

Sepsis-like Energy Deficit Is Not Sufficient to Induce Early Muscle Fiber Atrophy and Mitochondrial Dysfunction in a Murine Sepsis Model

 , , , , , and
, , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
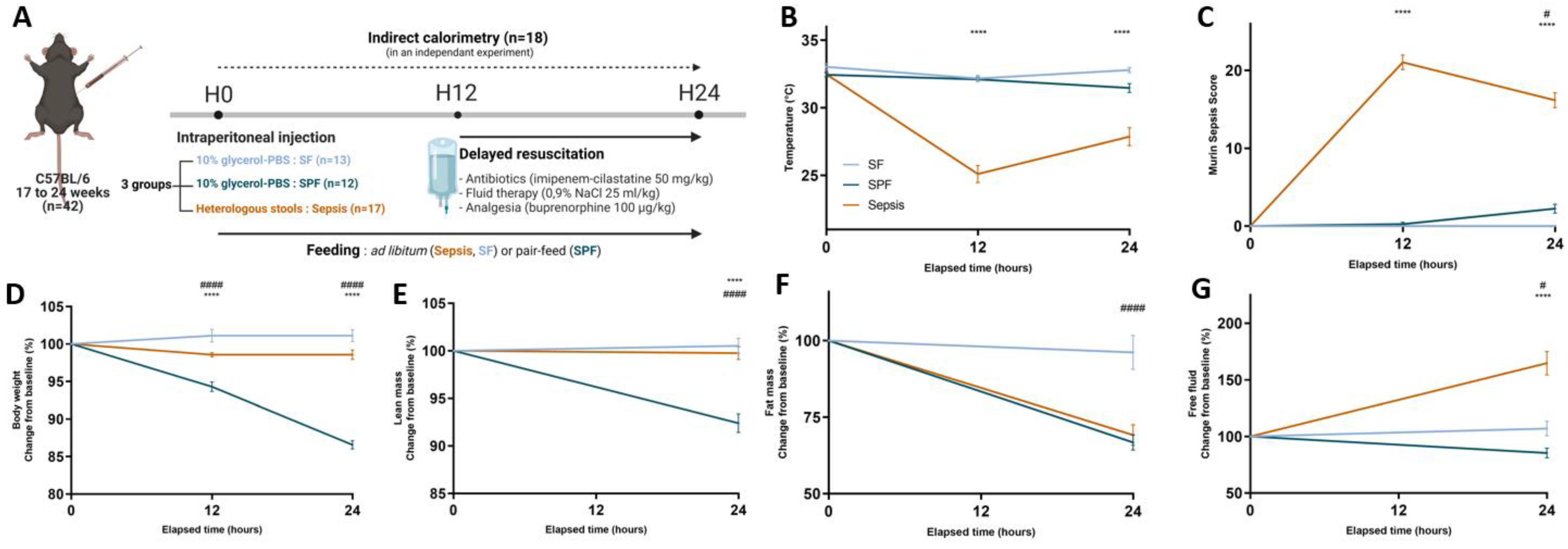
2.1. Murine Model of Resuscitated Sepsis
2.2. Metabolic Study
2.3. High-Resolution Respirometry on Permeabilized Muscle Fibers
2.4. Western Blot
2.5. Relative Gene Expression
2.6. Mitochondrial DNA Copy Number
2.7. Muscle Mass and Fibers’ Cross-Sectional Area
2.8. Statistical Analysis
3. Results
3.1. Sepsis-Induced Changes in Body Composition Were Not Mediated by Decreased Energy Intake Alone
3.2. Early Energy Deficit Was Identical between Sepsis and SPF Mice
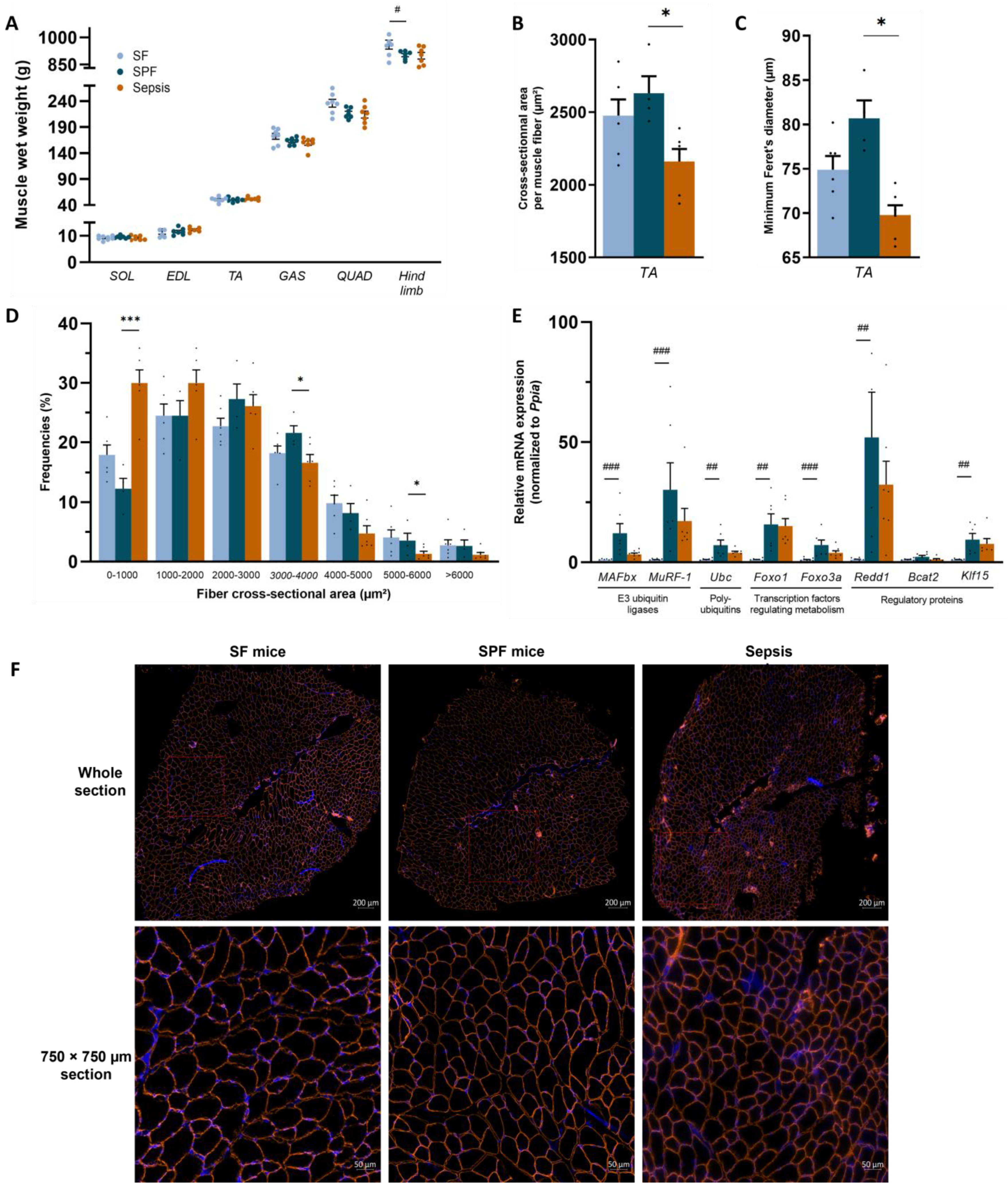
3.3. Sepsis-like Energy Deficit Itself Was Not Sufficient to Induce Muscle Fiber Atrophy
3.4. Sepsis-like Energy Deficit Was Not Sufficient Enough to Induce Mitochondrial Dysfunction in Skeletal Muscle
3.5. Sepsis-like Energy Deficit Induced Specific MtQC Adaptations Not Found in Sepsis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801. [Google Scholar] [CrossRef] [PubMed]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, Regional, and National Sepsis Incidence and Mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Hermans, G.; Van Mechelen, H.; Clerckx, B.; Vanhullebusch, T.; Mesotten, D.; Wilmer, A.; Casaer, M.P.; Meersseman, P.; Debaveye, Y.; Van Cromphaut, S.; et al. Acute Outcomes and 1-Year Mortality of Intensive Care Unit-Acquired Weakness. A Cohort Study and Propensity-Matched Analysis. Am. J. Respir. Crit. Care Med. 2014, 190, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Van Aerde, N.; Meersseman, P.; Debaveye, Y.; Wilmer, A.; Gunst, J.; Casaer, M.P.; Bruyninckx, F.; Wouters, P.J.; Gosselink, R.; Van den Berghe, G.; et al. Five-Year Impact of ICU-Acquired Neuromuscular Complications: A Prospective, Observational Study. Intensive Care Med. 2020, 46, 1184–1193. [Google Scholar] [CrossRef] [PubMed]
- Vanhorebeek, I.; Latronico, N.; Van den Berghe, G. ICU-Acquired Weakness. Intensive Care Med. 2020, 46, 637–653. [Google Scholar] [CrossRef] [PubMed]
- Voiriot, G.; Oualha, M.; Pierre, A.; Salmon-Gandonnière, C.; Gaudet, A.; Jouan, Y.; Kallel, H.; Radermacher, P.; Vodovar, D.; Sarton, B.; et al. Chronic Critical Illness and Post-Intensive Care Syndrome: From Pathophysiology to Clinical Challenges. Ann. Intensive Care 2022, 12, 58. [Google Scholar] [CrossRef]
- Fredriksson, K.; Hammarqvist, F.; Strigård, K.; Hultenby, K.; Ljungqvist, O.; Wernerman, J.; Rooyackers, O. Derangements in Mitochondrial Metabolism in Intercostal and Leg Muscle of Critically Ill Patients with Sepsis-Induced Multiple Organ Failure. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1044–E1050. [Google Scholar] [CrossRef]
- Fredriksson, K.; Tjäder, I.; Keller, P.; Petrovic, N.; Ahlman, B.; Schéele, C.; Wernerman, J.; Timmons, J.A.; Rooyackers, O. Dysregulation of Mitochondrial Dynamics and the Muscle Transcriptome in ICU Patients Suffering from Sepsis Induced Multiple Organ Failure. PLoS ONE 2008, 3, e3686. [Google Scholar] [CrossRef]
