
The Interplay between Atherosclerosis and Cancer: Breast Cancer Cells Increase the Expression of Endothelial Cell Adhesion Markers
 ,
, 
Abstract
Simple Summary
Abstract
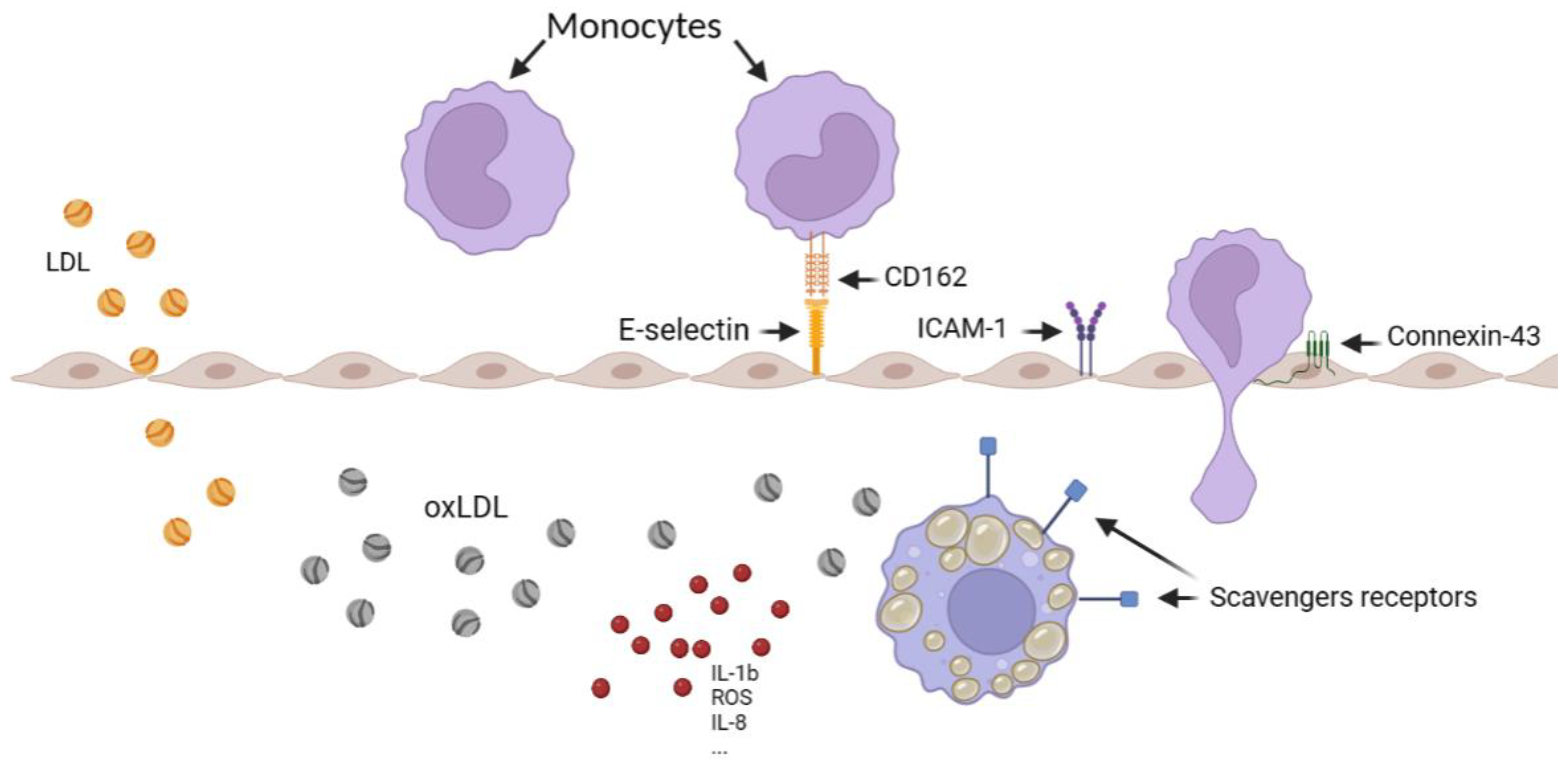
1. Introduction
