
New Perspectives on the Role of Nuclear Proteases in Cell Death Pathways

 , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Nuclear Proteases
3. Nuclear Compartment in Cell Death
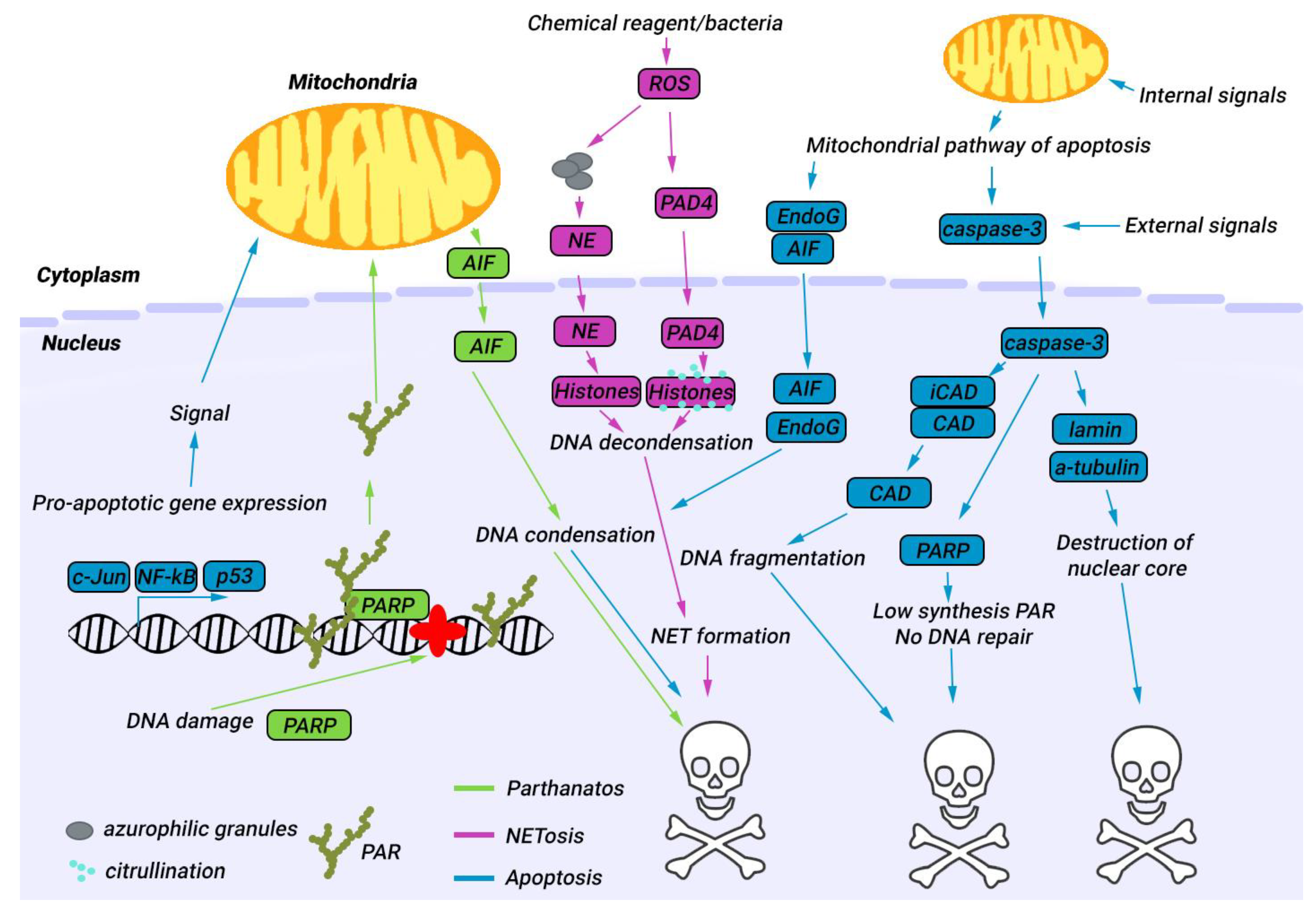
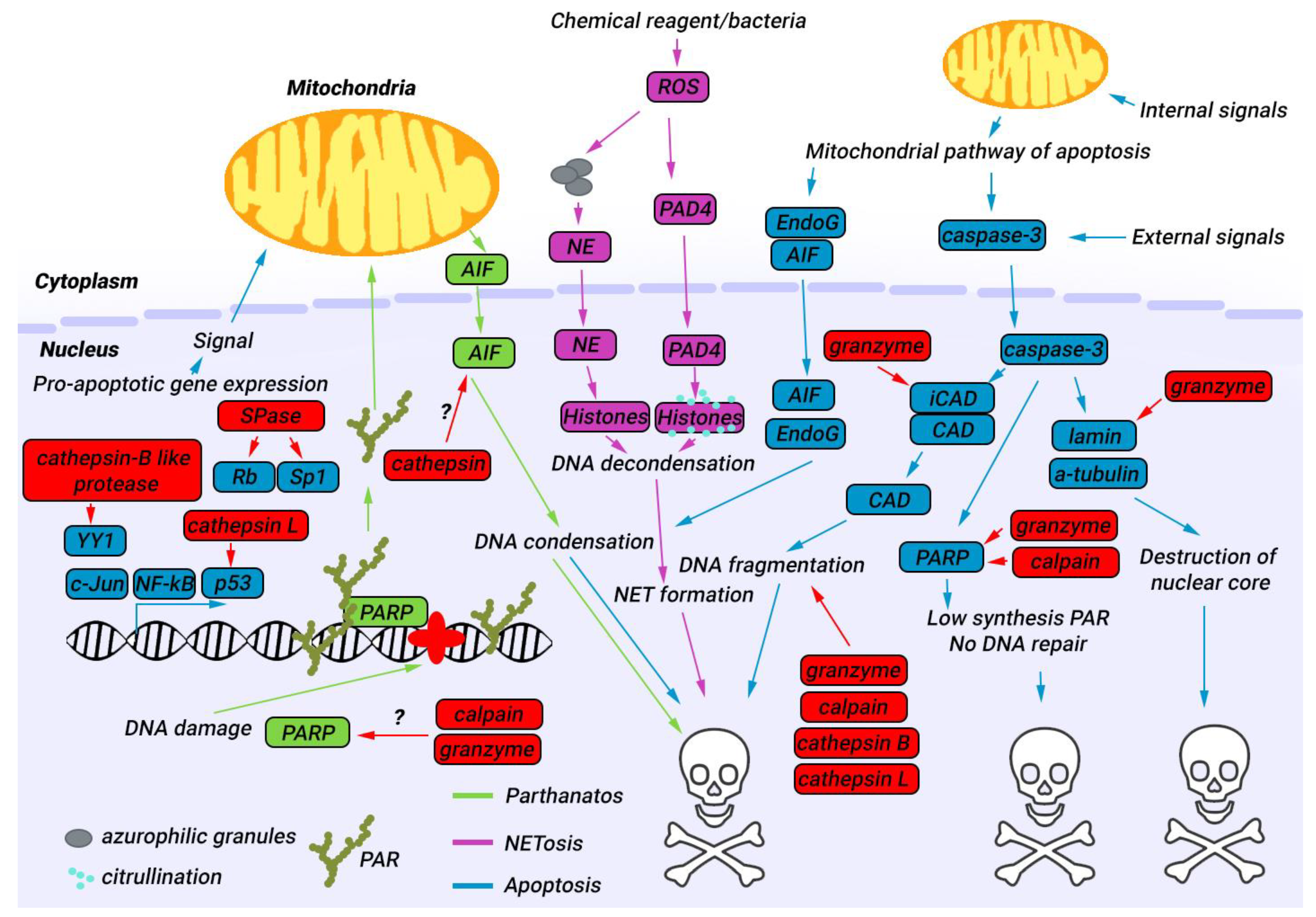
4. Nuclear Proteases in Apoptosis
5. Nuclear Proteases in Parthanatos
6. Nuclear Proteases in NETosis
7. Approaches for Modulating the Activity of Proteases in the Nucleus
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gudipaty, S.A.; Conner, C.M.; Rosenblatt, J.; Montell, D.J. Unconventional Ways to Live and Die: Cell Death and Survival in Development, Homeostasis, and Disease. Annu. Rev. Cell Dev. Biol. 2018, 34, 311–332. [Google Scholar] [CrossRef]
- Fernald, K.; Kurokawa, M. Evading apoptosis in cancer. Trends Cell Biol. 2013, 23, 620–633. [Google Scholar] [CrossRef]
- Cui, J.; Zhao, S.; Li, Y.; Zhang, D.; Wang, B.; Xie, J.; Wang, J. Regulated cell death: Discovery, features and implications for neurodegenerative diseases. Cell Commun. Signal. 2021, 19, 120. [Google Scholar] [CrossRef]
- Prokhorova, E.A.; Zamaraev, A.V.; Kopeina, G.S.; Zhivotovsky, B.; Lavrik, I.N. Role of the nucleus in apoptosis: Signaling and execution. Cell. Mol. Life Sci. 2015, 72, 4593–4612. [Google Scholar] [CrossRef]
- Saunders, C.A.; Parent, C.A. ScienceDirect Emerging roles for the nucleus during neutrophil signal relay and NETosis. Curr. Opin. Cell Biol. 2019, 62, 135–143. [Google Scholar] [CrossRef]
- Gurumallesh, P.; Alagu, K.; Ramakrishnan, B.; Muthusamy, S. A systematic reconsideration on proteases. Int. J. Biol. Macromol. 2019, 128, 254–267. [Google Scholar] [CrossRef] [PubMed]
- King, R.W.; Deshaies, R.J.; Peters, J.-M.; Kirschner, M.W. How Proteolysis Drives the Cell Cycle. Science 1996, 274, 1652–1659. [Google Scholar] [CrossRef]
- Burby, P.E.; Simmons, Z.W.; Schroeder, J.W.; Simmons, L.A. Discovery of a dual protease mechanism that promotes DNA damage checkpoint recovery. PLoS Genet. 2018, 14, e1007512. [Google Scholar] [CrossRef] [PubMed]
- Ghajar, C.M.; George, S.C.; Putnam, A.J. Matrix Metalloproteinase Control of Capillary Morphogenesis. Crit. Rev. Eukaryot. Gene Expr. 2008, 18, 251–278. [Google Scholar] [CrossRef]
- Lakka, S.S.; Gondi, C.S.; Rao, J.S. Proteases and glioma angiogenesis. Brain Pathol. 2005, 15, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, N.; Vogel, S.N.; Van Way, C.; Papasian, C.J.; Qureshi, A.A.; Morrison, D.C. The Proteasome: A Central Regulator of Inflammation and Macrophage Function. Immunol. Res. 2005, 31, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Kanneganti, T.D. Caspase-7: A protease involved in apoptosis and inflammation. Int. J. Biochem. Cell Biol. 2010, 42, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J. Caspases: Executioners of Apoptosis. In Pathobiology of Human Disease; Elsevier: Amsterdam, The Netherlands, 2014; Volume 16, pp. 145–152. ISBN 9780123864567. [Google Scholar]
- Tan, G.-J.; Peng, Z.-K.; Lu, J.-P.; Tang, F.-Q. Cathepsins mediate tumor metastasis. World J. Biol. Chem. 2013, 4, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Tedelind, S.; Jordans, S.; Resemann, H.; Blum, G.; Bogyo, M.; Führer, D.; Brix, K. Cathepsin B trafficking in thyroid carcinoma cells. Thyroid Res. 2011, 4, S2. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.M.; Sloane, B.F. Cysteine cathepsins: Multifunctional enzymes in cancer. Nat. Rev. Cancer 2006, 6, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Sloane, B.F.; Rozhin, J.; Johnson, K.; Taylor, H.; Crissman, J.D.; Honn, K.V. Cathepsin B: Association with plasma membrane in metastatic tumors. Proc. Natl. Acad. Sci. USA 1986, 83, 2483–2487. [Google Scholar] [CrossRef]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef]
