scholar]
- Reel, P.S.; Reel, S.; Pearson, E.; Trucco, E.; Jefferson, E. Using Machine Learning Approaches for Multi-Omics Data Analysis: A Review. Biotechnol. Adv. 2021, 49, 107739. [Google Scholar] [CrossRef]
- Michel, J.P.; Soto, J.O. Modern Approaches into Biochemical and Molecular Biomarkers: Key Roles in Environmental Biotechnology. J. Biotechnol. Biomater. 2016, 6, 2. [Google Scholar] [CrossRef]
- Ritchie, M.D.; Holzinger, E.R.; Li, R.; Pendergrass, S.A.; Kim, D. Methods of Integrating Data to Uncover Genotype–Phenotype Interactions. Nat. Rev. Genet. 2015, 16, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tan, N.G.J.; Fu, B.; Li, S.F.Y. Metallomics and NMR-Based Metabolomics of Chlorella sp. Reveal the Synergistic Role of Copper and Cadmium in Multi-Metal Toxicity and Oxidative Stress. Metallomics 2015, 7, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhou, Q.; Hu, X. Integrating Multi-Omics and Regular Analyses Identifies the Molecular Responses of Zebrafish Brains to Graphene Oxide: Perspectives in Environmental Criteria. Ecotoxicol. Environ. Saf. 2019, 180, 269–279. [Google Scholar] [CrossRef]
- Sundaray, J.K.; Dixit, S.; Rather, A.; Rasal, K.D.; Sahoo, L. Aquaculture Omics: An Update on the Current Status of Research and Data Analysis. Mar. Genom. 2022, 64, 100967. [Google Scholar] [CrossRef] [PubMed]
- Creusot, N.; Chaumet, B.; Eon, M.; Mazzella, N.; Moreira, A.; Morin, S. Metabolomics Insight into the Influence of Environmental Factors in Responses of Freshwater Biofilms to the Model Herbicide Diuron. Environ. Sci. Pollut. Res. 2022, 29, 29332–29347. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Singh, A.; Chana, I. Computational Techniques and Tools for Omics Data Analysis: State-of-the-Art, Challenges, and Future Directions. Arch. Comput. Methods Eng. 2021, 28, 4595–4631. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. MixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]


{kind=link}
| Technique | BQEs or Other as Example | Key Finding | Advantages | Disadvantages | References |
|---|---|---|---|---|---|
| Genomics | Diatoms | Seasonal carbon dynamics studies using photosynthesis rate. | Comparative genomics in ecotoxicology has identified new molecular biomarkers in species that have not yet been defined. | Lack of data on freshwater organisms in digital repositories, limiting progress in the field. | [76,77,78,79] |
| Fish | Gene expression with many chemical/pharmaceutical stressors. | ||||
| Metagenomics | Macroinvertebrates | Study of individual biomass in communities. | Provide an overview of metabolic potential and reconstruct whole genome sequencing to understand taxa with small genomes. | Pathway genes or markers have a low frequency of recovery. Taxonomic assignments based on markers are rare. Large number of unknown genes. | [80,81,82] |
| Diatoms | Estimation of biogeochemical processes in estuaries. | ||||
| Transcriptomics | Dreissena polymorpha | Ecotoxicological bioassessment of chemicals and insecticides. | Transcriptomic study is broad, and it is possible to study changes in environmental conditions, species adaptations, biomarkers, and alterations in metabolic pathways. | Transcriptomics can be associated with certain protein expression analyses that originate from oxygen related molecular responses. Lack of knowledge in non-coding RNAs. | [44,75,83,84,85,86,87,88] |
| Catfish | Adaptation in high altitudes environments and their implications. | ||||
| Aquatic insects | Life cycle adaptations and adaptative evolution in aquatic insects. | ||||
| Physa acuta and Chlamydomonas (Green algae) | Search of new biomarkers by expressed sequence typing | ||||
| Proteomics | Chlorella algae | Variation in trophic chain due to exposure to metals. | More accurate in recognizing the effect of responses due to environmental conditions and selective pressures. | Requires the construction of more suitable databases for protein identification and pathway analysis in non-model species. | [33,89,90,91,92,93,94] |
| Aquatic insect (Chironomidae) | Impact of natural insecticides on increased globin protein production. | ||||
| Daphnia | Stress caused by temperature changes and exposure to microplastics that affect protein production. | ||||
| Metabolomics | Daphnia | Metabolite determination versus sublethal contaminant exposure. | It is capable to estimate biochemical metabolic changes by low molecular weight metabolites that represent the most functional measure of an organism’s physiology and response to toxic stress. | It is a technique rarely used in biomonitoring practices. | [46,95,96,97,98,99] |
| Scenedesmus obliquus | Stress response in fullerenes conditions. | ||||
| Dreissena polymorpha | Micropollutants effects (carbamazepina and methylmercury) | ||||
| Chloromonas augustae | Interactive effects of stressors (Cu and heating) | ||||
| Stephanodiscus hantzchii | Effect of temperature on internal metabolism. | ||||
| Trichoptera larvae | Chemical contamination effect. | ||||
| Multi-omics | Algae | Metabolomic-transcriptomic, to measure the effect of the macrolide antibiotic clarithromycin on the reactions. | Can provide more information about mechanisms of action by which contaminants achieved adverse outcomes at higher levels of biological organization. | It is necessary to have an integrated platform that guarantees the correct analysis of the data obtained from the different omics. | [100,101,102,103,104,105,106] |
| Microbial communities | Genomics-metagenomics, resistome exploration of the community | ||||
| Elodea nuttallii | Transcriptomic-proteomic-metabolomic, asses the negative impact of methyl-Hg and inorganic Hg in food chains |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machuca-Sepúlveda, J.; Miranda, J.; Lefin, N.; Pedroso, A.; Beltrán, J.F.; Farias, J.G. Current Status of Omics in Biological Quality Elements for Freshwater Biomonitoring. Biology 2023, 12, 923. https://doi.org/10.3390/biology12070923
Machuca-Sepúlveda J, Miranda J, Lefin N, Pedroso A, Beltrán JF, Farias JG. Current Status of Omics in Biological Quality Elements for Freshwater Biomonitoring. Biology. 2023; 12(7):923. https://doi.org/10.3390/biology12070923
Chicago/Turabian StyleMachuca-Sepúlveda, Jorge, Javiera Miranda, Nicolás Lefin, Alejandro Pedroso, Jorge F. Beltrán, and Jorge G. Farias. 2023. "Current Status of Omics in Biological Quality Elements for Freshwater Biomonitoring" Biology 12, no. 7: 923. https://doi.org/10.3390/biology12070923
APA StyleMachuca-Sepúlveda, J., Miranda, J., Lefin, N., Pedroso, A., Beltrán, J. F., & Farias, J. G. (2023). Current Status of Omics in Biological Quality Elements for Freshwater Biomonitoring. Biology, 12(7), 923. https://doi.org/10.3390/biology12070923