
Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review
 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Bibliometric Analysis of Selenium Bioaccumulation in Microalgae
2.2. Systematic Analysis of Genes Related to Selenium Metabolism
3. Results and Discussion
3.1. Bibliometric Analysis of Selenium Bioaccumulation in Microalgae
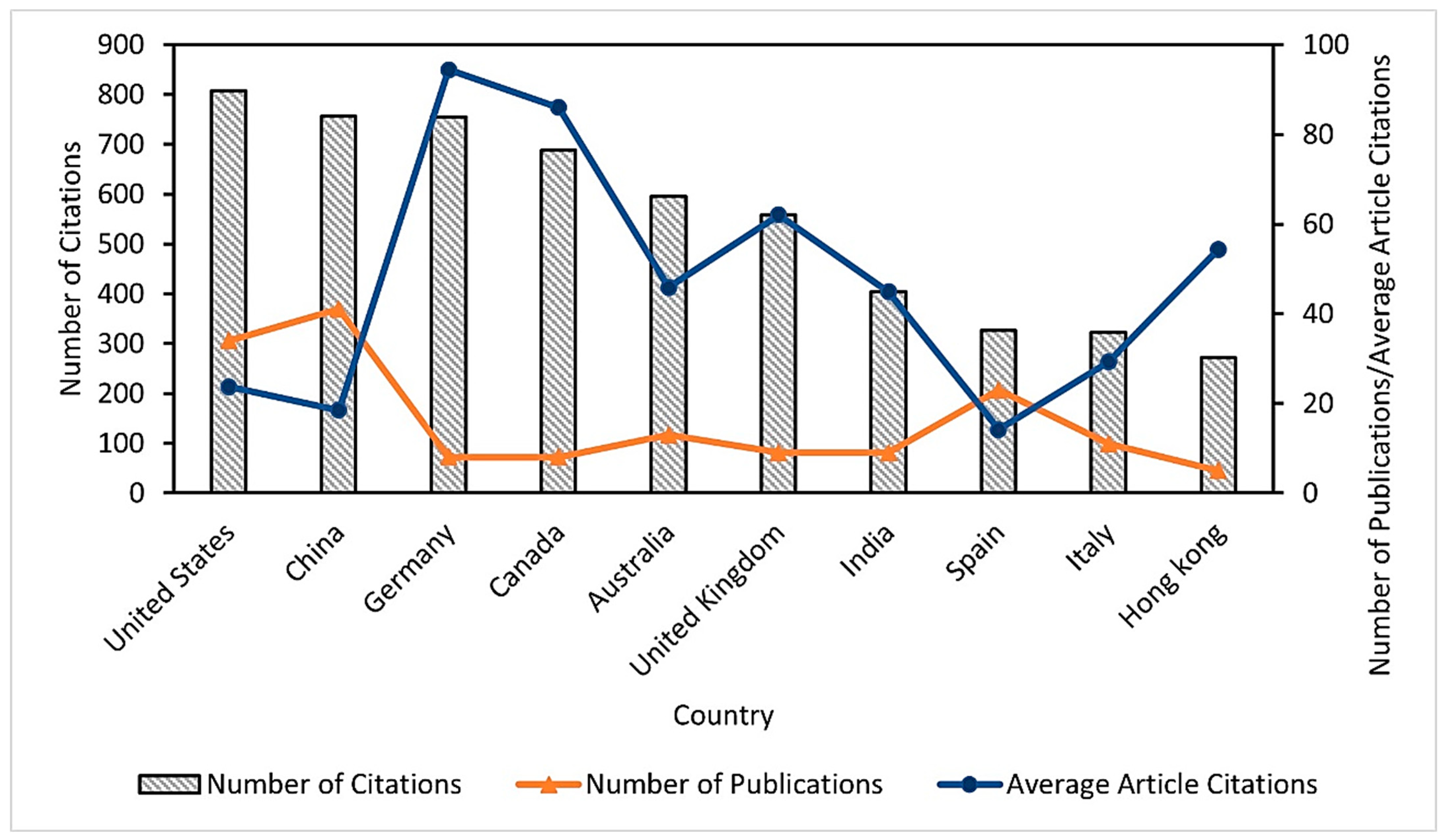
3.1.1. Scientific Production
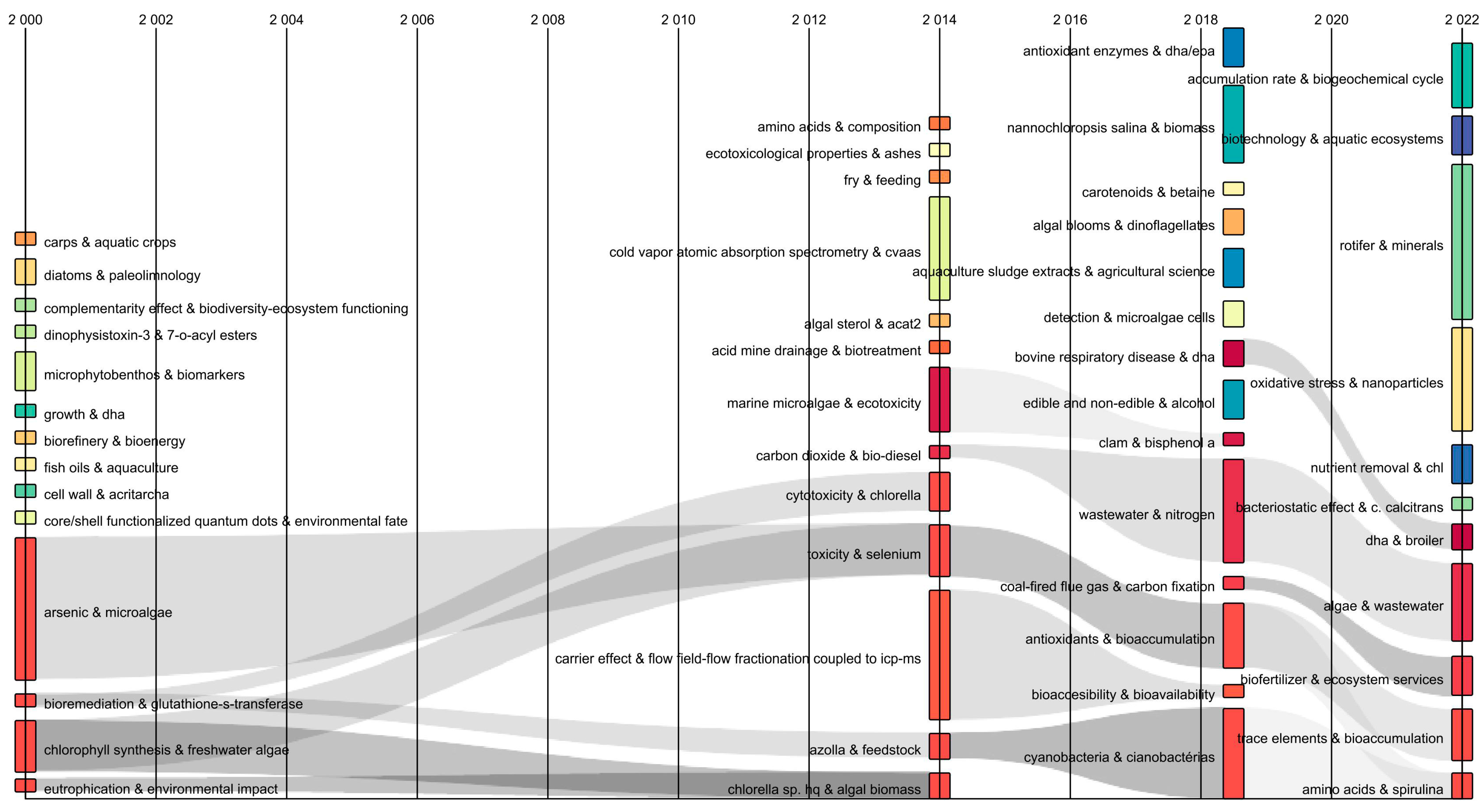
3.1.2. Sankey Diagram
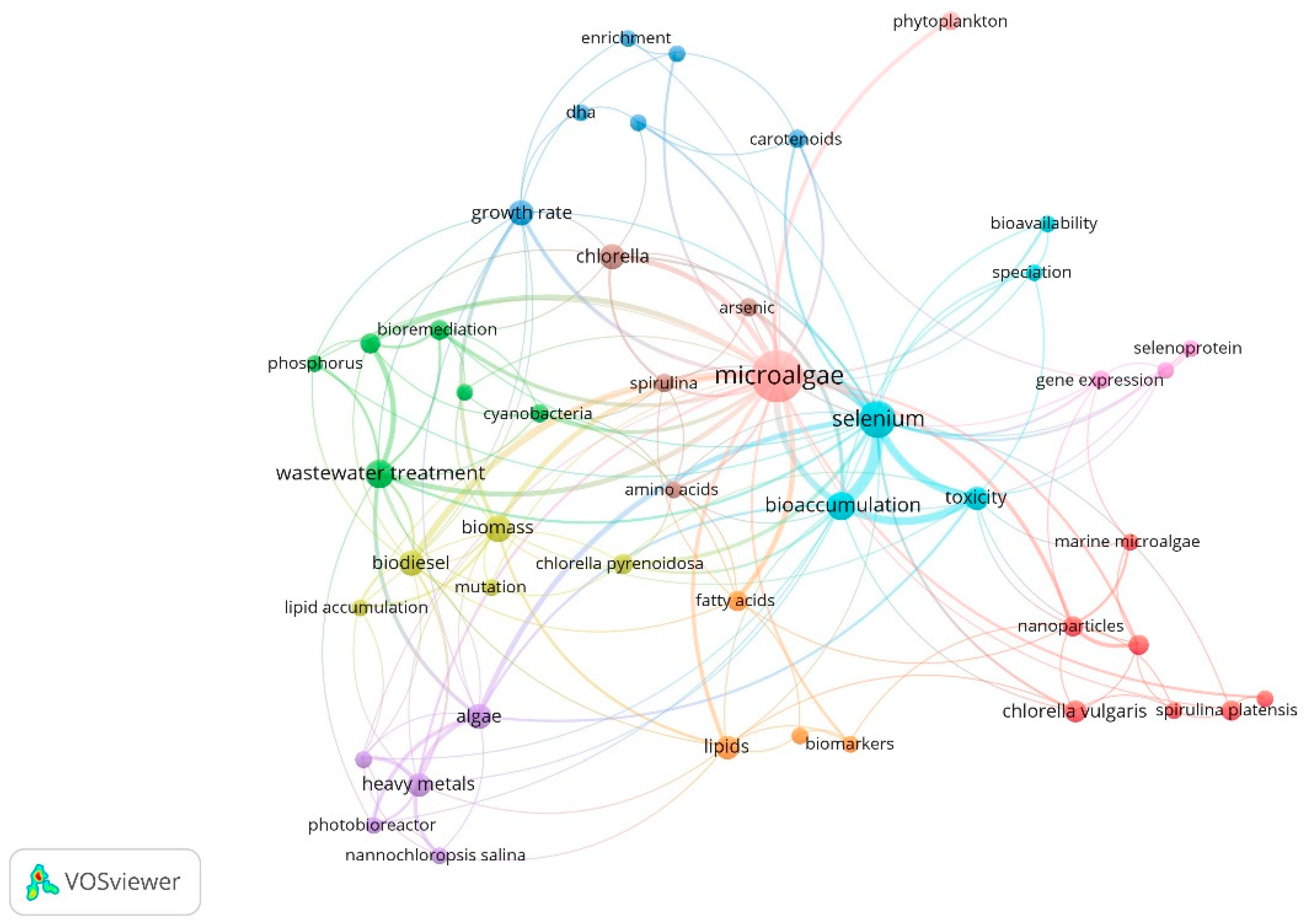
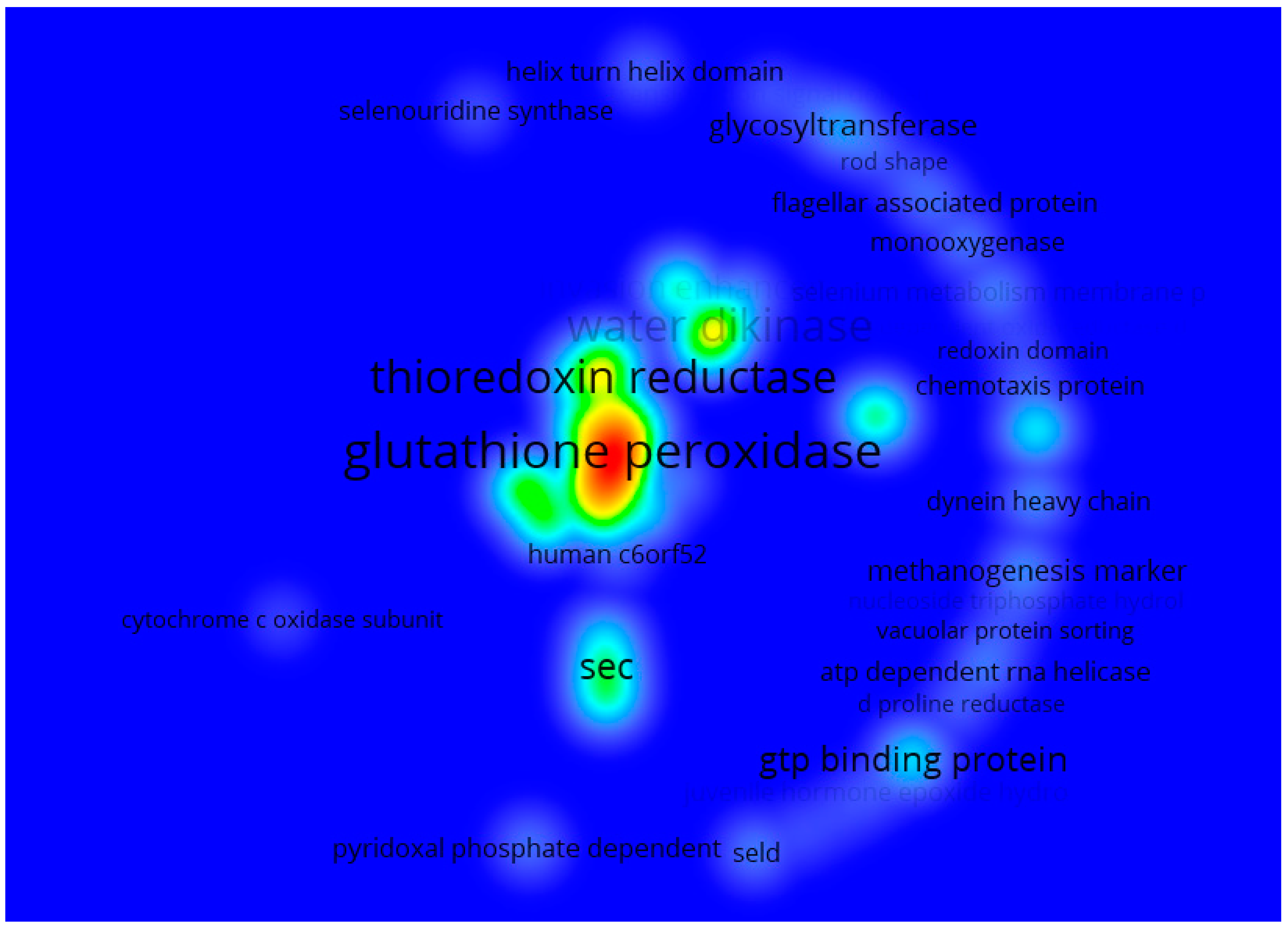
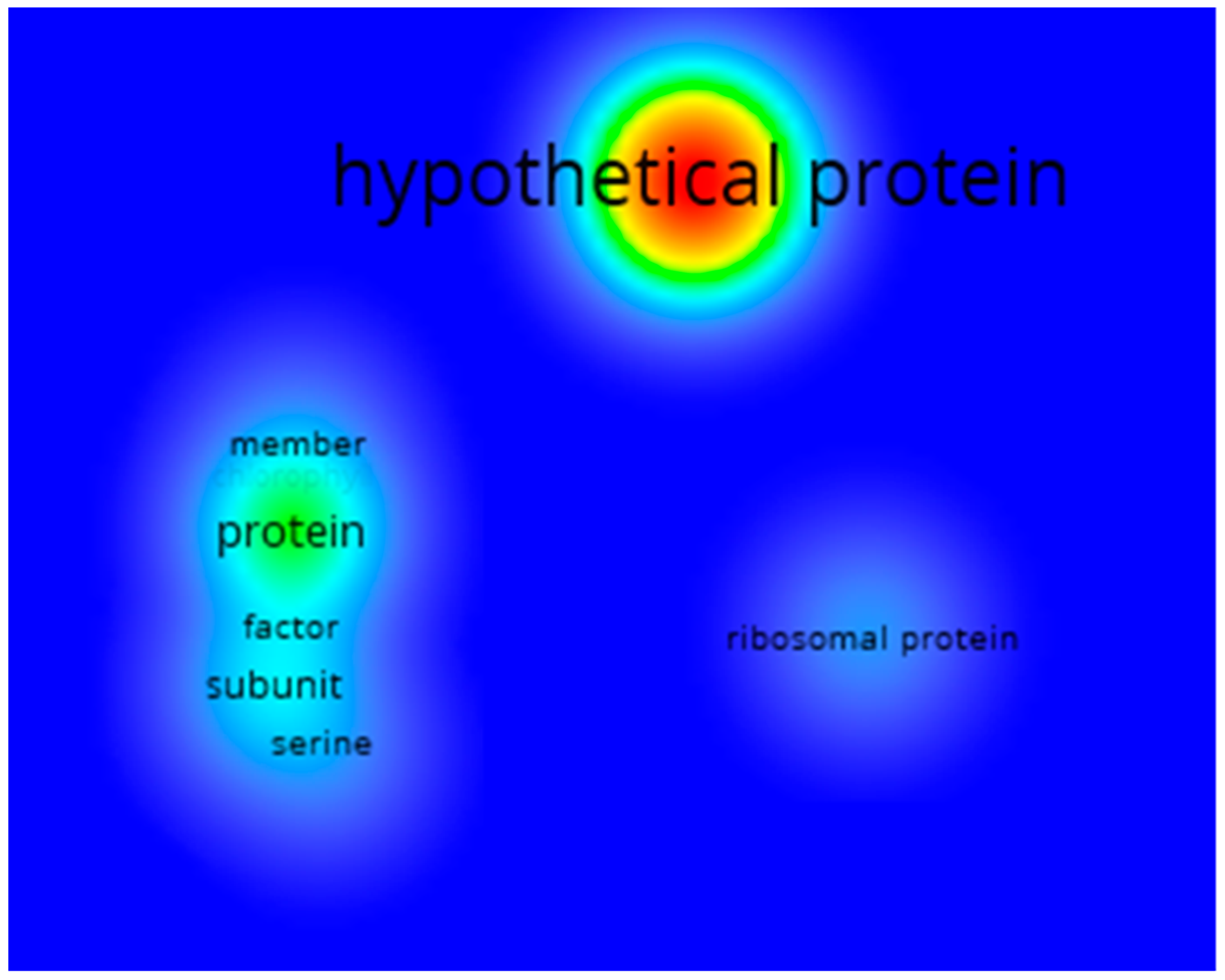
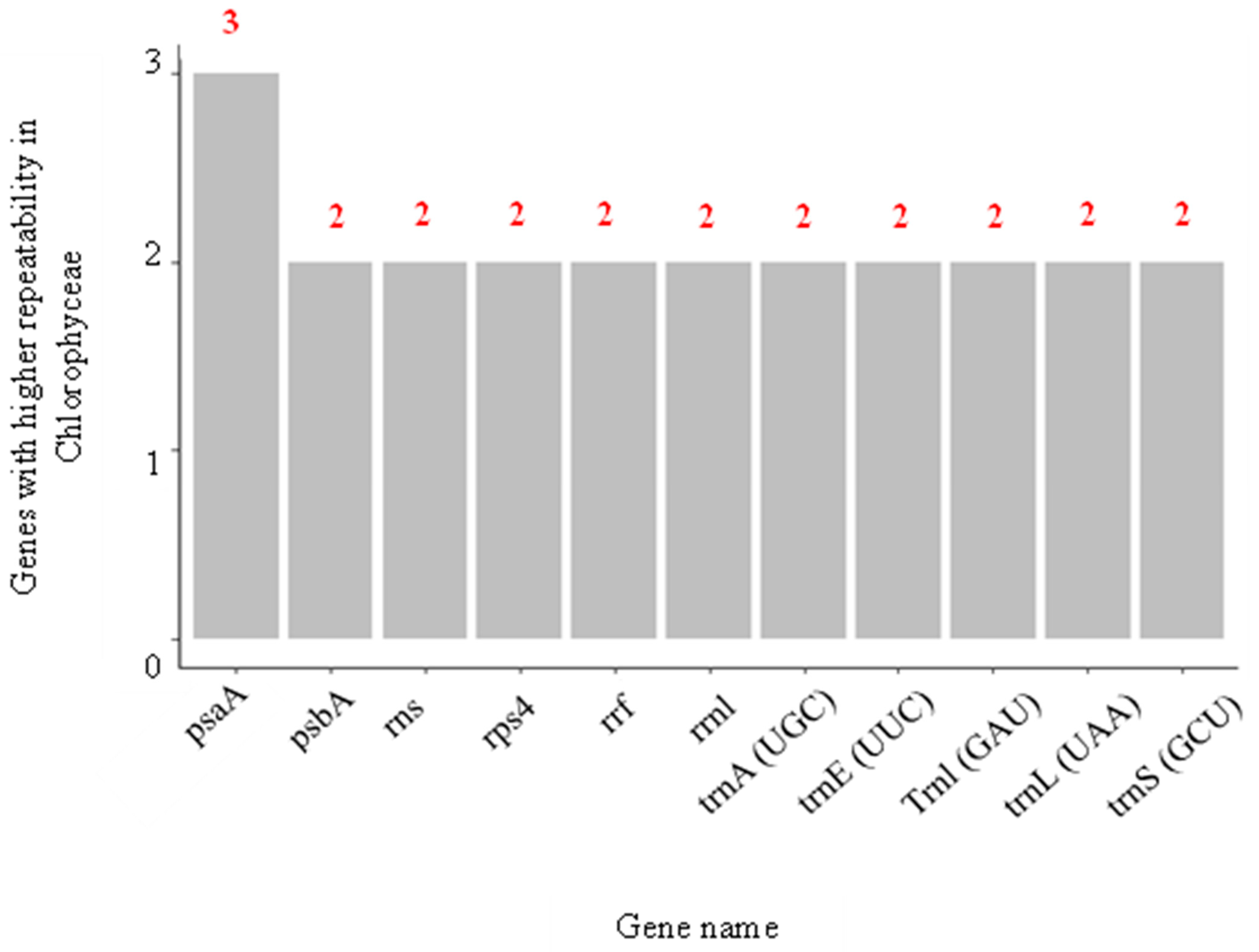
3.1.3. Co-Occurrences Analysis
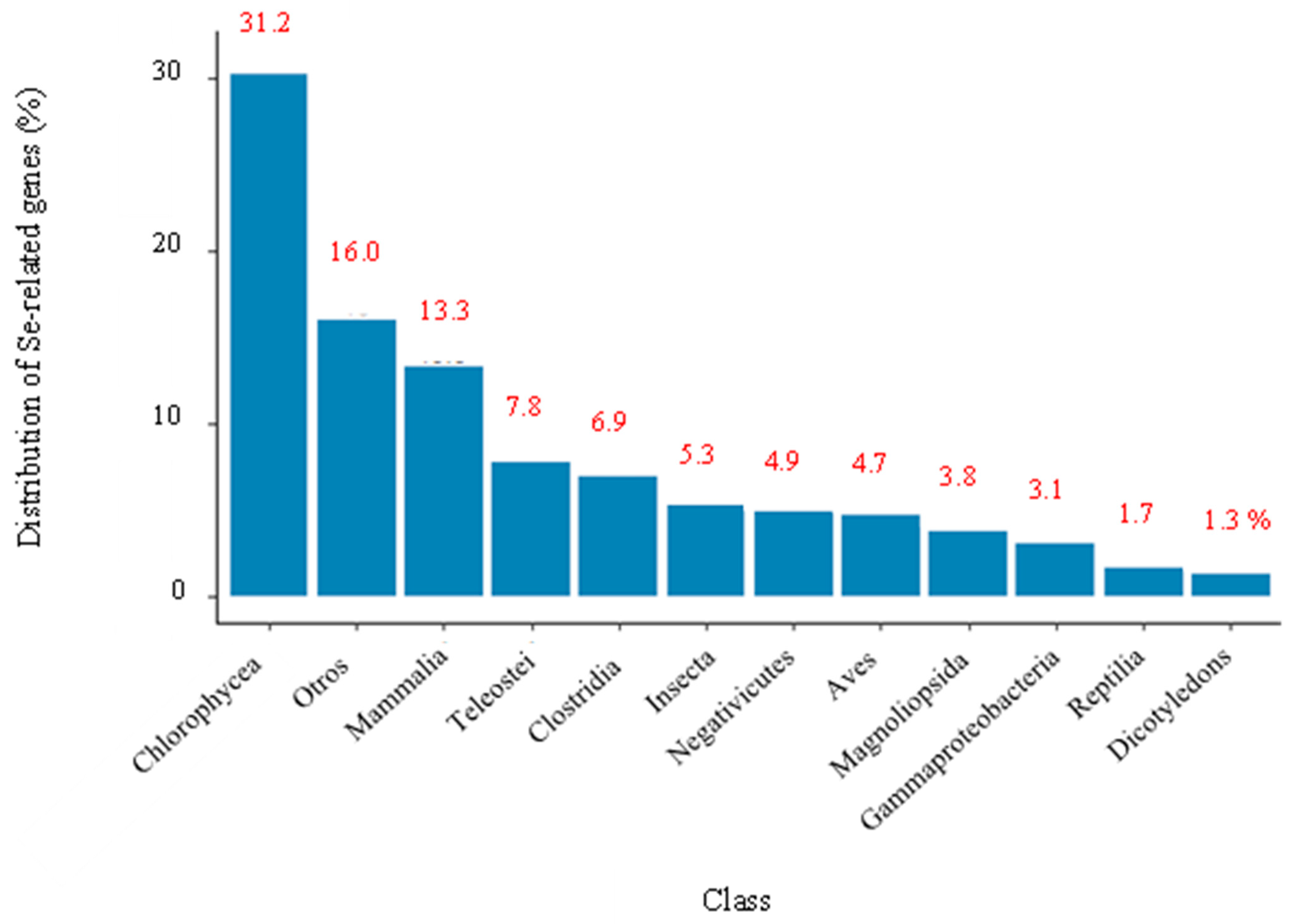
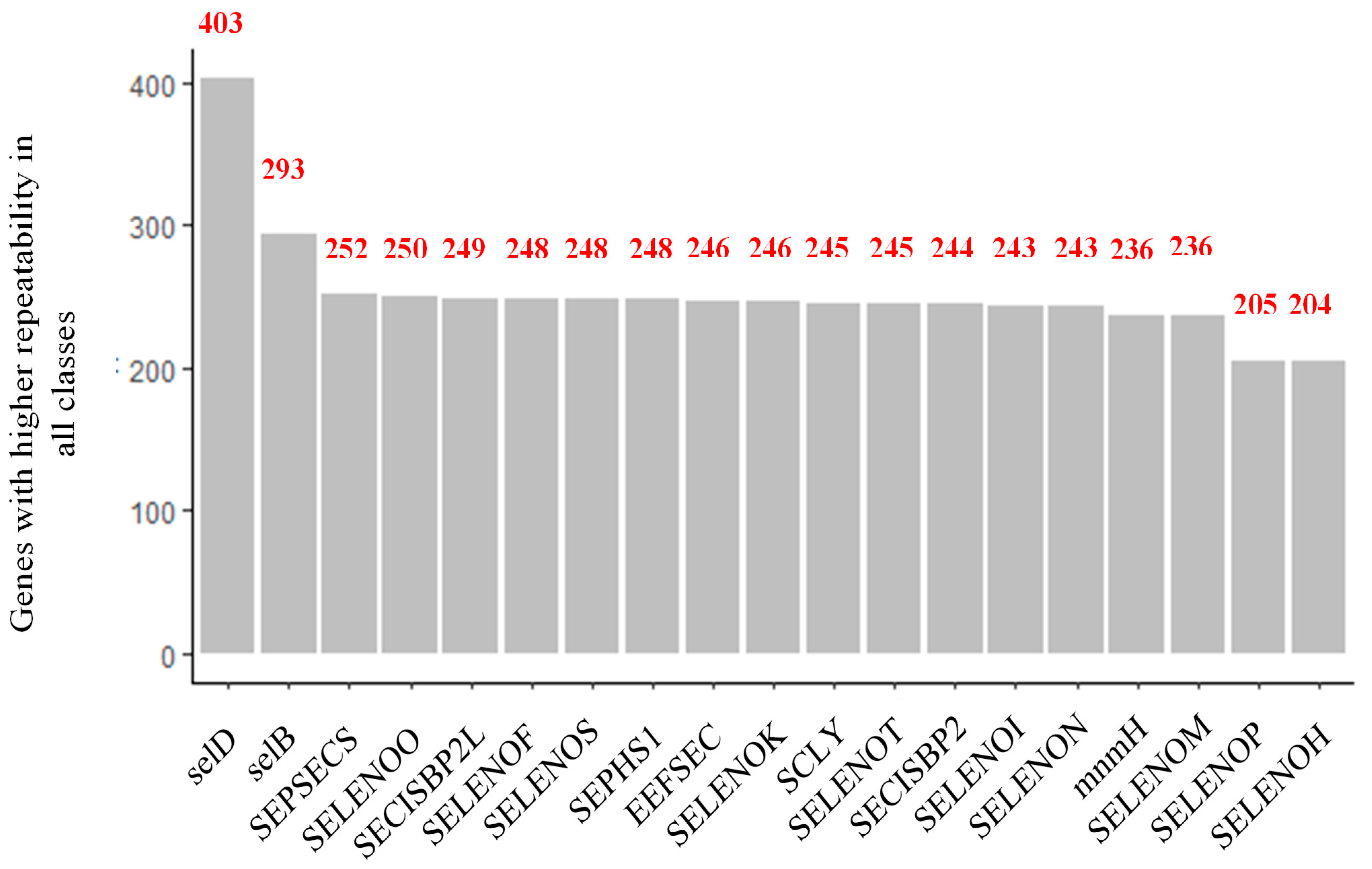
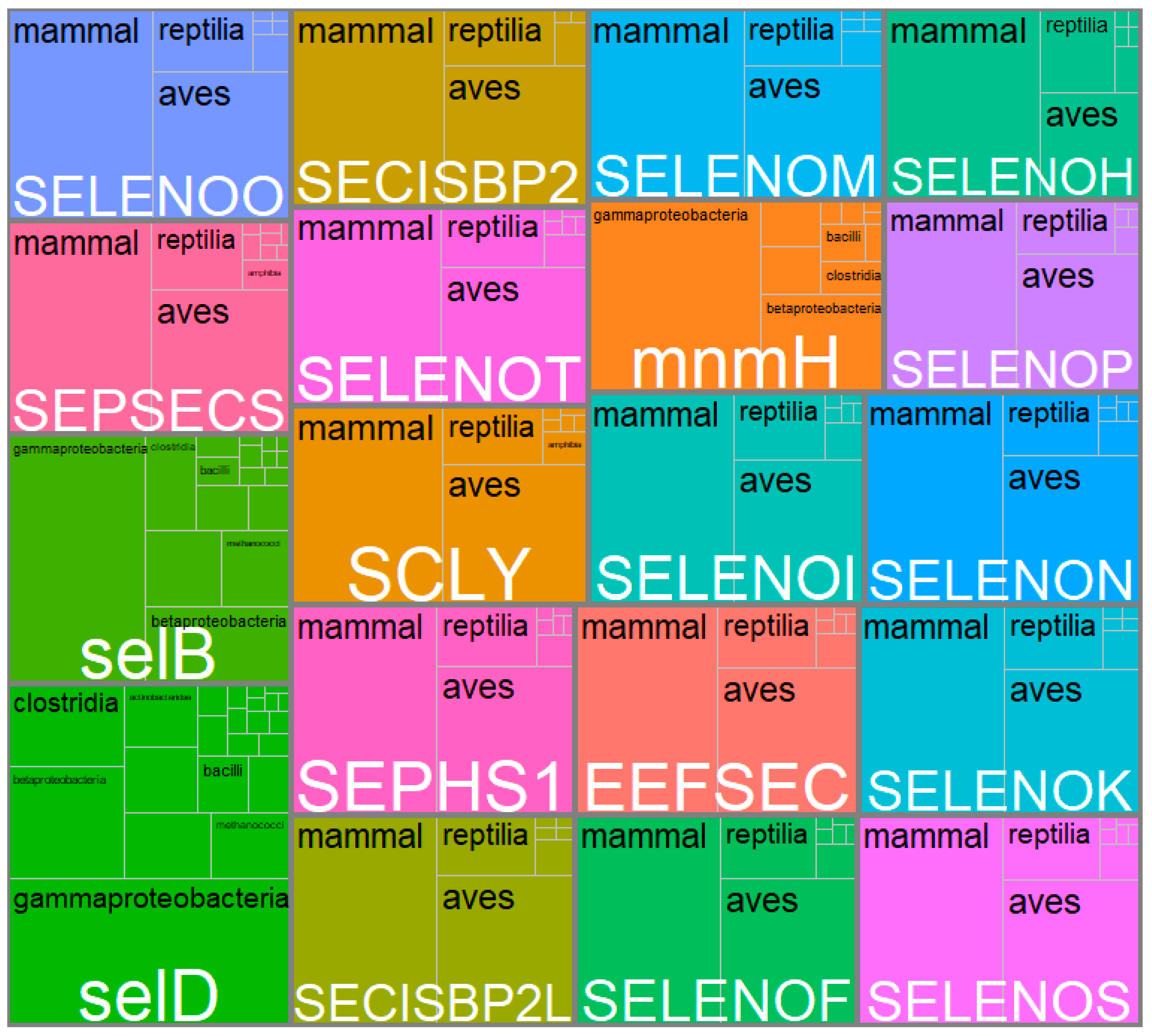
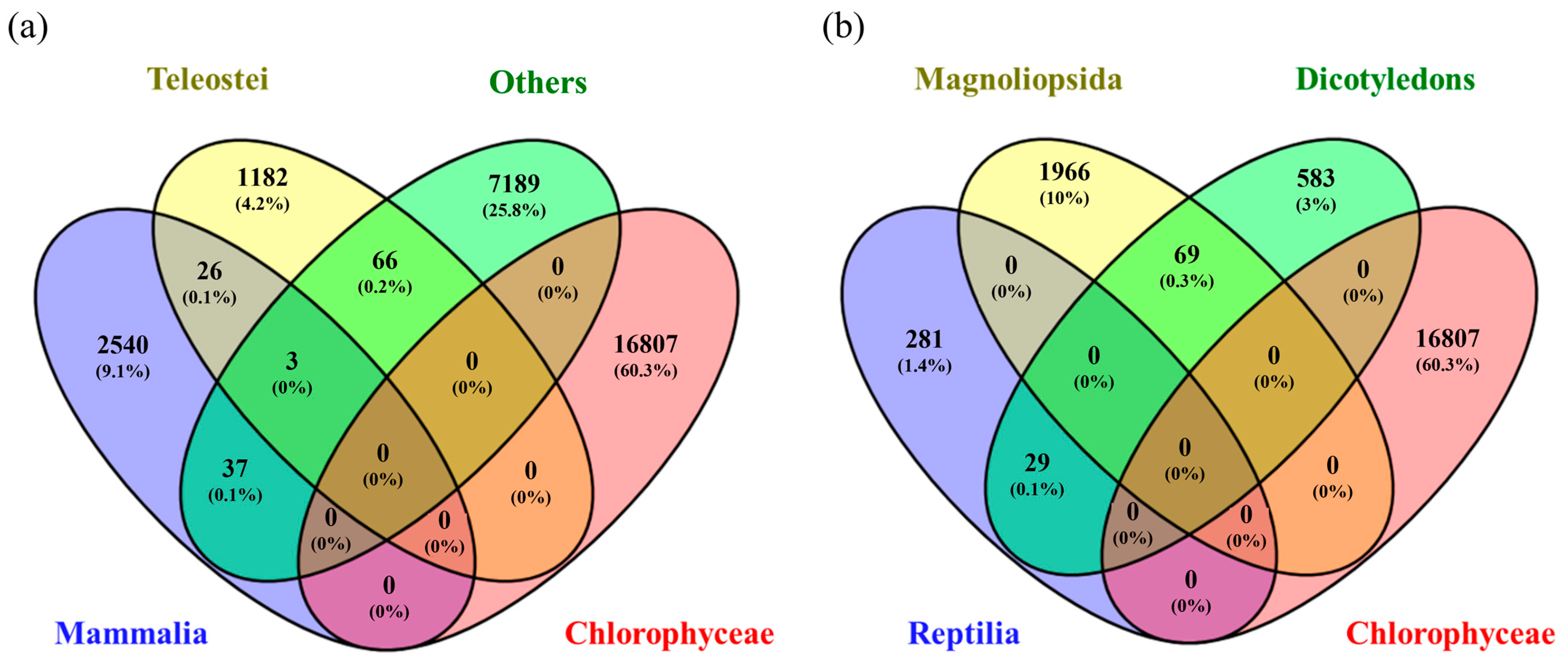
3.2. Systematic Analysis of Genes Related to Selenium Metabolism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Biological Function | Reference |
|---|---|---|
| SelD | This selenophosphate synthetase is highly specific for selenide, capable of discriminating sulfur. It has even been associated with regulating cell proliferation, growth, and differentiation. | [25,26] |
| SelB | It is a specialized translation elongation factor. Furthermore, it is an enzyme required for synthesizing and inserting selenocysteine into proteins. | [27] |
| SEPSECS | It provides instructions for making an O-phosphoseryl-tRNA (Sec) enzyme, which forms tRNAs critical to selenocysteine production. | [22] |