- Jiroutková, K.; Krajčová, A.; Ziak, J.; Fric, M.; Waldauf, P.; Džupa, V.; Gojda, J.; Němcova-Fürstová, V.; Kovář, J.; Elkalaf, M.; et al. Mitochondrial Function in Skeletal Muscle of Patients with Protracted Critical Illness and ICU-Acquired Weakness. Crit. Care 2015, 19, 448. [Google Scholar] [CrossRef]
- Brealey, D.; Brand, M.; Hargreaves, I.; Heales, S.; Land, J.; Smolenski, R.; Davies, N.A.; Cooper, C.E.; Singer, M. Association between Mitochondrial Dysfunction and Severity and Outcome of Septic Shock. Lancet 2002, 360, 219–223. [Google Scholar] [CrossRef]
- Preau, S.; Vodovar, D.; Jung, B.; Lancel, S.; Zafrani, L.; Flatres, A.; Oualha, M.; Voiriot, G.; Jouan, Y.; Joffre, J.; et al. Energetic Dysfunction in Sepsis: A Narrative Review. Ann. Intensive Care 2021, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Popov, L. Mitochondrial Biogenesis: An Update. J. Cell Mol. Med. 2020, 24, 4892–4899. [Google Scholar] [CrossRef] [PubMed]
- Losón, O.C.; Song, Z.; Chen, H.; Chan, D.C. Fis1, Mff, MiD49, and MiD51 Mediate Drp1 Recruitment in Mitochondrial Fission. Mol. Biol. Cell 2013, 24, 659–667. [Google Scholar] [CrossRef]
- Palmer, C.S.; Elgass, K.D.; Parton, R.G.; Osellame, L.D.; Stojanovski, D.; Ryan, M.T. Adaptor Proteins MiD49 and MiD51 Can Act Independently of Mff and Fis1 in Drp1 Recruitment and Are Specific for Mitochondrial Fission. J. Biol. Chem. 2013, 288, 27584–27593. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, M.; Singh, S.; Singh, A.P.; Dasgupta, S. Mitochondrial Fusion and Fission: The Fine-Tune Balance for Cellular Homeostasis. FASEB J. 2021, 35, e21620. [Google Scholar] [CrossRef] [PubMed]
- Patten, D.A.; Wong, J.; Khacho, M.; Soubannier, V.; Mailloux, R.J.; Pilon-Larose, K.; MacLaurin, J.G.; Park, D.S.; McBride, H.M.; Trinkle-Mulcahy, L.; et al. OPA1-Dependent Cristae Modulation Is Essential for Cellular Adaptation to Metabolic Demand. EMBO J. 2014, 33, 2676–2691. [Google Scholar] [CrossRef] [PubMed]
- Durcan, T.M.; Fon, E.A. The Three ’P’s of Mitophagy: PARKIN, PINK1, and Post-Translational Modifications. Genes Dev. 2015, 29, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. P62 Links the Autophagy Pathway and the Ubiqutin–Proteasome System upon Ubiquitinated Protein Degradation. Cell. Mol. Biol. Lett. 2016, 21, 29. [Google Scholar] [CrossRef]
- Villa, E.; Marchetti, S.; Ricci, J.-E. No Parkin Zone: Mitophagy without Parkin. Trends Cell Biol. 2018, 28, 882–895. [Google Scholar] [CrossRef]
- Trefts, E.; Shaw, R.J. AMPK: Restoring Metabolic Homeostasis over Space and Time. Mol. Cell 2021, 81, 3677–3690. [Google Scholar] [CrossRef]
- Mehrabani, S.; Bagherniya, M.; Askari, G.; Read, M.I.; Sahebkar, A. The Effect of Fasting or Calorie Restriction on Mitophagy Induction: A Literature Review. J. Cachexia Sarcopenia Muscle 2020, 11, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Singer, P.; Blaser, A.R.; Berger, M.M.; Alhazzani, W.; Calder, P.C.; Casaer, M.P.; Hiesmayr, M.; Mayer, K.; Montejo, J.C.; Pichard, C.; et al. ESPEN Guideline on Clinical Nutrition in the Intensive Care Unit. Clin. Nutr. 2019, 38, 48–79. [Google Scholar] [CrossRef] [PubMed]