2. Materials and Methods
2.1. oxLDL Production
2.2. Cell Culture
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Immunofluorescence Microscopy on Human Umbilical Vein Endothelial Cells (HUVECs)
2.5. Immunofluorescence Microscopy on Human Leukaemia Monocytic Cell Line (THP-1)
2.6. Statistical Analysis
3. Results
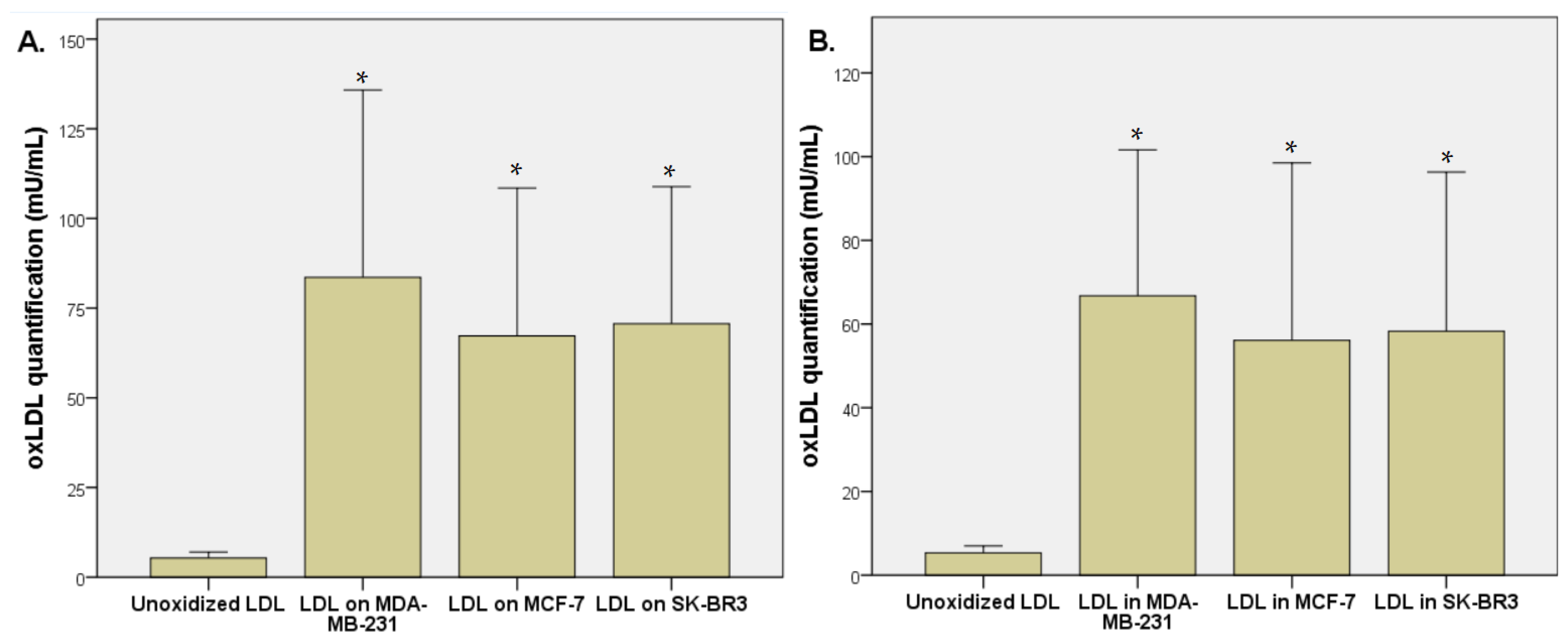
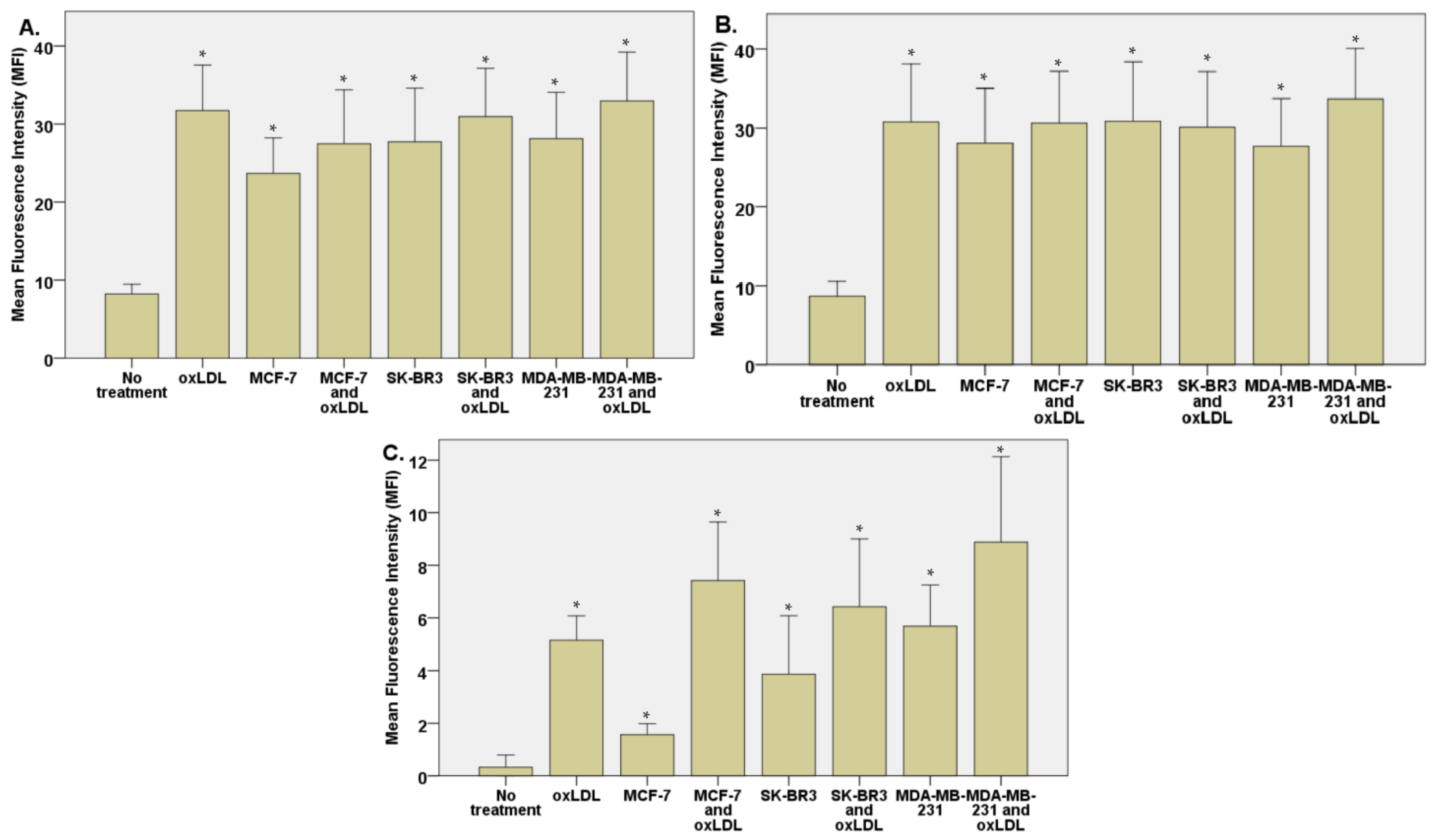
3.1. LDL Oxidation under Breast Cancer Cell Exposition