| SELENOO | It codes for a selenoprotein located in mitochondria and is known to be the largest selenocysteine-containing mammalian selenoprotein. However, the exact function of this is unknown, but it is believed to have redox activity. | [28] |
| SECISBP2L | It interacts with all known human SECIS (Sec insertion sequence) in vitro RNAs. Furthermore, selenoprotein mRNAs are co-immunoprecipitated with endogenous SBP2L, suggesting a role in regulating selenoprotein expression, but does not promote Sec incorporation into in vitro selenoproteins. | [29] |
| SELENOF | Due to S or Se, it could be involved in redox reactions associated with oxidative stress of the endoplasmic reticulum (ER) and carcinogenesis. In addition, it contributes to the quality control of protein folding in the ER. However, its biological function is still unclear. | [30] |
| SELENOS | It regulates inflammation, oxidative stress, and endoplasmic reticulum (ER) stress. It is also involved in the degradation process of misfolded proteins in the ER and could function as a modulator of Sec insertion. | [31] |
| SEPHS1 | It encodes an enzyme that synthesizes selenophosphate from selenide and ATP. Selenophosphate is the selenium donor used to synthesize selenocysteine. | [32] |
| EEFSEC | It is a necessary translation factor for the incorporation of selenocysteine into proteins. | [28] |
| SELENOK | It is a transmembrane protein that participates in the ER-associated degradation of misfolded glycosylated proteins. It also has a role in protecting cells from stress-induced apoptosis in the ER. Furthermore, knockout studies in mice show the importance of this gene in promoting Ca2+ flux in immune cells and generating an effective immune response. | [33,34] |