- Occhiali, E.; Urli, M.; Pressat-Laffouilhère, T.; Achamrah, N.; Veber, B.; Clavier, T. Dynamic Metabolic Changes Measured by Indirect Calorimetry during the Early Phase of Septic Shock: A Prospective Observational Pilot Study. Eur. J. Clin. Nutr. 2022, 76, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Carbone, J.W.; Pasiakos, S.M.; Vislocky, L.M.; Anderson, J.M.; Rodriguez, N.R. Effects of Short-Term Energy Deficit on Muscle Protein Breakdown and Intramuscular Proteolysis in Normal-Weight Young Adults. Appl. Physiol. Nutr. Metab. 2014, 39, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Preau, S.; Ambler, M.; Sigurta, A.; Kleyman, A.; Dyson, A.; Hill, N.E.; Boulanger, E.; Singer, M. Protein Recycling and Limb Muscle Recovery after Critical Illness in Slow- and Fast-Twitch Limb Muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R584–R593. [Google Scholar] [CrossRef]
- Rubinson, L.; Diette, G.B.; Song, X.; Brower, R.G.; Krishnan, J.A. Low Caloric Intake Is Associated with Nosocomial Bloodstream Infections in Patients in the Medical Intensive Care Unit. Crit. Care Med. 2004, 32, 350–357. [Google Scholar] [CrossRef]
- Villet, S.; Chiolero, R.L.; Bollmann, M.D.; Revelly, J.-P.; Rn, M.-C.C.; Delarue, J.; Berger, M.M. Negative Impact of Hypocaloric Feeding and Energy Balance on Clinical Outcome in ICU Patients. Clin. Nutr. 2005, 24, 502–509. [Google Scholar] [CrossRef]
- Alberda, C.; Gramlich, L.; Jones, N.; Jeejeebhoy, K.; Day, A.G.; Dhaliwal, R.; Heyland, D.K. The Relationship between Nutritional Intake and Clinical Outcomes in Critically Ill Patients: Results of an International Multicenter Observational Study. Intensive Care Med. 2009, 35, 1728–1737. [Google Scholar] [CrossRef]
- Carbone, J.W.; McClung, J.P.; Pasiakos, S.M. Skeletal Muscle Responses to Negative Energy Balance: Effects of Dietary Protein. Adv. Nutr. 2012, 3, 119–126. [Google Scholar] [CrossRef]
- Xu, Z.; Fu, T.; Guo, Q.; Zhou, D.; Sun, W.; Zhou, Z.; Chen, X.; Zhang, J.; Liu, L.; Xiao, L.; et al. Disuse-Associated Loss of the Protease LONP1 in Muscle Impairs Mitochondrial Function and Causes Reduced Skeletal Muscle Mass and Strength. Nat. Commun. 2022, 13, 894. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal Muscle Energy Metabolism during Exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Starr, M.E.; Steele, A.M.; Saito, M.; Hacker, B.J.; Evers, B.M.; Saito, H. A New Cecal Slurry Preparation Protocol with Improved Long-Term Reproducibility for Animal Models of Sepsis. PLoS ONE 2014, 9, e115705. [Google Scholar] [CrossRef] [PubMed]
- Steele, A.M.; Starr, M.E.; Saito, H. Late Therapeutic Intervention with Antibiotics and Fluid Resuscitation Allows for a Prolonged Disease Course with High Survival in a Severe Murine Model of Sepsis. Shock 2017, 47, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Shrum, B.; Anantha, R.V.; Xu, S.X.; Donnelly, M.; Haeryfar, S.M.; McCormick, J.K.; Mele, T. A Robust Scoring System to Evaluate Sepsis Severity in an Animal Model. BMC Res. Notes 2014, 7, 233. [Google Scholar] [CrossRef]