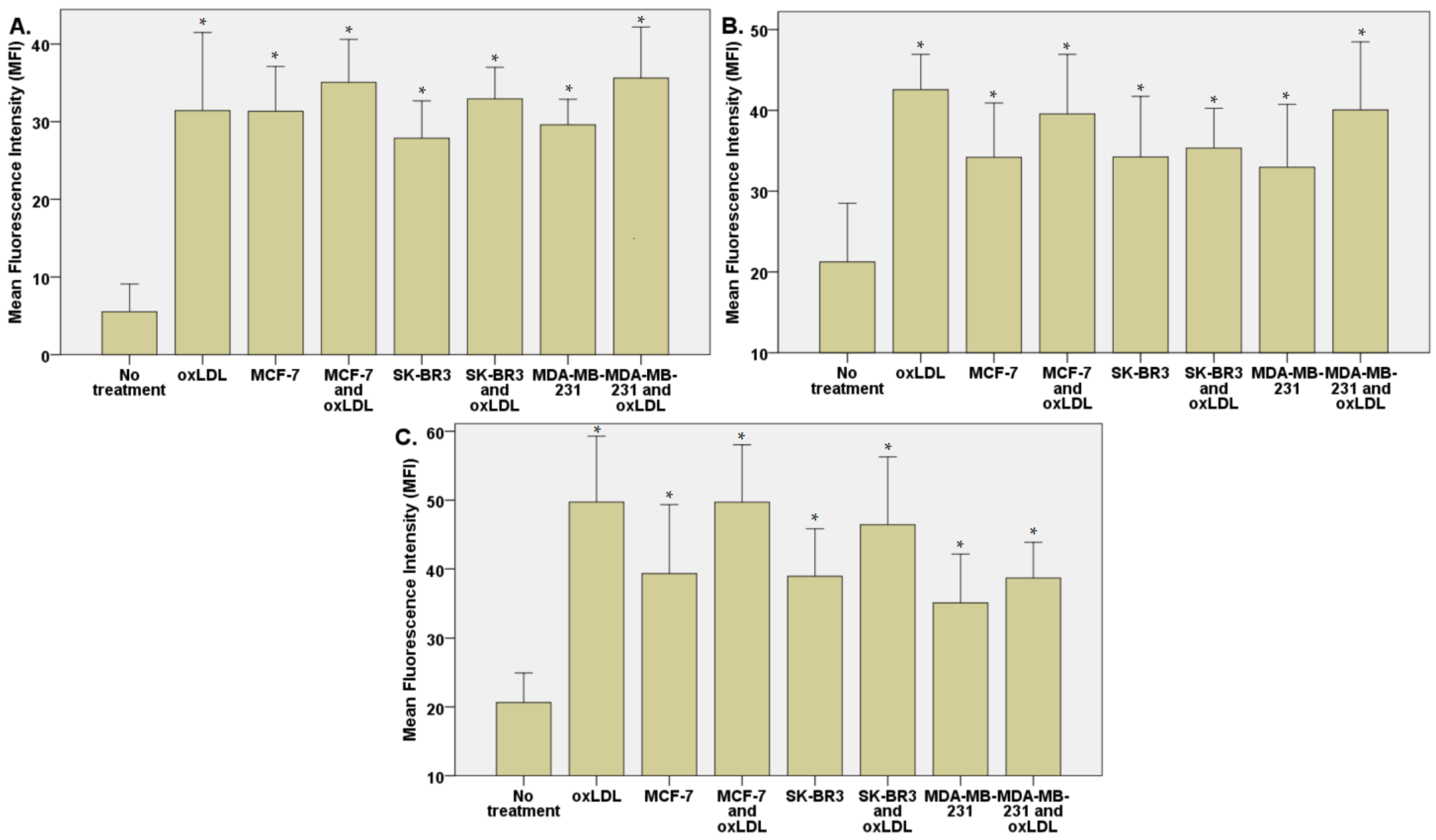
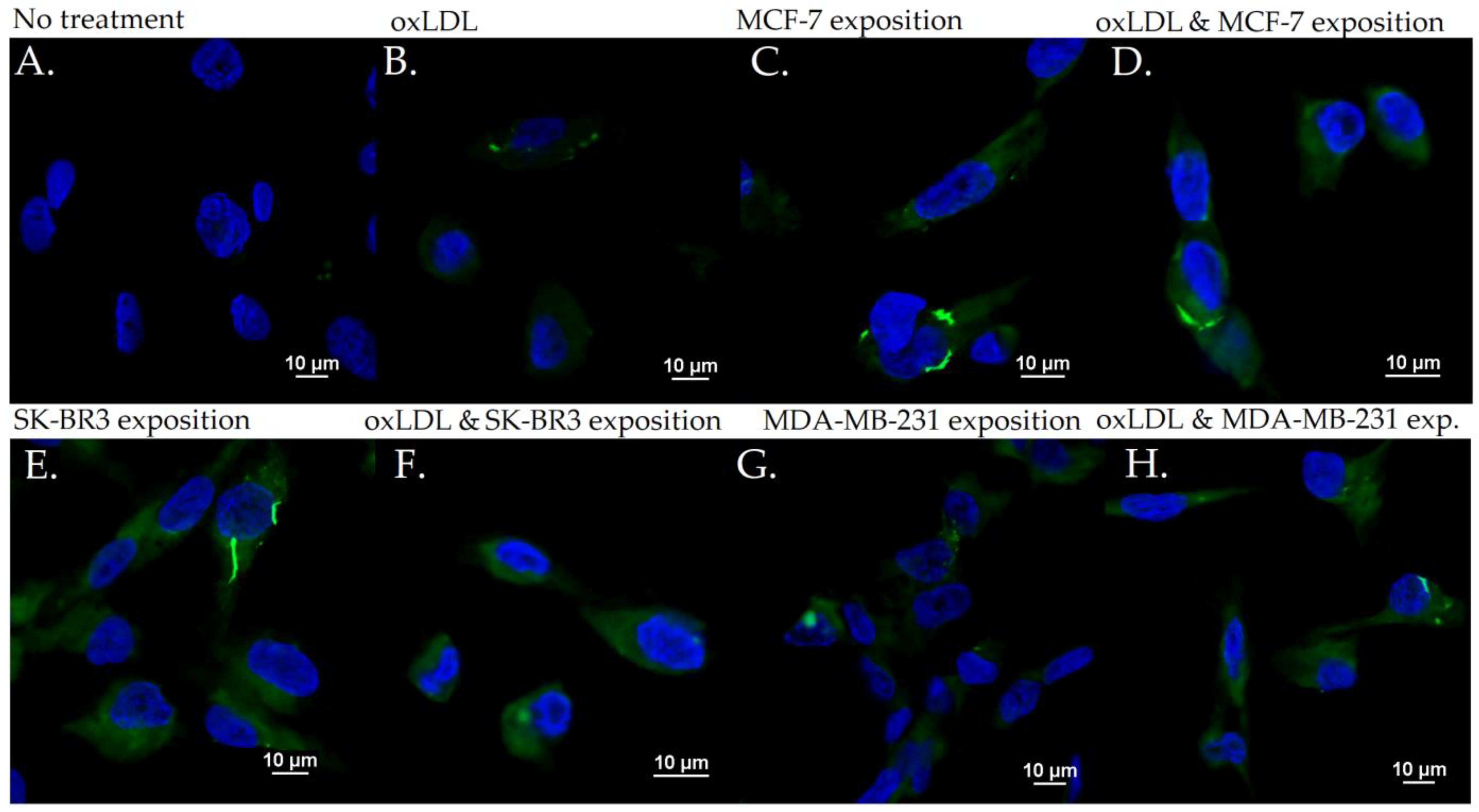
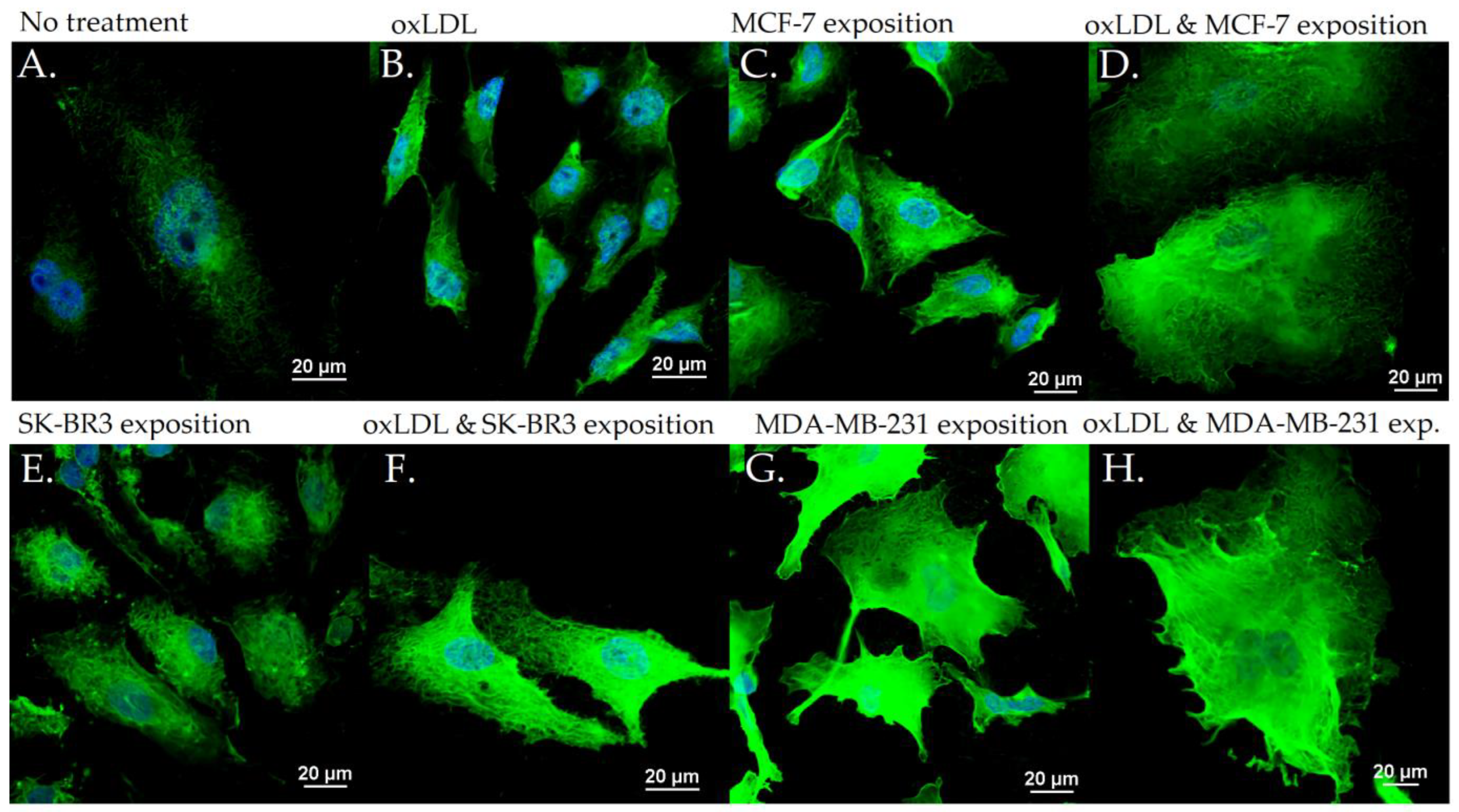
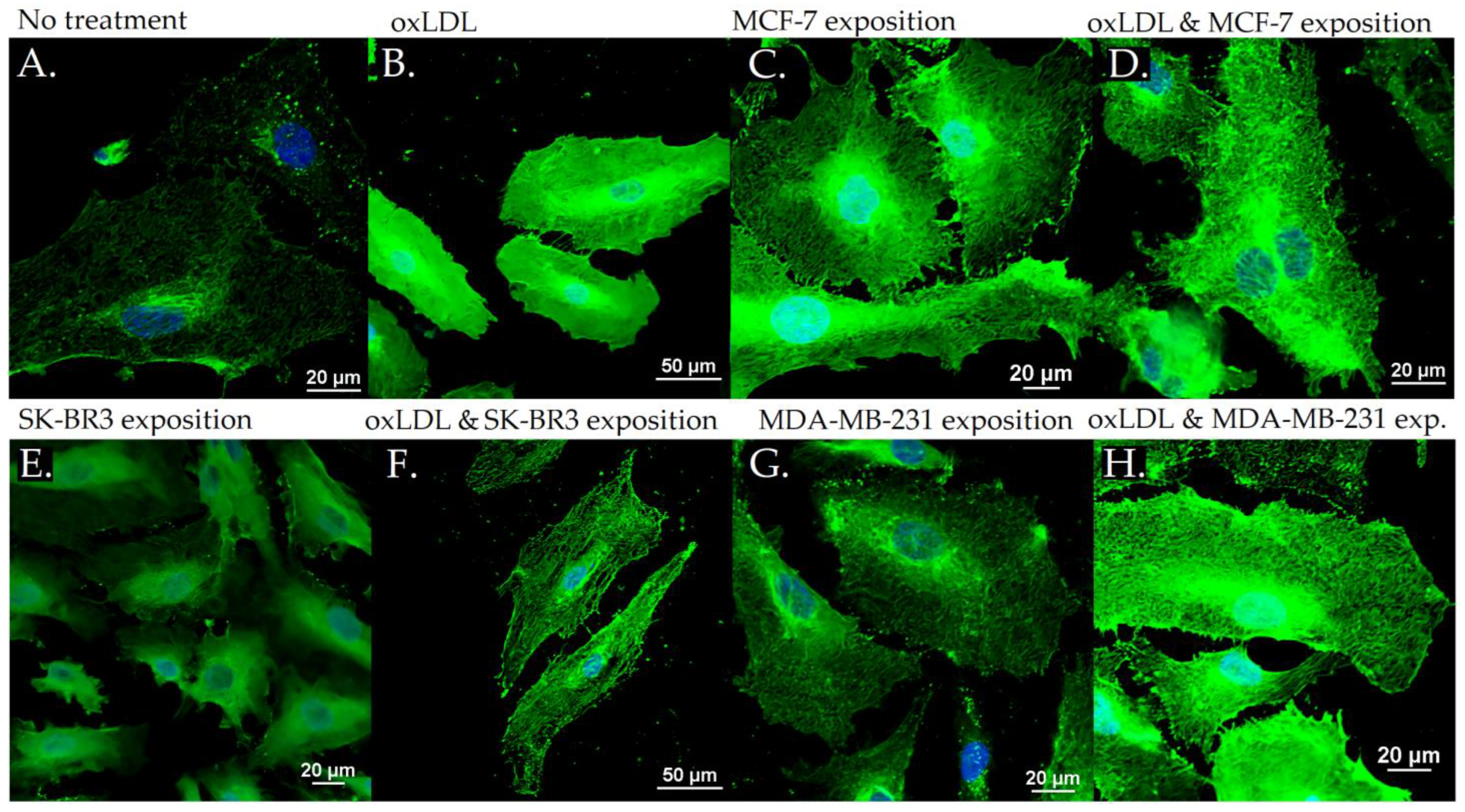
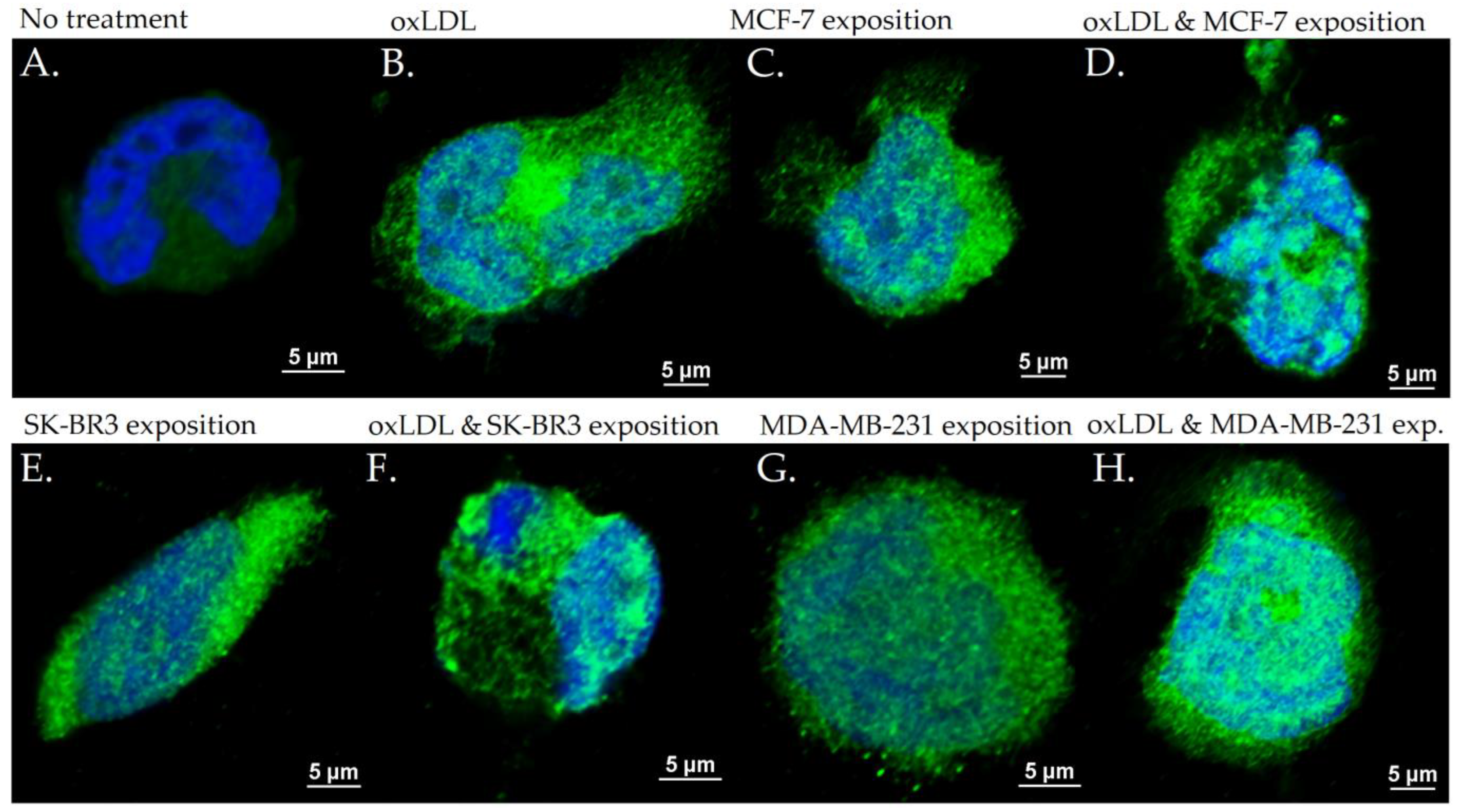
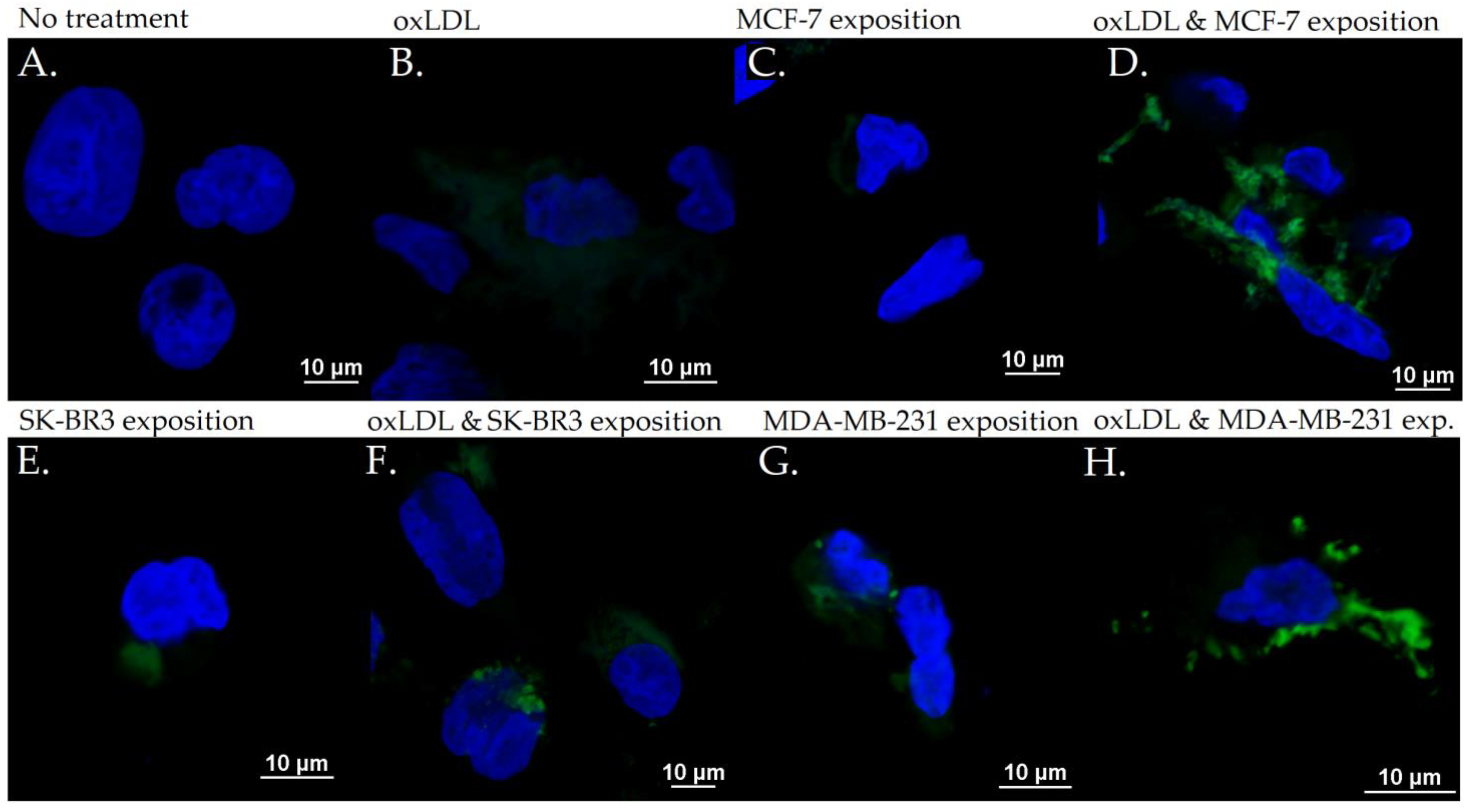