| SCLY | Catalyzes the breakdown of L-selenocysteine into L-alanine and elemental selenium. | [35] |
| SELENOT | Studies in mice indicate a crucial role for this gene in the protection of dopaminergic neurons against oxidative stress in Parkinson’s disease and the control of glucose homeostasis in pancreatic beta cells. | [36] |
| SECISBP2 | An essential component of the machinery involved in the co-translational insertion of selenocysteine (Sec) into selenoproteins. Mutations in this gene have been associated with reduced iodothyronine deiodinase type II enzyme activity (a selenoprotein) and abnormal thyroid hormone metabolism. | [37] |
| SELENOI | It is a multi-passage transmembrane protein. It catalyzes the transfer of phosphoethanolamine from CDP-ethanolamine to diacylglycerol to produce phosphatidylethanolamine, which is involved in forming and maintaining vesicular membranes, regulation of lipid metabolism, and protein folding. | [28] |
| SELENON | It plays an essential role in cell protection against oxidative stress and regulating calcium homeostasis in the ER related to redox activity. | [24] |
| mnmH | It codes for a selenophosphate-dependent tRNA 2-selenouridine synthase, essential for modifying some tRNAs to replace a sulfur atom with selenium. This enzyme works with SelD, the selenium donor protein, which also acts on the incorporation of selenocysteine. | [29] |
| SELENOM | Located in the perinuclear region, it is highly expressed in the brain and may be involved in neurodegenerative disorders. Studies have shown that transgenic mice with targeted deletion of this gene exhibit weight gain. That suggests a role for this gene in regulating body weight and energy metabolism. | [38,39] |