- Mannello, F.; Medda, V. Nuclear localization of Matrix metalloproteinases. Prog. Histochem. Cytochem. 2012, 47, 27–58. [Google Scholar] [CrossRef]
- Soond, S.M.; Kozhevnikova, M.V.; Frolova, A.S.; Savvateeva, L.V.; Plotnikov, E.Y.; Townsend, P.A.; Han, Y.P.; Zamyatnin, A.A. Lost or Forgotten: The nuclear cathepsin protein isoforms in cancer. Cancer Lett. 2019, 462, 43–50. [Google Scholar] [CrossRef]
- Frolova, A.S.; Petushkova, A.I.; Makarov, V.A.; Soond, S.M.; Zamyatnin, A.A. Unravelling the network of nuclear matrix metalloproteinases for targeted drug design. Biology 2020, 9, 480. [Google Scholar] [CrossRef]
- Sounni, N.E.; Roghi, C.; Chabottaux, V.; Janssen, M.; Munaut, C.; Maquoi, E.; Galvez, B.G.; Gilles, C.; Frankenne, F.; Murphy, G.; et al. Up-regulation of Vascular Endothelial Growth Factor-A by Active Membrane-type 1 Matrix Metalloproteinase Through Activation of Src-Tyrosine Kinases. J. Biol. Chem. 2004, 279, 13564–13574. [Google Scholar] [CrossRef]
- Burton, L.J.; Dougan, J.; Jones, J.; Smith, B.N.; Randle, D.; Henderson, V.; Odero-Marah, V.A. Targeting the Nuclear Cathepsin L CCAAT Displacement Protein/Cut Homeobox Transcription Factor-Epithelial Mesenchymal Transition Pathway in Prostate and Breast Cancer Cells with the Z-FY-CHO Inhibitor. Mol. Cell. Biol. 2017, 37, e00297-16. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Lu, W.; Liu, S.; Yang, Q.; Shawn Goodwin, J.; Sathyanarayana, S.A.; Pratap, S.; Chen, Z. MMP7 interacts with ARF in nucleus to potentiate tumor microenvironments for prostate cancer progression in vivo. Oncotarget 2016, 7, 47609–47619. [Google Scholar] [CrossRef] [PubMed]
- Si-Tayeb, K.; Monvoisin, A.; Mazzocco, C.; Lepreux, S.; Decossas, M.; Cubel, G.; Taras, D.; Blanc, J.F.; Robinson, D.R.; Rosenbaum, J. Matrix metalloproteinase 3 is present in the cell nucleus and is involved in apoptosis. Am. J. Pathol. 2006, 169, 1390–1401. [Google Scholar] [CrossRef]
- Knapinska, A.M.; Fields, G.B. The expanding role of mt1-mmp in cancer progression. Pharmaceuticals 2019, 12, 77. [Google Scholar] [CrossRef]
- Zheng, T.S.; Schlosser, S.F.; Dao, T.; Hingorani, R.; Crispe, I.N.; Boyer, J.L.; Flavell, R.A. Caspase-3 controls both cytoplasmic and nuclear events associated with Fas-mediated apoptosis in vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 13618–13623. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Dennemärker, J.; Reinheckel, T. Specific functions of lysosomal proteases in endocytic and autophagic pathways. Biochim. Biophys. Acta—Proteins Proteom. 2012, 1824, 34–43. [Google Scholar] [CrossRef]
- Malara, A.; Ligi, D.; Di Buduo, C.; Mannello, F.; Balduini, A. Sub-Cellular Localization of Metalloproteinases in Megakaryocytes. Cells 2018, 7, 80. [Google Scholar] [CrossRef]
- Tedelind, S.; Poliakova, K.; Valeta, A.; Hunegnaw, R.; Yemanaberhan, E.L.; Heldin, N.E.; Kurebayashi, J.; Weber, E.; Kopitar-Jerala, N.; Turk, B.; et al. Nuclear cysteine cathepsin variants in thyroid carcinoma cells. Biol. Chem. 2010, 391, 923–935. [Google Scholar] [CrossRef]