- Laitano, O.; Van Steenbergen, D.; Mattingly, A.J.; Garcia, C.K.; Robinson, G.P.; Murray, K.O.; Clanton, T.L.; Nunamaker, E.A. Xiphoid Surface Temperature Predicts Mortality in a Murine Model of Septic Shock. Shock 2018, 50, 226–232. [Google Scholar] [CrossRef]
- Gordon, C.J.; Phillips, P.M.; Johnstone, A.F.M. A Noninvasive Method to Study Regulation of Extracellular Fluid Volume in Rats Using Nuclear Magnetic Resonance. Am. J. Physiol. Renal. Physiol. 2016, 310, F426–F431. [Google Scholar] [CrossRef]
- Morla, L.; Shore, O.; Lynch, I.J.; Merritt, M.E.; Wingo, C.S. A Noninvasive Method to Study the Evolution of Extracellular Fluid Volume in Mice Using Time-Domain Nuclear Magnetic Resonance. Am. J. Physiol. Renal. Physiol. 2020, 319, F115–F124. [Google Scholar] [CrossRef]
- Subramaniam, A.; McPhee, M.; Nagappan, R. Predicting Energy Expenditure in Sepsis: Harris-Benedict and Schofield Equations versus the Weir Derivation. Crit. Care Resusc. 2012, 14, 202–210. [Google Scholar]
- Pesta, D.; Gnaiger, E. High-Resolution Respirometry: OXPHOS Protocols for Human Cells and Permeabilized Fibers from Small Biopsies of Human Muscle. Methods Mol. Biol. 2012, 810, 25–58. [Google Scholar] [CrossRef]
- Doerrier, C.; Garcia-Souza, L.F.; Krumschnabel, G.; Wohlfarter, Y.; Mészáros, A.T.; Gnaiger, E. High-Resolution FluoRespirometry and OXPHOS Protocols for Human Cells, Permeabilized Fibers from Small Biopsies of Muscle, and Isolated Mitochondria. Methods Mol. Biol. 2018, 1782, 31–70. [Google Scholar] [CrossRef]
- Briguet, A.; Courdier-Fruh, I.; Foster, M.; Meier, T.; Magyar, J.P. Histological Parameters for the Quantitative Assessment of Muscular Dystrophy in the Mdx-Mouse. Neuromuscul. Disord. 2004, 14, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Mayeuf-Louchart, A.; Hardy, D.; Thorel, Q.; Roux, P.; Gueniot, L.; Briand, D.; Mazeraud, A.; Bouglé, A.; Shorte, S.L.; Staels, B.; et al. MuscleJ: A High-Content Analysis Method to Study Skeletal Muscle with a New Fiji Tool. Skeletal. Muscle 2018, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Osuchowski, M.F.; Ayala, A.; Bahrami, S.; Bauer, M.; Boros, M.; Cavaillon, J.-M.; Chaudry, I.H.; Coopersmith, C.M.; Deutschman, C.; Drechsler, S.; et al. Minimum Quality Threshold in Pre-Clinical Sepsis Studies (MQTiPSS): An International Expert Consensus Initiative for Improvement of Animal Modeling in Sepsis. Intensive Care Med. Exp. 2018, 6, 26. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Callahan, L.A.; Supinski, G.S. Sepsis-Induced Myopathy. Crit. Care Med. 2009, 37, S354–S367. [Google Scholar] [CrossRef]
- Kreymann, G.; Grosser, S.; Buggisch, P.; Gottschall, C.; Matthaei, S.; Greten, H. Oxygen Consumption and Resting Metabolic Rate in Sepsis, Sepsis Syndrome, and Septic Shock. Crit. Care Med. 1993, 21, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Sorriento, D.; Di Vaia, E.; Iaccarino, G. Physical Exercise: A Novel Tool to Protect Mitochondrial Health. Front. Physiol. 2021, 12, 660068. [Google Scholar] [CrossRef]