| SELENOP | It codes for a selenoprotein predominantly expressed in the liver and secreted into the plasma. This selenoprotein is unique because it contains multiple selenocysteine (Sec) residues per polypeptide (ten in humans) and accounts for most of the selenium in plasma. Mice lacking this gene exhibit neurological dysfunction. | [29] |
| SELENOH | Codes for a nucleolar selenoprotein. It functions as an oxidoreductase and has been shown to protect neurons against UVB-induced damage by inhibiting apoptotic cell death pathways. Promotes mitochondrial biogenesis and function and suppresses cellular senescence through genome maintenance and redox regulation. | [40] |
3.3. Selenium Accumulation and Metabolism
4. Conclusions and Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hefnawy, A.E.G.; Tórtora-Pérez, J.L. The Importance of Selenium and the Effects of Its Deficiency in Animal Health. Small Rumin. Res. 2010, 89, 185–192. [Google Scholar] [CrossRef]
- Yang, H.; Yang, X.; Ning, Z.; Kwon, S.Y.; Li, M.L.; Tack, F.M.G.; Kwon, E.E.; Rinklebe, J.; Yin, R. The Beneficial and Hazardous Effects of Selenium on the Health of the Soil-Plant-Human System: An Overview. J. Hazard Mater. 2022, 422, 126876. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and Human Health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Pieniz, S.; Andreazza, R.; Pereira, J.Q.; de Oliveira Camargo, F.A.; Brandelli, A. Production of Selenium-Enriched Biomass