- Puchi, M.; García-Huidobro, J.; Cordova, C.; Aguilar, R.; Dufey, E.; Imschenetzky, M.; Bustos, P.; Morin, V. A new nuclear protease with cathepsin L properties is present in HeLa and Caco-2 cells. J. Cell. Biochem. 2010, 111, 1099–1106. [Google Scholar] [CrossRef]
- Tamhane, T.; Wolters, B.K.; Illukkumbura, R.; Maelandsmo, G.M.; Haugen, M.H.; Brix, K. Construction of a plasmid coding for green fluores protein tagged cathepsin L and data on expression in colorectal carcinoma cells. Data Br. 2015, 5, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Bestvater, F.; Dallner, C.; Spiess, E. The C-terminal subunit of artificially truncated human cathepsin B mediates its nuclear targeting and contributes to cell viability. BMC Cell Biol. 2005, 6, 16. [Google Scholar] [CrossRef]
- Hiwasa, T.; Sakiyama, S. Nuclear localization of procathepsin L/MEP in ras-transformed mouse fibroblasts. Cancer Lett. 1996, 99, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.; Tosetto, M.; Kevans, D.; Coss, A.; Wang, L.; O’Donoghue, D.; Hyland, J.; Sheahan, K.; Mulcahy, H.; O’Sullivan, J. Localization of nuclear cathepsin L and its association with disease progression and poor outcome in colorectal cancer. Int. J. Cancer 2009, 125, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Goulet, B.; Truscott, M.; Nepveu, A. A novel proteolytically processed CDP/Cux isoform of 90 kDa is generated by cathepsin L. Biol. Chem. 2006, 387, 1285–1293. [Google Scholar] [CrossRef]
- Hill, J.W.; Poddar, R.; Thompson, J.F.; Rosenberg, G.A.; Yang, Y. Intranuclear matrix metalloproteinases promote DNA damage and apoptosis induced by oxygen-glucose deprivation in neurons. Neuroscience 2012, 220, 277–290. [Google Scholar] [CrossRef]
- Duarte, L.F.; Young, A.R.J.; Wang, Z.; Wu, H.A.; Panda, T.; Kou, Y.; Kapoor, A.; Hasson, D.; Mills, N.R.; Ma’ayan, A.; et al. Histone H3.3 and its proteolytically processed form drive a cellular senescence programme. Nat. Commun. 2014, 5, 5210. [Google Scholar] [CrossRef]
- Ip, Y.C.; Cheung, S.T.; Fan, S.T. Atypical localization of membrane type 1-matrix metalloproteinase in the nucleus is associated with aggressive features of hepatocellular carcinoma. Mol. Carcinog. 2007, 46, 225–230. [Google Scholar] [CrossRef]
- Mäkinen, L.K.; Häyry, V.; Atula, T.; Haglund, C.; Keski-Säntti, H.; Leivo, I.; Mäkitie, A.; Passador-Santos, F.; Böckelman, C.; Salo, T.; et al. Prognostic significance of matrix metalloproteinase-2, -8, -9, and -13 in oral tongue cancer. J. Oral Pathol. Med. 2012, 41, 394–399. [Google Scholar] [CrossRef]
- Stratigopoulos, G.; LeDuc, C.A.; Cremona, M.L.; Chung, W.K.; Leibel, R.L. Cut-like homeobox 1 (CUX1) regulates expression of the fat mass and obesity-associated and retinitis pigmentosa GTPase regulator-interacting protein-1-like (RPGRIP1L) genes and coordinates leptin receptor signaling. J. Biol. Chem. 2011, 286, 2155–2170. [Google Scholar] [CrossRef]
- Havel, L.S.; Li, S.; Li, X.-J. Nuclear accumulation of polyglutamine disease proteins and neuropathology. Mol. Brain 2009, 2, 21. [Google Scholar] [CrossRef]
- Békés, M.; Langley, D.R.; Crews, C.M. PROTAC targeted protein degraders: The past is prologue. Nat. Rev. Drug Discov. 2022, 21, 181–200. [Google Scholar] [CrossRef]