- Acheampong, A.; Vincent, J.-L. A Positive Fluid Balance Is an Independent Prognostic Factor in Patients with Sepsis. Crit. Care 2015, 19, 251. [Google Scholar] [CrossRef]
- Malbrain, M.L.N.G.; Van Regenmortel, N.; Saugel, B.; De Tavernier, B.; Van Gaal, P.-J.; Joannes-Boyau, O.; Teboul, J.-L.; Rice, T.W.; Mythen, M.; Monnet, X. Principles of Fluid Management and Stewardship in Septic Shock: It Is Time to Consider the Four D’s and the Four Phases of Fluid Therapy. Ann. Intensive Care 2018, 8, 66. [Google Scholar] [CrossRef]
- Chae, B.; Shin, Y.S.; Hong, S.-I.; Kim, S.M.; Kim, Y.-J.; Ryoo, S.M.; Kim, W.Y. Extracellular Water to Total Body Water Ratio in Septic Shock Patients Receiving Protocol-Driven Resuscitation Bundle Therapy. J. Clin. Med. 2021, 10, 2917. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, F.; Li, S.; Zheng, X.; Zheng, S.; Liu, H.; Lyu, J.; Yin, H. Influence of Fluid Balance on the Prognosis of Patients with Sepsis. BMC Anesthesiol. 2021, 21, 269. [Google Scholar] [CrossRef]
- Segar, J.L.; Balapattabi, K.; Reho, J.J.; Grobe, C.C.; Burnett, C.M.L.; Grobe, J.L. Quantification of Body Fluid Compartmentalization by Combined Time-Domain Nuclear Magnetic Resonance and Bioimpedance Spectroscopy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R44–R54. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, I.; Serra-Prat, M.; Yébenes, J.C. The Role of Water Homeostasis in Muscle Function and Frailty: A Review. Nutrients 2019, 11, 1857. [Google Scholar] [CrossRef] [PubMed]
- Hauptmann, S.; Klosterhalfen, B.; Weis, J.; Mittermayer, C.; Kirkpatrick, C.J. Skeletal Muscle Oedema and Muscle Fibre Necrosis during Septic Shock. Observations with a Porcine Septic Shock Model. Virchows Arch. 1994, 424, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, T.R.; Coakley, J.H.; Wagenmakers, A.J.M.; Griffiths, R.D.; Campbell, I.T.; Green, C.J.; McClelland, P.; Bone, J.M. Necrotizing Myopathy in Critically-Ill Patients. J. Pathol. 1991, 164, 307–314. [Google Scholar] [CrossRef]
- Jagoe, R.T.; Lecker, S.H.; Gomes, M.; Goldberg, A.L. Patterns of Gene Expression in Atrophying Skeletal Muscles: Response to Food Deprivation. FASEB J. 2002, 16, 1697–1712. [Google Scholar] [CrossRef]
- Chaillou, T.; Lee, J.D.; England, J.H.; Esser, K.A.; McCarthy, J.J. Time Course of Gene Expression during Mouse Skeletal Muscle Hypertrophy. J. Appl. Physiol. 2013, 115, 1065. [Google Scholar] [CrossRef]
- Calura, E.; Cagnin, S.; Raffaello, A.; Laveder, P.; Lanfranchi, G.; Romualdi, C. Meta-Analysis of Expression Signatures of Muscle Atrophy: Gene Interaction Networks in Early and Late Stages. BMC Genom. 2008, 9, 630. [Google Scholar] [CrossRef]
- Sandri, M.; Lin, J.; Handschin, C.; Yang, W.; Arany, Z.P.; Lecker, S.H.; Goldberg, A.L.; Spiegelman, B.M. PGC-1α Protects Skeletal Muscle from Atrophy by Suppressing FoxO3 Action and Atrophy-Specific Gene Transcription. Proc. Natl. Acad. Sci. USA 2006, 103, 16260–16265. [Google Scholar] [CrossRef]