by Enterococcus Durans. Biol. Trace Elem. Res. 2013, 155, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zheng, W.; Wong, Y.S.; Yang, F.; Bai, Y. Accumulation of Selenium in Mixotrophic Culture of Spirulina Platensis on Glucose. Bioresour. Technol. 2006, 97, 2260–2265. [Google Scholar] [CrossRef]
- Guimarães, B.O.; de Boer, K.; Gremmen, P.; Drinkwaard, A.; Wieggers, R.; Wijffels, R.H.; Barbosa, M.J.; D’Adamo, S. Selenium Enrichment in the Marine Microalga Nannochloropsis Oceanica. Algal Res. 2021, 59, 102427. [Google Scholar] [CrossRef]
- Bouain, N.; Krouk, G.; Lacombe, B.; Rouached, H. Getting to the Root of Plant Mineral Nutrition: Combinatorial Nutrient Stresses Reveal Emergent Properties. Trends Plant Sci. 2019, 24, 542–552. [Google Scholar] [CrossRef]
- Ekumah, J.N.; Ma, Y.; Akpabli-Tsigbe, N.D.K.; Kwaw, E.; Ma, S.; Hu, J. Global Soil Distribution, Dietary Access Routes, Bioconversion Mechanisms and the Human Health Significance of Selenium: A Review. Food Biosci. 2021, 41, 100960. [Google Scholar] [CrossRef]
- Gojkovic, Ž.; Garbayo, I.; Ariza, J.L.G.; Márová, I.; Vílchez, C. Selenium Bioaccumulation and Toxicity in Cultures of Green Microalgae. Algal Res. 2015, 7, 106–116. [Google Scholar] [CrossRef]
- de Morais, E.G.; Murillo, A.M.; Lens, P.N.L.; Ferrer, I.; Uggetti, E. Selenium Recovery from Wastewater by the Green Microalgae Chlorella vulgaris and Scenedesmus sp. Sci. Total Environ. 2022, 851, 158337. [Google Scholar] [CrossRef]
- Gać, P.; Czerwińska, K.; Macek, P.; Jaremków, A.; Mazur, G.; Pawlas, K.; Poręba, R. The Importance of Selenium and Zinc Deficiency in Cardiovascular Disorders. Environ. Toxicol. Pharmacol. 2021, 82, 103553. [Google Scholar] [CrossRef] [PubMed]