3.2. Endothelial Adhesion Markers in HUVEC upon Breast Cancer Environment Exposure
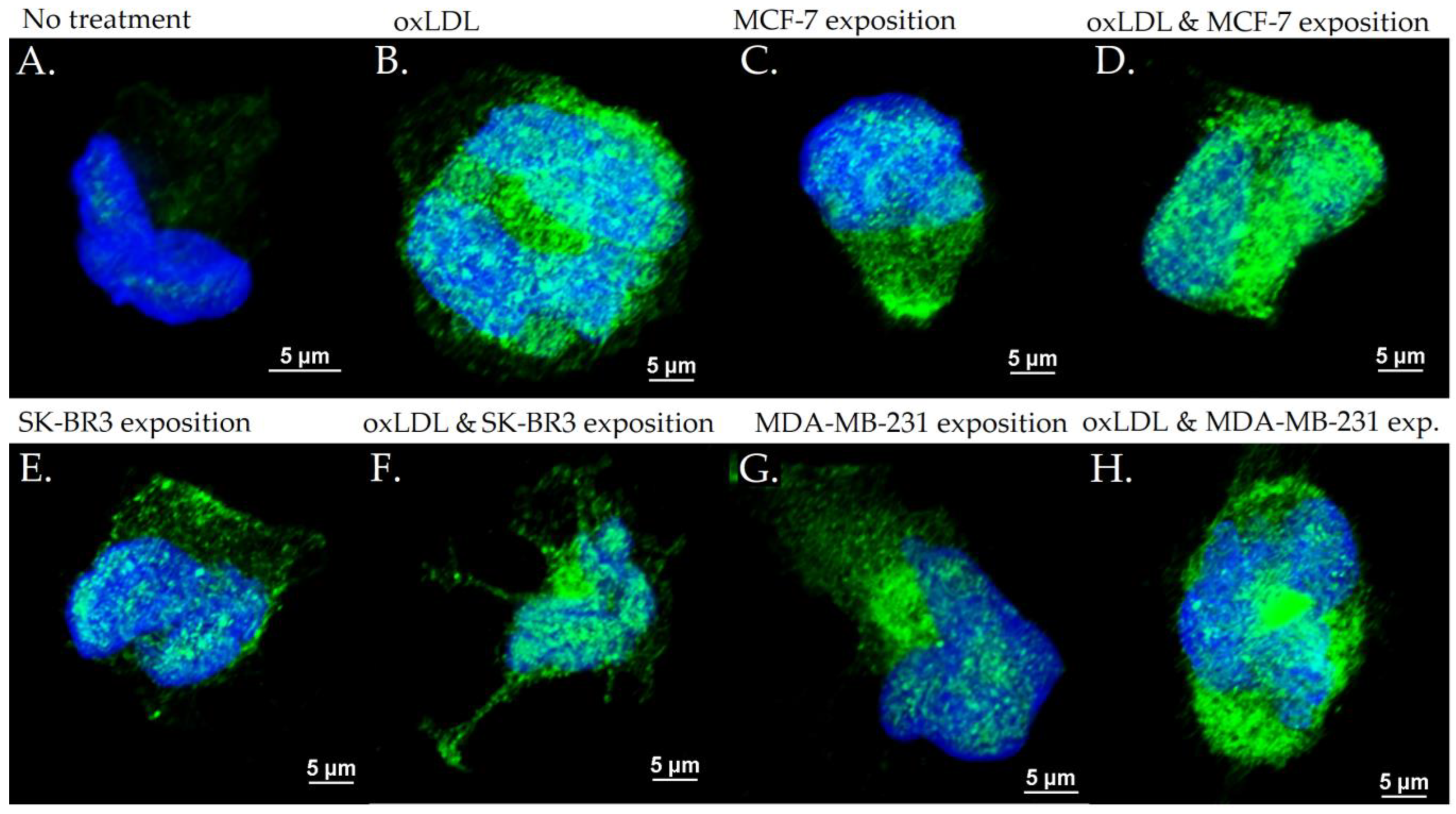
3.3. Scavenger Receptors and Adhesive Marker in THP-1 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cardiovascular Diseases. Available online: https://www.who.int/health-topics/cardiovascular-diseases (accessed on 9 November 2022).
- Cancer. Available online: http://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 18 May 2022).