- Kwan, J.A.; Schulze, C.J.; Wang, W.; Leon, H.; Sariahmetoglu, M.; Sung, M.; Sawicka, J.; Sims, D.E.; Sawicki, G.; Schulz, R. Matrix metalloproteinase-2 (MMP-2) is present in the nucleus of cardiac myocytes and is capable of cleaving poly (ADP-ribose) polymerase (PARP) in vitro. FASEB J. 2004, 18, 690–692. [Google Scholar] [CrossRef] [PubMed]
- Lammerding, J. Mechanics of the nucleus. Compr. Physiol. 2011, 1, 783–807. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Llambi, F. Cell Death Signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef]
- Schweichel, J.-U.; Merker, H.-J. The morphology of various types of cell death in prenatal tissues. Teratology 1973, 7, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Kroemer, G.; El-Deiry, W.S.; Golstein, P.; Peter, M.E.; Vaux, D.; Vandenabeele, P.; Zhivotovsky, B.; Blagosklonny, M.V.; Malorni, W.; Knight, R.A.; et al. Classification of cell death: Recommendations of the nomenclature committee on cell death. Cell Death Differ. 2005, 12, 1463–1467. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Nicholson, D.W.; Thornberry, N.A. Caspases: Killer proteases. Trends Biochem. Sci. 1997, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zou, H.; Slaughter, C.; Wang, X. DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell 1997, 89, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Sakahira, H.; Enari, M.; Nagata, S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 1998, 391, 96–99. [Google Scholar] [CrossRef]
- Enari, M.; Sakahira, H.; Yokoyama, H.; Okawa, K.; Iwamatsu, A.; Nagata, S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 1998, 391, 43–50. [Google Scholar] [CrossRef]
- Andrabi, S.A.; Dawson, T.M.; Dawson, V.L. Mitochondrial and nuclear cross talk in cell death: Parthanatos. Ann. N. Y. Acad. Sci. 2008, 1147, 233–241. [Google Scholar] [CrossRef]
- David, K.K.; Andrabi, S.A.; Dawson, T.M.; Dawson, V.L. Parthanatos, a messenger of death. Front. Biosci. 2009, 14, 1116. [Google Scholar] [CrossRef] [PubMed]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochemistry (Mosc.) 2020, 85, 1178. [Google Scholar] [CrossRef]
- Thiam, H.R.; Wong, S.L.; Wagner, D.D.; Waterman, C.M. Cellular Mechanisms of NETosis. Annu. Rev. Cell Dev. Biol. 2021, 36, 191–218. [Google Scholar] [CrossRef]
- Neubert, E.; Meyer, D.; Rocca, F.; Günay, G.; Kwaczala-tessmann, A.; Grandke, J.; Senger-sander, S.; Geisler, C.; Egner, A.; Schön, M.P.; et al. Chromatin swelling drives neutrophil extracellular trap release. Nat. Commun. 2018, 9, 3767. [Google Scholar] [CrossRef]
- Sheshachalam, A.; Srivastava, N.; Mitchell, T.; Lacy, P.; Eitzen, G. Granule protein processing and regulated secretion in neutrophils. Front. Immunol. 2014, 5, 448. [Google Scholar] [CrossRef]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Hamam, H.J.; Palaniyar, N. Post-Translational Modifications in NETosis and NETs-Mediated Diseases. Biomolecules 2019, 9, 369. [Google Scholar] [CrossRef]
- Ruchaud, S.; Korfali, N.; Villa, P.; Kottke, T.J.; Dingwall, C.; Kaufmann, S.H.; Earnshaw, W.C. Caspase-6 gene disruption reveals a requirement for lamin A cleavage in apoptotic chromatin condensation. EMBO J. 2002, 21, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Kenji Takahashi Calpain Substrate Specificity. In Intracellular Calcium-Dependent Proteolysis; CRC Press: Boca Raton, FL, USA, 1990; p. 20. ISBN 9781351073813.