- Guillon, A.; Preau, S.; Aboab, J.; Azabou, E.; Jung, B.; Silva, S.; Textoris, J.; Uhel, F.; Vodovar, D.; Zafrani, L.; et al. Preclinical Septic Shock Research: Why We Need an Animal ICU. Ann. Intensive Care 2019, 9, 66. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Qiao, S.; Li, H.; Che, T.; Wang, C.; An, J. Effects of Isoflurane on Complex II-associated Mitochondrial Respiration and Reactive Oxygen Species Production: Roles of Nitric Oxide and Mitochondrial KATP Channels. Mol. Med. Rep. 2019, 20, 4383–4390. [Google Scholar] [CrossRef]
- Suo, Z.; Yang, J.; Zhou, B.; Qu, Y.; Xu, W.; Li, M.; Xiao, T.; Zheng, H.; Ni, C. Whole-Transcriptome Sequencing Identifies Neuroinflammation, Metabolism and Blood-Brain Barrier Related Processes in the Hippocampus of Aged Mice during Perioperative Period. CNS Neurosci. Ther. 2022, 28, 1576–1595. [Google Scholar] [CrossRef] [PubMed]
- Bordon, Y. Protect the Species with Mitohormesis? Nat. Rev. Immunol. 2021, 21, 407. [Google Scholar] [CrossRef] [PubMed]
- van der Slikke, E.C.; Star, B.S.; van Meurs, M.; Henning, R.H.; Moser, J.; Bouma, H.R. Sepsis Is Associated with Mitochondrial DNA Damage and a Reduced Mitochondrial Mass in the Kidney of Patients with Sepsis-AKI. Crit. Care 2021, 25, 36. [Google Scholar] [CrossRef]
- Lancel, S.; Hassoun, S.M.; Favory, R.; Decoster, B.; Motterlini, R.; Neviere, R. Carbon Monoxide Rescues Mice from Lethal Sepsis by Supporting Mitochondrial Energetic Metabolism and Activating Mitochondrial Biogenesis. J. Pharmacol. Exp. Ther. 2009, 329, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, P.; Fovez, Q.; Germain, N.; Khamari, R.; Kluza, J. Mitochondrial Spare Respiratory Capacity: Mechanisms, Regulation, and Significance in Non-Transformed and Cancer Cells. FASEB J. 2020, 34, 13106–13124. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Guardia, D.; Palomer, X.; Coll, T.; Davidson, M.M.; Chan, T.O.; Feldman, A.M.; Laguna, J.C.; Vázquez-Carrera, M. The P65 Subunit of NF-KappaB Binds to PGC-1alpha, Linking Inflammation and Metabolic Disturbances in Cardiac Cells. Cardiovasc. Res. 2010, 87, 449–458. [Google Scholar] [CrossRef]
- Eisele, P.S.; Handschin, C. Functional Crosstalk of PGC-1 Coactivators and Inflammation in Skeletal Muscle Pathophysiology. Semin. Immunopathol. 2014, 36, 27–53. [Google Scholar] [CrossRef]
- Carré, J.E.; Orban, J.-C.; Re, L.; Felsmann, K.; Iffert, W.; Bauer, M.; Suliman, H.B.; Piantadosi, C.A.; Mayhew, T.M.; Breen, P.; et al. Survival in Critical Illness Is Associated with Early Activation of Mitochondrial Biogenesis. Am. J. Respir. Crit. Care Med. 2010, 182, 745–751. [Google Scholar] [CrossRef]
- Larche, J.; Lancel, S.; Hassoun, S.M.; Favory, R.; Decoster, B.; Marchetti, P.; Chopin, C.; Neviere, R. Inhibition of Mitochondrial Permeability Transition Prevents Sepsis-Induced Myocardial Dysfunction and Mortality. J. Am. Coll. Cardiol. 2006, 48, 377–385. [Google Scholar] [CrossRef]
- Boveris, A.; Alvarez, S.; Navarro, A. The Role of Mitochondrial Nitric Oxide Synthase in Inflammation and Septic Shock. Free. Radic. Biol. Med. 2002, 33, 1186–1193. [Google Scholar] [CrossRef]