- Dafni, U.; Tsourti, Z.; Alatsathianos, I. Breast Cancer Statistics in the European Union: Incidence and Survival across European Countries. Breast Care 2019, 14, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Dyba, T.; Randi, G.; Bettio, M.; Gavin, A.; Visser, O.; Bray, F. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries and 25 major cancers in 2018. Eur. J. Cancer 2018, 103, 356–387. [Google Scholar] [CrossRef] [PubMed]
- Carioli, G.; Malvezzi, M.; Rodriguez, T.; Bertuccio, P.; Negri, E.; La Vecchia, C. Trends and predictions to 2020 in breast cancer mortality in Europe. Breast 2017, 36, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Roubín, S.R.; Cordero, A. The Two-way Relationship Between Cancer and Atherosclerosis. Rev. Española Cardiol. (Engl. Ed.) 2019, 72, 487–494. [Google Scholar] [CrossRef]
- Florido, R.; Daya, N.R.; Ndumele, C.E.; Koton, S.; Russell, S.D.; Prizment, A.; Blumenthal, R.S.; Matsushita, K.; Mok, Y.; Felix, A.S.; et al. Cardiovascular Disease Risk Among Cancer Survivors. J. Am. Coll. Cardiol. 2022, 80, 22–32. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Rafieian-Kopaei, M.; Setorki, M.; Doudi, M.; Baradaran, A.; Nasri, H. Atherosclerosis: Process, Indicators, Risk Factors and New Hopes. Int. J. Prev. Med. 2014, 5, 927–946. [Google Scholar]
- Tapia-Vieyra, J.V.; Delgado-Coello, B.; Mas-Oliva, J. Atherosclerosis and Cancer; A Resemblance with Far-reaching Implications. Arch. Med Res. 2017, 48, 12–26. [Google Scholar] [CrossRef]
- Murdocca, M.; De Masi, C.; Pucci, S.; Mango, R.; Novelli, G.; Di Natale, C.; Sangiuolo, F. LOX-1 and cancer: An indissoluble liaison. Cancer Gene Ther. 2021, 28, 1088–1098. [Google Scholar] [CrossRef]
- Galkina, E.; Ley, K. Vascular Adhesion Molecules in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2292–2301. [Google Scholar] [CrossRef]
- Woollard, K.J.; Geissmann, F. Monocytes in atherosclerosis: Subsets and functions. Nat. Rev. Cardiol. 2010, 7, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Hidalgo, A.; Furie, B.C.; Vestweber, D.; Furie, B.; Frenette, P.S. PSGL-1 participates in E-selectin–mediated progenitor homing to bone marrow: Evidence for cooperation between E-selectin ligands and α4 integrin. Blood 2003, 102, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Faxon, D.P.; Fuster, V.; Libby, P.; Beckman, J.A.; Hiatt, W.R.; Thompson, R.W.; Topper, J.N.; Annex, B.H.; Rundback, J.H.; Fabunmi, R.P.; et al. Atherosclerotic Vascular Disease Conference. Circulation 2004, 109, 2617–2625. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Neyen, C.; Gordon, S. Role of macrophage scavenger receptors in atherosclerosis. Immunobiology 2012, 217, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Wingo, P.A.; Ries, L.A.G.; Rosenberg, H.M.; Miller, D.S.; Edwards, B.K. Cancer incidence and mortality, 1973–1995. Cancer 1998, 82, 1197–1207. [Google Scholar] [CrossRef]
- Rotheneder, M.; Kostner, G.M. Effects of low- and high-density lipoproteins on the proliferation of human breast cancer cellsIn vitro: Differences between hormone-dependent and hormone-independent cell lines. Int. J. Cancer 1989, 43, 875–879. [Google Scholar] [CrossRef]
- Dos Santos, C.R.; Fonseca, I.; Dias, S.; De Almeida, J.C.M. Plasma level of LDL-cholesterol at diagnosis is a predictor factor of breast tumor progression. BMC Cancer 2014, 14, 132. [Google Scholar] [CrossRef]
- Khaidakov, M.; Mehta, J.L. Oxidized LDL Triggers Pro-Oncogenic Signaling in Human Breast Mammary Epithelial Cells Partly via Stimulation of MiR-21. PLoS ONE 2012, 7, e46973. [Google Scholar] [CrossRef]
- Pucci, S.; Polidoro, C.; Greggi, C.; Amati, F.; Morini, E.; Murdocca, M.; Biancolella, M.; Orlandi, A.; Sangiuolo, F.; Novelli, G. Pro-oncogenic action of LOX-1 and its splice variant LOX-1Δ4 in breast cancer phenotypes. Cell Death Dis. 2019, 10, 53. [Google Scholar] [CrossRef]
- Klooster, C.C.V.; Ridker, P.M.; Hjortnaes, J.; van der Graaf, Y.; Asselbergs, F.W.; Westerink, J.; Aerts, J.G.J.V.; Visseren, F.L.J. The relation between systemic inflammation and incident cancer in patients with stable cardiovascular disease: A cohort study. Eur. Heart J. 2019, 40, 3901–3909. [Google Scholar] [CrossRef]
- Raza, U.; Asif, M.R.; Bin Rehman, A.; Sheikh, A. Hyperlipidemia and hyper glycaemia in Breast Cancer Patients is related to disease stage. Pak. J. Med. Sci. 2018, 34, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Pope, A.; Thomson, L.; Cantu, S.; Setia, G.; Torosyan, N.; Merz, N.B.; Atkins, K.; Anderson, E.M.; Cheng, S.; Tamarappoo, B. Detection of subclinical atherosclerosis from PET-CT in patients with breast cancer. J. Cardiovasc. Comput. Tomogr. 2022, 16, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Cauley, J.A.; Zmuda, J.M.; Lui, L.-Y.; Hillier, T.A.; Ness, R.B.; Stone, K.L.; Cummings, S.R.; Bauer, D.C. Lipid-lowering drug use and breast cancer in older women: A prospective study. J. Women’s Health 2003, 12, 749–756. [Google Scholar] [CrossRef]
- Bonovas, S.; Filioussi, K.; Tsavaris, N.; Sitaras, N.M. Use of statins and breast cancer: A meta-analysis of seven randomized clinical trials and nine observational studies. J. Clin. Oncol. 2005, 23, 8606–8612. [Google Scholar] [CrossRef] [PubMed]
- Scalia, A.; Kindt, N.; Trelcat, A.; Nachtergael, A.; Duez, P.; Journé, F.; Carlier, S. Development of a Method for Producing oxLDL: Characterization of Their Effects on HPV-Positive Head and Neck Cancer Cells. Int. J. Mol. Sci. 2022, 23, 12552. [Google Scholar] [CrossRef]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef]
- Rathod, K.S.; Siddiqui, U.; Hartley, A.; Khambata, R.; Ahluwalia, A. Investigation of the influence of sex on cantharidin-induced inflammation in healthy volunteers. FASEB J. 2016, 30, 716.9. [Google Scholar] [CrossRef]
- Li, D.; Mehta, J.L. Intracellular signaling of LOX-1 in endothelial cell apoptosis. Circ. Res. 2009, 104, 566–568. [Google Scholar] [CrossRef]