- Chen, K.W.; Monteleone, M.; Boucher, D.; Sollberger, G.; Ramnath, D.; Condon, N.D.; von Pein, J.B.; Broz, P.; Sweet, M.J.; Schroder, K. Noncanonical inflammasome signaling elicits gasdermin D–dependent neutrophil extracellular traps. Sci. Immunol. 2018, 3, eaar6676. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Weigel, B.; Mall, M.; Werth, V.P.; Liu, M. Nuclear envelope rupture and NET formation is driven by PKCα-mediated lamin B disassembly. EMBO Rep. 2020, 21, e48779. [Google Scholar] [CrossRef] [PubMed]
- Torabi, B.; Flashner, S.; Beishline, K.; Sowash, A.; Donovan, K.; Bassett, G.; Azizkhan-Clifford, J. Caspase cleavage of transcription factor Sp1 enhances apoptosis. Apoptosis 2018, 23, 65–78. [Google Scholar] [CrossRef]
- Tewari, M.; Quan, L.T.; O’Rourke, K.; Desnoyers, S.; Zeng, Z.; Beidler, D.R.; Poirier, G.G.; Salvesen, G.S.; Dixit, V.M. Yama/CPP32β, a mammalian homolog of CED-3, is a CrmA-inhibitable protease that cleaves the death substrate poly(ADP-ribose) polymerase. Cell 1995, 81, 801–809. [Google Scholar] [CrossRef]
- Kim, B.J.; Lee, H. Caspase-mediated cleavage of importin-α increases its affinity for MCM and downregulates DNA synthesis by interrupting the binding of MCM to chromatin. Biochim. Biophys. Acta-Mol. Cell Res. 2008, 1783, 2287–2293. [Google Scholar] [CrossRef] [PubMed]
- Ubeda, M.; Habener, J.F. The Large Subunit of the DNA Replication Complex C (DSEB/RF-C140) Cleaved and Inactivated by Caspase-3 (CPP32/YAMA) during Fas-induced Apoptosis. J. Biol. Chem. 1997, 272, 19562–19568. [Google Scholar] [CrossRef]
- Huang, Y.; Nakada, S.; Ishiko, T.; Utsugisawa, T.; Datta, R.; Kharbanda, S.; Yoshida, K.; Talanian, R.V.; Weichselbaum, R.; Kufe, D.; et al. Role for Caspase-Mediated Cleavage of Rad51 in Induction of Apoptosis by DNA Damage. Mol. Cell. Biol. 1999, 19, 2986–2997. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kidd, V.J. Cleavage of DFF-45/ICAD by Multiple Caspases Is Essential for Its Function during Apoptosis. J. Biol. Chem. 1998, 273, 28549–28552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Larner, S.F.; Liu, M.C.; Zheng, W.; Hayes, R.L.; Wang, K.K.W. Multiple alphaII-spectrin breakdown products distinguish calpain and caspase dominated necrotic and apoptotic cell death pathways. Apoptosis 2009, 14, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Kenig, S.; Frangež, R.; Pucer, A.; Lah, T. Inhibition of cathepsin L lowers the apoptotic threshold of glioblastoma cells by up-regulating p53 and transcription of caspases 3 and 7. Apoptosis 2011, 16, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Vancompernolle, K.; Van Herreweghe, F.; Pynaert, G.; Van de Craen, M.; De Vos, K.; Totty, N.; Sterling, A.; Fiers, W.; Vandenabeele, P.; Grooten, J. Atractyloside-induced release of cathepsin B, a protease with caspase-processing activity. FEBS Lett. 1998, 438, 150–158. [Google Scholar] [CrossRef]
- Zhang, D.; Beresford, P.J.; Greenberg, A.H.; Lieberman, J. Granzymes A and B directly cleave lamins and disrupt the nuclear lamina during granule-mediated cytolysis. Proc. Natl. Acad. Sci. USA 2001, 98, 5746–5751. [Google Scholar] [CrossRef]
- Lu, H.; Hou, Q.; Zhao, T.; Zhang, H.; Zhang, Q.; Wu, L.; Fan, Z. Granzyme M Directly Cleaves Inhibitor of Caspase-Activated DNase (CAD) to Unleash CAD Leading to DNA Fragmentation. J. Immunol. 2006, 177, 1171–1178. [Google Scholar] [CrossRef]
- Froelich, C.J.; Hanna, W.L.; Poirier, G.G.; Duriez, P.J.; D’amours, D.; Salvesen, G.S.; Alnemri, E.S.; Earnshaw, W.C.; Shah, G.M. Granzyme B/Perforin-Mediated Apoptosis of Jurkat Cells Results in Cleavage of Poly(ADP-ribose) Polymerase to the 89-kDa Apoptotic Fragment and Less Abundant 64-kDa Fragment. Biochem. Biophys. Res. Commun. 1996, 227, 658–665. [Google Scholar] [CrossRef]
- Pinkoski, M.J.; Heibein, J.A.; Barry, M.; Bleackley, R.C. Nuclear translocation of granzyme B in target cell apoptosis. Cell Death Differ. 2000, 7, 17–24. [Google Scholar] [CrossRef]
- McGinnis, K.M.; Gnegy, M.E.; Park, Y.H.; Mukerjee, N.; Wang, K.K.W. Procaspase-3 and Poly(ADP)ribose polymerase (PARP) are calpain substrates. Biochem. Biophys. Res. Commun. 1999, 263, 94–99. [Google Scholar] [CrossRef]