- Rua, R.M.; Nogales, F.; Carreras, O.; Ojeda, M.L. Selenium, Selenoproteins and Cancer of the Thyroid. J. Trace Elem. Med. Biol. 2023, 76, 127115. [Google Scholar] [CrossRef] [PubMed]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef]
- Vu, D.L.; Saurav, K.; Mylenko, M.; Ranglová, K.; Kuta, J.; Ewe, D.; Masojídek, J.; Hrouzek, P. In Vitro Bioaccessibility of Selenoamino Acids from Selenium (Se)-Enriched Chlorella vulgaris Biomass in Comparison to Selenized Yeast; a Se-Enriched Food Supplement; and Se-Rich Foods. Food Chem. 2019, 279, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Zhang, J.; Leng, X.; Fan, Y.; Fu, B.; Wang, P. A Scientometrics Analysis and Visualization of Low Back Pain. Int. J. Osteopath. Med. 2023, 47, 100655. [Google Scholar] [CrossRef]
- Miranda, A.M.; Hernandez-Tenorio, F.; Ocampo, D.; Vargas, G.J.; Sáez, A.A. Trends on CO2 Capture with Microalgae: A Bibliometric Analysis. Molecules 2022, 27, 4669. [Google Scholar] [CrossRef]
- Hernandez-Tenorio, F.; Miranda, A.M.; Rodríguez, C.A.; Giraldo-Estrada, C.; Sáez, A.A. Potential Strategies in the Biopesticide Formulations: A Bibliometric Analysis. Agronomy 2022, 12, 2665. [Google Scholar] [CrossRef]
- Ubando, A.T.; Africa, A.D.M.; Maniquiz-Redillas, M.C.; Culaba, A.B.; Chen, W.H.; Chang, J.S. Microalgal Biosorption of Heavy Metals: A Comprehensive Bibliometric Review. J. Hazard Mater. 2021, 402, 123431. [Google Scholar] [CrossRef]
- El Gamal, A.A. Biological Importance of Marine Algae. Saudi Pharm. J. 2010, 18, 1–25. [Google Scholar] [CrossRef]