- Schaeffer, D.F.; Riazy, M.; Parhar, K.S.; Chen, J.H.; Duronio, V.; Sawamura, T.; Steinbrecher, U.P. LOX-1 augments oxLDL uptake by lysoPC-stimulated murine macrophages but is not required for oxLDL clearance from plasma. J. Lipid Res. 2009, 50, 1676–1684. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, X.-H.; Jin, L. Macrophage Polarization in Physiological and Pathological Pregnancy. Front. Immunol. 2019, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Stöger, J.L.; Gijbels, M.J.; van der Velden, S.; Manca, M.; van der Loos, C.M.; Biessen, E.A.; Daemen, M.J.; Lutgens, E.; de Winther, M.P. Distribution of macrophage polarization markers in human atherosclerosis. Atherosclerosis 2012, 225, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, L.S.; Mogensen, C.K.; Rosendahl, A.; Cucak, H.; Nielsen, L.B.; Rasmussen, S.E.; Pedersen, T.X. Bone marrow-derived and peritoneal macrophages have different inflammatory response to oxLDL and M1/M2 marker expression—Implications for atherosclerosis research. Sci. Rep. 2016, 6, 35234. [Google Scholar] [CrossRef] [PubMed]
- Gleissner, C.A.; Shaked, I.; Little, K.M.; Ley, K. CXC Chemokine Ligand 4 Induces a Unique Transcriptome in Monocyte-Derived Macrophages. J. Immunol. 2010, 184, 4810–4818. [Google Scholar] [CrossRef] [PubMed]
- Weagel, E.; Smith, C.; Liu, P.G.; Robison, R.; O’neill, K. Macrophage Polarization and Its Role in Cancer. J. Clin. Cell. Immunol. 2015, 6, 338. [Google Scholar] [CrossRef]
- Guiducci, C.; Vicari, A.P.; Sangaletti, S.; Trinchieri, G.; Colombo, M.P. Redirecting in vivo elicited tumor infiltrating macrophages and dendritic cells towards tumor rejection. Cancer Res. 2005, 65, 3437–3446. [Google Scholar] [CrossRef]
- Pe, K.C.S.; Saetung, R.; Yodsurang, V.; Chaotham, C.; Suppipat, K.; Chanvorachote, P.; Tawinwung, S. Triple-negative breast cancer influences a mixed M1/M2 macrophage phenotype associated with tumor aggressiveness. PLoS ONE 2022, 17, e0273044. [Google Scholar] [CrossRef]
- Nguyen, L.T.H.; Muktabar, A.; Tang, J.; Wong, Y.S.; Thaxton, C.S.; Venkatraman, S.S.; Ng, K.W. The Potential of Fluocinolone Acetonide to Mitigate Inflammation and Lipid Accumulation in 2D and 3D Foam Cell Cultures. BioMed Res. Int. 2018, 2018, 3739251. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Millican, R.; Lynd, T.; Gangasani, M.; Malhotra, S.; Sherwood, J.; Hwang, P.T.; Cho, Y.; Brott, B.C.; et al. Recent Progress in in vitro Models for Atherosclerosis Studies. Front. Cardiovasc. Med. 2022, 8, 790529. [Google Scholar] [CrossRef]
- Chen, C.-N.; Chang, S.-F.; Lee, P.-L.; Chang, K.; Chen, L.-J.; Usami, S.; Chien, S.; Chiu, J.-J. Neutrophils, lymphocytes, and monocytes exhibit diverse behaviors in transendothelial and subendothelial migrations under coculture with smooth muscle cells in disturbed flow. Blood 2006, 107, 1933–1942. [Google Scholar] [CrossRef]
- Ragaseema, V.M.; Columbus, S.; Ramesh, R.; Krishnan, L. Potential of Tissue Engineered Blood Vessel as Model to Study Effect of Flow and Wall Thickness on Cellular Communication. Curr. Tissue Eng. 2013, 3, 39–46. [Google Scholar] [CrossRef]
- Robert, J.; Weber, B.; Frese, L.; Emmert, M.Y.; Schmidt, D.; von Eckardstein, A.; Rohrer, L.; Hoerstrup, S.P. A Three-Dimensional Engineered Artery Model for In Vitro Atherosclerosis Research. PLoS ONE 2013, 8, e79821. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Fu, F.; Cheng, Y.; Wang, C.; Zhao, Y.; Gu, Z. Organ-on-a-Chip Systems: Microengineering to Biomimic Living Systems. Small 2016, 12, 2253–2282. [Google Scholar] [CrossRef]
- Cochrane, A.; Albers, H.J.; Passier, R.; Mummery, C.L.; van den Berg, A.; Orlova, V.V.; van der Meer, A.D. Advanced in vitro models of vascular biology: Human induced pluripotent stem cells and organ-on-chip technology. Adv. Drug Deliv. Rev. 2019, 140, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Poussin, C.; Kramer, B.; Lanz, H.L.; Heuvel, A.V.D.; Laurent, A.; Olivier, T.; Vermeer, M.; Peric, D.; Baumer, K.; Dulize, R.; et al. 3D human microvessel-on-a-chip model for studying monocyte-to-endothelium adhesion under flow—Application in systems toxicology. ALTEX-Altern. Anim. Exp. 2020, 37, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Heo, K.-S.; Fujiwara, K.; Abe, J.-I. Shear stress and atherosclerosis. Mol. Cells 2014, 37, 435–440. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Cell Origin | Cell Markers | Culture Medium |
|---|---|---|---|
| SK-BR3 | human breast adenocarcinoma | HER2-Neu + Oestrogen − Progesterone − | DMEM + 10% FBS + 2% L-Glutamin + 1% Penicillin/Streptomycin + 1% non-essential amino acids |
| MCF-7 | human breast ductal carcinoma | HER2/Neu − Oestrogen + Progesterone + | See SK-BR3 |
| MDA-MB-231 | human breast adenocarcinoma | HER2-Neu − Oestrogen − Progesterone − | See SK-BR3 |
| HUVEC | human umbilical vein endothelial cell | Endothelial Cell Growth Base Media + Endothelial Cell Growth Supplement + 1% Penicillin/Streptomycin | |
| THP-1 | human leukaemia monocytic cell | RPMI + 10% FBS + 5% L-Glutamin + 1% Penicillin/Streptomycin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scalia, A.; Doumani, L.; Kindt, N.; Journé, F.; Trelcat, A.; Carlier, S. The Interplay between Atherosclerosis and Cancer: Breast Cancer Cells Increase the Expression of Endothelial Cell Adhesion Markers. Biology 2023, 12, 896. https://doi.org/10.3390/biology12070896
Scalia A, Doumani L, Kindt N, Journé F, Trelcat A, Carlier S. The Interplay between Atherosclerosis and Cancer: Breast Cancer Cells Increase the Expression of Endothelial Cell Adhesion Markers. Biology. 2023; 12(7):896. https://doi.org/10.3390/biology12070896
Chicago/Turabian StyleScalia, Alessandro, Lesly Doumani, Nadège Kindt, Fabrice Journé, Anne Trelcat, and Stéphane Carlier. 2023. "The Interplay between Atherosclerosis and Cancer: Breast Cancer Cells Increase the Expression of Endothelial Cell Adhesion Markers" Biology 12, no. 7: 896. https://doi.org/10.3390/biology12070896
APA StyleScalia, A., Doumani, L., Kindt, N., Journé, F., Trelcat, A., & Carlier, S. (2023). The Interplay between Atherosclerosis and Cancer: Breast Cancer Cells Increase the Expression of Endothelial Cell Adhesion Markers. Biology, 12(7), 896. https://doi.org/10.3390/biology12070896