- Tremper-Wells, B.; Vallano, M. Lou Nuclear calpain regulates Ca2+-dependent signaling via proteolysis of nuclear Ca2+/calmodulin-dependent protein kinase type IV in cultured neurons. J. Biol. Chem. 2005, 280, 2165–2175. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Takeichi, M. NMDA-Receptor Activation Induces Calpain-Mediated β-Catenin Cleavages for Triggering Gene Expression. Neuron 2007, 53, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Hirai, S.; Kawasaki, H.; Yaniv, M.; Suzuki, K. Degradation of transcription factors, c-Jun and c-Fos, by calpain. FEBS Lett. 1991, 287, 57–61. [Google Scholar] [CrossRef]
- Pariat, M.; Salvat, C.; Bebien, M.; Brockly, F.; Altieri, E.; Carillo, S.; Jariel-Encontre, I.; Piechaczyk, M. The sensitivity of c-Jun and c-Fos proteins to calpains depends on conformational determinants of the monomers and not on formation of dimers. Society 2000, 138, 129–138. [Google Scholar] [CrossRef]
- Mao, X.; Yang, S.-H.; Simpkins, J.W.; Barger, S.W. Glutamate receptor activation evokes calpain-mediated degradation of Sp3 and Sp4, the prominent Sp-family transcription factors in neurons. J. Neurochem. 2007, 100, 1300–1314. [Google Scholar] [CrossRef] [PubMed]
- Kubbutat, M.H.; Vousden, K.H. Proteolytic cleavage of human p53 by calpain: A potential regulator of protein stability. Mol. Cell. Biol. 1997, 17, 460–468. [Google Scholar] [CrossRef]
- Nishinaka, T.; Fu, Y.H.F.; Chen, L.I.; Yokoyama, K.; Chiu, R. A unique cathepsin-like protease isolated from CV-1 cells is involved in rapid degradation of retinoblastoma susceptibility gene product, RB, and transcription factor SP1. Biochim. Biophys. Acta-Gene Struct. Expr. 1997, 1351, 274–286. [Google Scholar] [CrossRef]
- Gößwein, S.; Lindemann, A.; Mahajan, A.; Maueröder, C.; Martini, E.; Patankar, J.; Schett, G.; Becker, C.; Wirtz, S.; Naumann-Bartsch, N.; et al. Citrullination licenses calpain to decondense nuclei in neutrophil extracellular trap formation. Front. Immunol. 2019, 10, 2481. [Google Scholar] [CrossRef]
- Dhaenens, M.; Glibert, P.; Lambrecht, S.; Vossaert, L.; Van Steendam, K.; Elewaut, D.; Deforce, D. Neutrophil Elastase in the capacity of the “H2A-specific protease”. Int. J. Biochem. Cell Biol. 2014, 51, 39–44. [Google Scholar] [CrossRef]
- Yuste, V.J.; Moubarak, R.S.; Delettre, C.; Bras, M.; Sancho, P.; Robert, N.; D’Alayer, J.; Susin, S.A. Cysteine protease inhibition prevents mitochondrial apoptosis-inducing factor (AIF) release. Cell Death Differ. 2005, 12, 1445–1448. [Google Scholar] [CrossRef]
- Solary, E.; Eymin, B.; Droin, N.; Haugg, M. Proteases, proteolysis, and apoptosis. Cell Biol. Toxicol. 1998, 14, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, D.W. Caspase structure, proteolytic substrates, and function during apoptotic cell death. Cell Death Differ. 1999, 6, 1028–1042. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, G.; Benedict, M.A.; Hu, Y.; Inohara, N. Caspases: The proteases of the apoptotic pathway. Oncogene 1998, 17, 3237–3245. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Ishisaka, R.; Utsumi, K.; Utsumi, T. Involvement of Lysosomal Cysteine Proteases in Hydrogen Peroxide-induced Apoptosis in HL-60 Cells. Biosci. Biotechnol. Biochem. 2002, 66, 1865–1872. [Google Scholar] [CrossRef]
- Chow, S.C.; Weis, M.; Kass, G.E.; Holmstro, T.H.; Eriksson, J.E.; Orrenius, S. Involvement of multiple proteases during Fas-mediated apoptosis in T lymphocytes. FEBS Lett. 1995, 364, 134–138. [Google Scholar] [CrossRef]
- Schlegel, J.; Peters, I.; Orrenius, S. Isolation and partial characterization of a protease involved in Fas-induced apoptosis. FEBS Lett. 1995, 364, 139–142. [Google Scholar] [CrossRef]
- Zhivotovsky, B.; Wade, D.; Gahm, A.; Orrenius, S.; Nicotera, P. Formation of 50 kbp chromatin fragments in isolated liver nuclei is mediated by protease and endonuclease activation. FEBS Lett. 1994, 351, 150–154. [Google Scholar] [CrossRef]
- Momeni, H.R. Role of Calpain in Apoptosis. Cell J. 2011, 13, 65–72. [Google Scholar]
- Hay, Z.L.Z.; Slansky, J.E. Granzymes: The Molecular Executors of Immune-Mediated Cytotoxicity. Int. J. Mol. Sci. 2022, 23, 1833. [Google Scholar] [CrossRef]
- Chowdhury, D.; Lieberman, J. Death by a Thousand Cuts: Granzyme Pathways of Programmed Cell Death. Annu. Rev. Immunol. 2008, 26, 389–420. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jiao, Y.; Gong, X.; Liu, J.; Xiao, H.; Zheng, Q. Role of transcription factors in apoptotic cells clearance. Front. Cell Dev. Biol. 2023, 11, 1110225. [Google Scholar] [CrossRef] [PubMed]