- Clementi, E.; Brown, G.C.; Feelisch, M.; Moncada, S. Persistent Inhibition of Cell Respiration by Nitric Oxide: Crucial Role of S-Nitrosylation of Mitochondrial Complex I and Protective Action of Glutathione. Proc. Natl. Acad. Sci. USA 1998, 95, 7631–7636. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C.; Borutaite, V. Inhibition of Mitochondrial Respiratory Complex I by Nitric Oxide, Peroxynitrite and S-Nitrosothiols. Biochim. Biophys. Acta 2004, 1658, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Beltrán, B.; Orsi, A.; Clementi, E.; Moncada, S. Oxidative Stress and S-Nitrosylation of Proteins in Cells. Br. J. Pharmacol. 2000, 129, 953–960. [Google Scholar] [CrossRef]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy Pathway: Cellular and Molecular Mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Xin, H.; Mao, S.; Wu, G.; Guo, L. The Role of Autophagy in Sepsis: Protection and Injury to Organs. Front. Physiol. 2019, 10, 1071. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.; Palao, J.-C.; Castells, J.; Desgeorges, M.; Busso, T.; Molliex, S.; Jahnke, V.; Del Carmine, P.; Gondin, J.; Arnould, D.; et al. Regulation of Akt-MTOR, Ubiquitin-Proteasome and Autophagy-Lysosome Pathways in Locomotor and Respiratory Muscles during Experimental Sepsis in Mice. Sci. Rep. 2017, 7, 10866. [Google Scholar] [CrossRef]
- Masiero, E.; Agatea, L.; Mammucari, C.; Blaauw, B.; Loro, E.; Komatsu, M.; Metzger, D.; Reggiani, C.; Schiaffino, S.; Sandri, M. Autophagy Is Required to Maintain Muscle Mass. Cell Metab. 2009, 10, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Leduc-Gaudet, J.-P.; Miguez, K.; Cefis, M.; Moamer, A.; Chaffer, T.J.; Faitg, J.; Reynaud, O.; Broering, F.E.; Shams, A.; Mayaki, D.; et al. Role of Autophagy in Sepsis-Induced Skeletal Muscle Dysfunction, Whole-Body Metabolism, and Survival. bioRxiv 2021. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the Use and Interpretation of Assays for Monitoring Autophagy (4th Edition)1. Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef]
- Oh, C.-K.; Sultan, A.; Platzer, J.; Dolatabadi, N.; Soldner, F.; McClatchy, D.B.; Diedrich, J.K.; Yates, J.R.; Ambasudhan, R.; Nakamura, T.; et al. S-Nitrosylation of PINK1 Attenuates PINK1/Parkin-Dependent Mitophagy in HiPSC-Based Parkinson’s Disease Models. Cell Rep. 2017, 21, 2171–2182. [Google Scholar] [CrossRef] [PubMed]
- Westermann, B. Bioenergetic Role of Mitochondrial Fusion and Fission. Biochim. et Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 1833–1838. [Google Scholar] [CrossRef] [PubMed]
- Gomes, L.C.; Scorrano, L. High Levels of Fis1, a pro-Fission Mitochondrial Protein, Trigger Autophagy. Biochim. Biophys. Acta 2008, 1777, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Reignier, J.; Le Gouge, A.; Lascarrou, J.-B.; Annane, D.; Argaud, L.; Hourmant, Y.; Asfar, P.; Badie, J.; Nay, M.-A.; Botoc, N.-V.; et al. Impact of Early Low-Calorie Low-Protein versus Standard-Calorie Standard-Protein Feeding on Outcomes of Ventilated Adults with Shock: Design and Conduct of a Randomised, Controlled, Multicentre, Open-Label, Parallel-Group Trial (NUTRIREA-3). BMJ Open 2021, 11, e045041. [Google Scholar] [CrossRef]