- Baker, R.D.; Baker, S.S.; LaRosa, K.; Whitney, C.; Newburger, P.E. Selenium Regulation of Glutathione Peroxidase in Human Hepatoma Cell Line Hep3B. Arch. Biochem. Biophys. 1993, 304, 53–57. [Google Scholar] [CrossRef]
- Madeja, Z.; Sroka, J.; Nyström, C.; Björkhem-Bergman, L.; Nordman, T.; Damdimopoulos, A.; Nalvarte, I.; Eriksson, L.C.; Spyrou, G.; Olsson, J.M.; et al. The Role of Thioredoxin Reductase Activity in Selenium-Induced Cytotoxicity. Biochem. Pharmacol. 2005, 69, 1765–1772. [Google Scholar] [CrossRef] [PubMed]
- UniProt. 2022. Available online: https://www.uniprot.org/ (accessed on 10 December 2022).
- Caţianis, A.G.; Virgolici, B.; Dogaru, B.C.; Virgolici, H.; Mohora, M. From Selenium to Selenoproteins and Their Role—Minireview. Acta Medica Transilvanica 2020, 25, 56–59. [Google Scholar] [CrossRef]
- Pitts, M.W.; Hoffmann, P.R. Endoplasmic Reticulum-Resident Selenoproteins as Regulators of Calcium Signaling and Homeostasis. Cell Calcium. 2018, 70, 76. [Google Scholar] [CrossRef] [PubMed]
- Lacourciere, G.M. Biosynthesis of Selenophosphate. BioFactors 1999, 10, 237–244. [Google Scholar] [CrossRef]
- Serras, F.; Morey, M.; Alsina, B.; Baguñà, J.; Corominas, M. The Drosophilaselenophosphate Synthetase (SelD) Gene Is Required for Development and Cell Proliferation. BioFactors 2001, 14, 143–149. [Google Scholar] [CrossRef]
- Wolfe, M.D. Selenophosphate Synthetase. EcoSal. Plus 2004, 1, 1. [Google Scholar] [CrossRef]
- Selenoo Protein Expression Summary. Available online: https://www.proteinatlas.org/ENSG00000073169-SELENOO (accessed on 12 December 2022).