- Pizzorno, M.C. Nuclear cathepsin B-like protease cleaves transcription factor YY1 in differentiated cells. Biochim. Biophys. Acta-Mol. Basis Dis. 2001, 1536, 31–42. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhu, Y.; Yang, C.; Liu, X.; Peng, Y.P.; Jiang, K.R.; Miao, Y.; Xu, Z.K. Yin Yang-1 increases apoptosis through Bax activation in pancreatic cancer cells. Oncotarget 2016, 7, 28498–28509. [Google Scholar] [CrossRef] [PubMed]
- Deniaud, E.; Baguet, J.; Mathieu, A.L.; Pagès, G.; Marvel, J.; Leverrier, Y. Overexpression of Sp1 transcription factor induces apoptosis. Oncogene 2006, 25, 7096–7105. [Google Scholar] [CrossRef]
- Zhu, T.; Zheng, J.Y.; Huang, L.L.; Wang, Y.H.; Yao, D.F.; Dai, H. Bin Human PARP1 substrates and regulators of its catalytic activity: An updated overview. Front. Pharmacol. 2023, 14, 1137151. [Google Scholar] [CrossRef] [PubMed]
- Fatokun, A.A.; Dawson, V.L.; Dawson, T.M. Parthanatos: Mechanisms and therapeutic opportunities. Br. J. Pharmacol. 2016, 171, 2000–2016. [Google Scholar] [CrossRef]
- Von Köckritz-Blickwede, M.; Goldmann, O.; Thulin, P.; Heinemann, K.; Norrby-Teglund, A.; Rohde, M.; Medina, E. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 2008, 111, 3070–3080. [Google Scholar] [CrossRef]
- Obata-ninomiya, K.; Karasuyama, H.; Djonov, V.; Eggel, A. NADPH Oxidase–Independent Formation of Extracellular DNA Traps by Basophils. J. Immunol. 2014, 192, 5314–5423. [Google Scholar] [CrossRef]
- Granger, V.; Faille, D.; Marani, V.; Noël, B.; Gallais, Y.; Szely, N.; Flament, H.; Pallardy, M.; Chollet-Martin, S.; de Chaisemartin, L. Human blood monocytes are able to form extracellular traps. J. Leukoc. Biol. 2017, 102, 775–781. [Google Scholar] [CrossRef]
- Chow, O.A.; von Köckritz-Blickwede, M.; Bright, A.T.; Hensler, M.E.; Zinkernagel, A.S.; Cogen, A.L.; Gallo, R.L.; Monestier, M.; Wang, Y.; Glass, C.K.; et al. Statins Enhance Formation of Phagocyte Extracellular Traps. Cell Host Microbe 2010, 8, 445–454. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Wang, Q.; Wei, X.; Wang, H.; Gu, K. The regulatory mechanism of neutrophil extracellular traps in cancer biological behavior. Cell Biosci. 2021, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A. A Myeloperoxidase-Containing Complex Regulates Neutrophil Elastase Release and Actin Dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Dhaenens, M.; Glibert, P.; Meert, P.; Vossaert, L.; Deforce, D. Histone proteolysis: A proposal for categorization into ‘clipping’ and ‘degradation’. BioEssays 2015, 37, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, G.S. NETosis occurs independently of neutrophil serine proteases. J. Biol. Chem. 2020, 295, 17624–17631. [Google Scholar] [CrossRef]
- Jackson, H.W.; Defamie, V.; Waterhouse, P.; Khokha, R. TIMPs: Versatile extracellular regulators in cancer. Nat. Rev. Cancer 2017, 17, 38–53. [Google Scholar] [CrossRef]
- Turk, V.; Stoka, V.; Turk, D. Cystatins: Biochemical and structural properties, and medical relevance. Front. Biosci. 2008, 13, 5406–5420. [Google Scholar] [CrossRef]
- Imre, G.; Berthelet, J.; Heering, J.; Kehrloesser, S.; Melzer, I.M.; Lee, B.I.; Thiede, B.; Dötsch, V.; Rajalingam, K. Apoptosis inhibitor 5 is an endogenous inhibitor of caspase-2. EMBO Rep. 2017, 18, 733–744. [Google Scholar] [CrossRef]
- Wendt, A.; Thompson, V.F.; Goll, D.E. Interaction of Calpastatin with Calpain: A Review. Biol. Chem. 2004, 385, 465–472. [Google Scholar] [CrossRef]
- Li, H.; Nishio, K.; Yamashita, K.; Hayakawa, T.; Hoshino, T. Cell cycle-dependent localization of tissue inhibitor of metalloproteinases-1 immunoreactivity in cultured human gingival fibroblasts. Nagoya J. Med. Sci. 1995, 58, 133–142. [Google Scholar] [CrossRef]
- Rudzinska-Radecka, M.; Frolova, A.S.; Balakireva, A.V.; Gorokhovets, N.V.; Pokrovsky, V.S.; Sokolova, D.V.; Korolev, D.O.; Potoldykova, N.V.; Vinarov, A.Z.; Parodi, A.; et al. In Silico, In Vitro, and Clinical Investigations of Cathepsin B and Stefin A mRNA Expression and a Correlation Analysis in Kidney Cancer. Cells 2022, 11, 1455. [Google Scholar] [CrossRef]
- Kopitar-Jerala, N. Cysteine Proteinase Inhibitors in the Nucleus and Nucleolus in Activated Macrophages. In Proteins of the Nucleolus: Regulation, Translocation, & Biomedical Functions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 305–321. [Google Scholar] [CrossRef]