- Marik, P.E.; Hooper, M.H. Normocaloric versus Hypocaloric Feeding on the Outcomes of ICU Patients: A Systematic Review and Meta-Analysis. Intensive Care Med. 2016, 42, 316–323. [Google Scholar] [CrossRef]
- Petros, S.; Horbach, M.; Seidel, F.; Weidhase, L. Hypocaloric vs Normocaloric Nutrition in Critically Ill Patients: A Prospective Randomized Pilot Trial. JPEN J. Parenter Enter. Nutr. 2016, 40, 242–249. [Google Scholar] [CrossRef]
- Brinkman, J.E.; Dorius, B.; Sharma, S. Physiology, Body Fluids. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Shin, J.; Park, I.; Lee, J.H.; Han, J.S.; Kim, B.; Jang, D.-H.; Lee, S.-M.; Lee, C.U.; Jo, Y.H. Comparison of Body Water Status and Its Distribution in Patients with Non-Septic Infection, Patients with Sepsis, and Healthy Controls. Clin. Exp. Emerg. Med. 2021, 8, 173–181. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Designation | Group Acronym | Intraperitoneal Injection | Feeding | Aim |
|---|---|---|---|---|
| Sham fed | SF | 10% glycerol–PBS | Free access | To control the energy balance |
| Sham pair-fed | SPF | 10% glycerol–PBS | Restricted | To reproduce the sepsis- like energy deficit |
| Sepsis | Sepsis | Heterologous stools | Free access | To assess the effect of sepsis on metabolic pathways |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pierre, A.; Bourel, C.; Favory, R.; Brassart, B.; Wallet, F.; Daussin, F.N.; Normandin, S.; Howsam, M.; Romien, R.; Lemaire, J.; et al. Sepsis-like Energy Deficit Is Not Sufficient to Induce Early Muscle Fiber Atrophy and Mitochondrial Dysfunction in a Murine Sepsis Model. Biology 2023, 12, 529. https://doi.org/10.3390/biology12040529
Pierre A, Bourel C, Favory R, Brassart B, Wallet F, Daussin FN, Normandin S, Howsam M, Romien R, Lemaire J, et al. Sepsis-like Energy Deficit Is Not Sufficient to Induce Early Muscle Fiber Atrophy and Mitochondrial Dysfunction in a Murine Sepsis Model. Biology. 2023; 12(4):529. https://doi.org/10.3390/biology12040529
Chicago/Turabian StylePierre, Alexandre, Claire Bourel, Raphael Favory, Benoit Brassart, Frederic Wallet, Frederic N. Daussin, Sylvain Normandin, Michael Howsam, Raphael Romien, Jeremy Lemaire, and et al. 2023. "Sepsis-like Energy Deficit Is Not Sufficient to Induce Early Muscle Fiber Atrophy and Mitochondrial Dysfunction in a Murine Sepsis Model" Biology 12, no. 4: 529. https://doi.org/10.3390/biology12040529
APA StylePierre, A., Bourel, C., Favory, R., Brassart, B., Wallet, F., Daussin, F. N., Normandin, S., Howsam, M., Romien, R., Lemaire, J., Grolaux, G., Durand, A., Frimat, M., Bastide, B., Amouyel, P., Boulanger, E., Preau, S., & Lancel, S. (2023). Sepsis-like Energy Deficit Is Not Sufficient to Induce Early Muscle Fiber Atrophy and Mitochondrial Dysfunction in a Murine Sepsis Model. Biology, 12(4), 529. https://doi.org/10.3390/biology12040529