- NCBI. 2022. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 11 December 2022).
- Tian, J.; Liu, J.; Li, J.; Zheng, J.; Chen, L.; Wang, Y.; Liu, Q.; Ni, J. The Interaction of Selenoprotein F (SELENOF) with Retinol Dehydrogenase 11 (RDH11) Implied a Role of SELENOF in Vitamin A Metabolism. Nutr. Metab 2018, 15, 7. [Google Scholar] [CrossRef]
- Yu, S.; Du, J. ling. Selenoprotein S: A Therapeutic Target for Diabetes and Macroangiopathy? Cardiovasc. Diabetol. 2017, 16, 101. [Google Scholar] [CrossRef]
- Tobe, R.; Carlson, B.A.; Huh, J.H.; Castro, N.P.; Xu, X.M.; Tsuji, P.A.; Lee, S.G.; Bang, J.; Na, J.W.; Kong, Y.Y.; et al. Selenophosphate Synthetase 1 Is an Essential Protein with Roles in Regulation of Redox Homoeostasis in Mammals. Biochem. J. 2016, 473, 2141–2154. [Google Scholar] [CrossRef]
- Gonzalez-Flores, J.N.; Gupta, N.; DeMong, L.W.; Copeland, P.R. The Selenocysteine-Specific Elongation Factor Contains a Novel and Multi-Functional Domain. J. Biol. Chem. 2012, 287, 38936–38945. [Google Scholar] [CrossRef]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The Role of Selenium in Inflammation and Immunity: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [PubMed]
- Seale, L.A. Selenocysteine β-Lyase: Biochemistry, Regulation and Physiological Role of the Selenocysteine Decomposition Enzyme. Antioxidants 2019, 8, 357. [Google Scholar] [CrossRef] [PubMed]
- Alsharif, I.; Boukhzar, L.; Lefranc, B.; Godefroy, D.; Aury-Landas, J.; do Rego, J.L.; do Rego, J.C.; Naudet, F.; Arabo, A.; Chagraoui, A.; et al. Cell-Penetrating, Antioxidant SELENOT Mimetic Protects Dopaminergic Neurons and Ameliorates Motor Dysfunction in Parkinson’s Disease Animal Models. Redox. Biol. 2021, 40, 101839. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Korwutthikulrangsri, M.; Gönç, E.N.; Sillers, L.; Liao, X.H.; Alikaşifoǧlu, A.; Kandemir, N.; Menucci, M.B.; Burman, K.D.; Weiss, R.E.; et al. Clinical and Molecular Analysis in 2 Families with Novel Compound Heterozygous SBP2 (SECISBP2) Mutations. J. Clin. Endocrinol. Metab. 2020, 105, e6–e11. [Google Scholar] [CrossRef] [PubMed]
- Lanza, M.G.D.B.; dos Reis, A.R. Roles of Selenium in Mineral Plant Nutrition: ROS Scavenging Responses against Abiotic Stresses. Plant Physiol. Biochem. 2021, 164, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Pitts, M.W.; Reeves, M.A.; Hashimoto, A.C.; Ogawa, A.; Kremer, P.; Seale, L.A.; Berry, M.J. Deletion of Selenoprotein M Leads to Obesity without Cognitive Deficits. J. Biol. Chem. 2013, 288, 26121–26134. [Google Scholar] [CrossRef]
- Mendelev, N.; Mehta, S.L.; Witherspoon, S.; He, Q.; Sexton, J.Z.; Li, P.A. Upregulation of Human Selenoprotein H in Murine Hippocampal Neuronal Cells Promotes Mitochondrial Biogenesis and Functional Performance. Mitochondrion 2011, 11, 76–82. [Google Scholar] [CrossRef]
- Zhu, Q.L.; Guo, S.N.; Wen, F.; Zhang, X.L.; Wang, C.C.; Si, L.F.; Zheng, J.L.; Liu, J. Transcriptional and Physiological Responses of Dunaliella salina to Cadmium Reveals Time-Dependent Turnover of Ribosome, Photosystem, and ROS-Scavenging Pathways. Aquat. Toxicol. 2019, 207, 153–162. [Google Scholar] [CrossRef]
- Kumar, G.; Shekh, A.; Jakhu, S.; Sharma, Y.; Kapoor, R.; Sharma, T.R. Bioengineering of Microalgae: Recent Advances, Perspectives, and Regulatory Challenges for Industrial Application. Front. Bioeng. Biotechnol. 2020, 8, 914. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Parrasia, S.; Vecchia, F.D. Selenium Accumulation and Metabolism in Algae. Aquat. Toxicol. 2017, 189, 1–8. [Google Scholar] [CrossRef]
- Araie, H.; Shiraiwa, Y. Selenium Utilization Strategy by Microalgae. Molecules 2009, 14, 4880–4891. [Google Scholar] [CrossRef] [PubMed]
- Novoselov, S.V.; Rao, M.; Onoshko, N.V.; Zhi, H.; Kryukov, G.V.; Xiang, Y.; Weeks, D.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins and Selenocysteine Insertion System in the Model Plant Cell System, Chlamydomonas reinhardtii. EMBO J. 2002, 21, 3681–3693. [Google Scholar] [CrossRef] [PubMed]
- Morlon, H.; Fortin, C.; Adam, C.; Garnier-Laplace, J. Selenite Transport and Its Inhibition in the Unicellular Green Alga Chlamydomonas reinhardtii. Environ. Toxicol. Chem. 2006, 25, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Tao, M.; Li, Z.; Hu, Z. Cloning and Characterization of Selenoprotein Thioredoxin Reductase Gene in Haematococcus pluvialis. Algal Res. 2016, 17, 97–104. [Google Scholar] [CrossRef]
- Ponton, D.E.; Graves, S.D.; Fortin, C.; Janz, D.; Amyot, M.; Schiavon, M. Selenium Interactions with Algae: Chemical Processes at Biological Uptake Sites, Bioaccumulation, and Intracellular Metabolism. Plants 2020, 9, 528. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoyos, B.S.; Hernandez-Tenorio, F.; Miranda, A.M.; Villanueva-Mejía, D.F.; Sáez, A.A. Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review. Biology 2023, 12, 703. https://doi.org/10.3390/biology12050703
Hoyos BS, Hernandez-Tenorio F, Miranda AM, Villanueva-Mejía DF, Sáez AA. Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review. Biology. 2023; 12(5):703. https://doi.org/10.3390/biology12050703
Chicago/Turabian StyleHoyos, Brenda S., Fabian Hernandez-Tenorio, Alejandra M. Miranda, Diego F. Villanueva-Mejía, and Alex A. Sáez. 2023. "Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review" Biology 12, no. 5: 703. https://doi.org/10.3390/biology12050703
APA StyleHoyos, B. S., Hernandez-Tenorio, F., Miranda, A. M., Villanueva-Mejía, D. F., & Sáez, A. A. (2023). Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review. Biology, 12(5), 703. https://doi.org/10.3390/biology12050703