- Lamort, A.S.; Hamon, Y.; Czaplewski, C.; Gieldon, A.; Seren, S.; Coquet, L.; Lecaille, F.; Lesner, A.; Lalmanach, G.; Gauthier, F.; et al. Processing and Maturation of Cathepsin C Zymogen: A Biochemical and Molecular Modeling Analysis. Int. J. Mol. Sci. 2019, 20, 4747. [Google Scholar] [CrossRef] [PubMed]
- Turk, B.; Dolenc, I.; Lenarčič, B.; Križaj, I.; Turk, V.; Bieth, J.G.; Björk, I. Acidic pH as a physiological regulator of human cathepsin L activity. Eur. J. Biochem. 1999, 259, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Young Choi, K.; Swierczewska, M.; Lee, S.; Chen, X. Protease-Activated Drug Development. Theranostics 2012, 2012, 156–178. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Dixit, R.; Pandey, K.C. Cysteine proteases: Modes of activation and future prospects as pharmacological targets. Front. Pharmacol. 2016, 7, 107. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cell Death | Protease | Substrate in Nucleus | Substrate Cell Function | What Happened after Cleavage | Ref. |
|---|---|---|---|---|---|
| apoptosis | caspase-3 | Sp1 | Transcription factor | Apoptosis | [69] |
| PARP-1 | DNA repair | Activation of apoptosis | [70] | ||
| lamin | Nuclear envelope | Degradation of nucleus | [65] | ||
| importin-α | Import of protein in cell nucleus | Downregulate DNA synthesis | [71] | ||
| large subunit of the DNA replication complex C | Regulation of DNA replication | Decrease DNA binding | [72] | ||
| Rad51 | DNA repair | Activation of apoptosis | [73] | ||
| ICAD | Inhibition of CAD | DNA fragmentation | [54,74] | ||
| calpain | lamin A | Nuclear envelope | Degradation of nucleus | [66] | |
| lamin B | Nuclear envelope | Degradation of nucleus | [66] | ||
| spectrin | Skeletal protein | Product of SBPD145, 150i, 120 Activation of apoptosis | [75] | ||
| cathepsin L | p53 | Transcription factor, regulation of caspase-7 expression | Silencing of CtsL induce the decrease in p53 | [76] | |
| prohibitin | Transcription factor, regulation of caspase-7 expression | Silencing of CtsL induce the decrease in p53 | [76] | ||
| cathepsin B | - | - | DNA condensation and fragmentation | [77] | |
| granzyme | lamin | Nuclear envelope | - | [78] | |
| PARP | DNA repair | - | [79,80] | ||
| ICAD | Inhibition of CAD | - | [79] | ||
| - | - | DNA fragmentation | [81] | ||
| ?—apoptosis | calpain | PARP | DNA repair | - | [82] |
| CaMK4 | Calcium signaling, regulates β-cell apoptosis | - | [83] | ||
| β-catenin | Transcription factor, regular expression of Wnt pathways genes | - | [84] | ||
| c-Fos | Transcription factor | - | [85,86] | ||
| c-Jun | Transcription factor | - | [85,86] | ||
| Sp3, Sp4 | Transcription factor | - | [87] | ||
| p53 | Transcription factor | - | [88] | ||
| SPase | Sp1 | Transcription factor | - | [89] | |
| Rb | Regulates cell growth | - | [89] | ||
| NETosis | calpain | H3 | Maintains structure of DNA | Degradation of nuclear envelope | [90] |
| HP1a | Gene regulation | Chromatin decondensation | [90] | ||
| lamin A/C | Nuclear core structure | Degradation of nuclear envelope | [90] | ||
| H3 | Maintains structure of DNA | Degradation of nuclear envelope | [90] | ||
| ? | ? | Chromatin decondensation | [90] | ||
| neutrophil elastase | H1, H2A, H2B, H3, H3 | Maintains structure of DNA | Chromatin decondensation | [62,91] | |
| ?—parthanatos | calpain | PARP | - | - | [82] |
| granzyme | PARP | - | - | [79] | |
| cysteine protease/cathepsin | AIF | - | - | [76,92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frolova, A.S.; Chepikova, O.E.; Deviataikina, A.S.; Solonkina, A.D.; Zamyatnin, A.A., Jr. New Perspectives on the Role of Nuclear Proteases in Cell Death Pathways. Biology 2023, 12, 797. https://doi.org/10.3390/biology12060797
Frolova AS, Chepikova OE, Deviataikina AS, Solonkina AD, Zamyatnin AA Jr. New Perspectives on the Role of Nuclear Proteases in Cell Death Pathways. Biology. 2023; 12(6):797. https://doi.org/10.3390/biology12060797
Chicago/Turabian StyleFrolova, Anastasia S., Olga E. Chepikova, Anna S. Deviataikina, Alena D. Solonkina, and Andrey A. Zamyatnin, Jr. 2023. "New Perspectives on the Role of Nuclear Proteases in Cell Death Pathways" Biology 12, no. 6: 797. https://doi.org/10.3390/biology12060797
APA StyleFrolova, A. S., Chepikova, O. E., Deviataikina, A. S., Solonkina, A. D., & Zamyatnin, A. A., Jr. (2023). New Perspectives on the Role of Nuclear Proteases in Cell Death Pathways. Biology, 12(6), 797. https://doi.org/10.3390/biology12060797