




The Effects of Downhill Running and Maturation on Histological and Morphological Properties of Tendon and Enthesis in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Target Animals and Exercise Protocol
2.2. Sample Tissue Collection
2.3. Histological Evaluations of the Supraspinatus Muscles, Tendons, and Entheses
2.4. Immunohistochemical Evaluations of the Supraspinatus Tendon and Enthesis
2.5. Bone Morphological and Quality Evaluations of the Humerus Bone
2.6. Molecular Biological Evaluations of the Supraspinatus Tendon and Enthesis
2.7. Statistical Analysis
3. Results
3.1. Downhill Running Exercise Hypertrophies the Supraspinatus Muscle–Tendon Complex
3.2. Downhill Running Exercise Induces Histological Changes in the Enthesis Fibrocartilage Areas Depending on Maturity and Slight Changes in the Tissues Surrounding the Tendon
3.3. High Speed Downhill Running Exercise Induces Changes in Bone Morphology in Adolescents but Has Little Effect in Adults
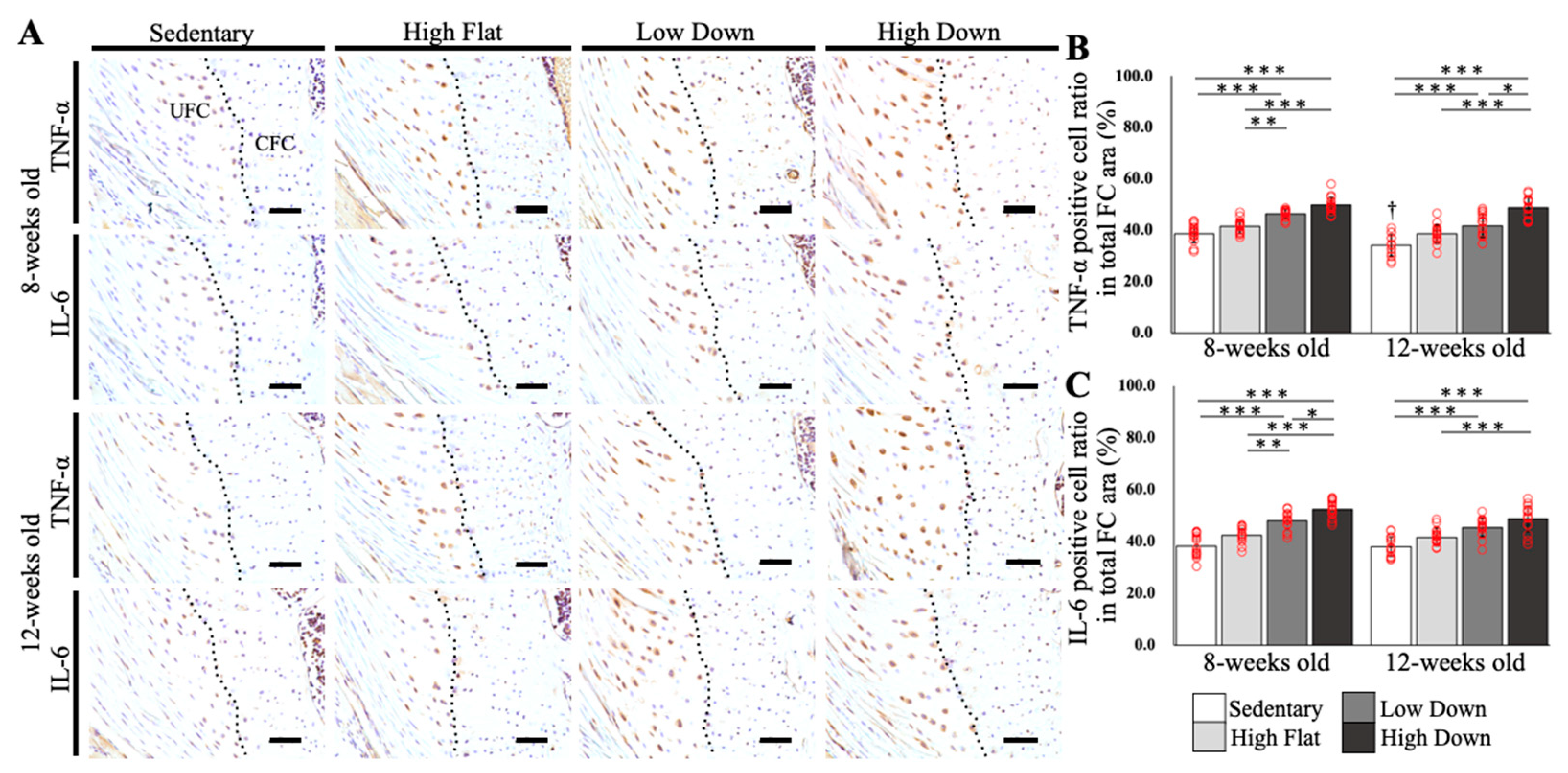
3.4. Downhill Running Exercise Induces Expression of Inflammation-Related Factors in Enthesis Fibrocartilage and Tendons
3.5. Inflammatory Factors Were Upregulated in the Fibrocartilages and Tendons in Young Mice and the Tendons in Adult Mice after Running Downhill
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Järvinen, A.H.; Kannus, P.; Maffulli, N.; Khan, K.M. Achilles Tendon Disorders: Etiology and Epidemiology. Foot Ankle Clin. 2005, 10, 255–266. [Google Scholar] [CrossRef]
- Blankstein, A.; Cohen, I.; Heim, M.; Diamant, L.; Salai, M.; Chechick, A.; Ganel, A. Ultrasonography as a diagnostic modality in Osgood-Schlatter disease. A clinical study and review of the literature. Arch. Orthop. Trauma Surg. 2001, 121, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Fujii, M.; Yoshimoto, M.; Abe, H.; Toda, N.; Higashiyama, R.; Takahira, N. Pathogenic factors associated with Osgood-Schlatter disease in adolescent male soccer players: A prospective cohort study. Orthop. J. Sport. Med. 2018, 6, 2325967118792192. [Google Scholar] [CrossRef] [PubMed]
- Draghi, F.; Danesino, G.M.; Coscia, D.; Precerutti, M.; Pagani, C. Overload syndromes of the knee in adolescents: Sonographic findings. J. Ultrasound 2008, 11, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.L.; Khan, K.M.; Kiss, Z.S.; Griffiths, L. Patellar tendinopathy in junior basketball players: A controlled clinical and ultrasonographic study of 268 patellar tendons in players aged 14–18 years. Scand. J. Med. Sci. Sport. 2000, 10, 216–220. [Google Scholar] [CrossRef]
- Magnusson, S.P.; Langberg, H.; Kjaer, M. The pathogenesis of tendinopathy: Balancing the response to loading. Nat. Rev. Rheumatol. 2010, 6, 262–268. [Google Scholar] [CrossRef]
- Adirim, T.A.; Cheng, T.L. Overview of Injuries in the Young Athlete. Sport. Med. 2003, 33, 75–81. [Google Scholar] [CrossRef]
- Millar, N.L.; Silbernagel, K.G.; Thorborg, K.; Kirwan, P.D.; Galatz, L.M.; Abrams, G.D.; Murrell, G.A.C.; McInnes, I.B.; Rodeo, S.A. Tendinopathy. Nat. Rev. Dis. Prim. 2021, 7, 1539–1554. [Google Scholar] [CrossRef]
- Khassetarash, A.; Vernillo, G.; Martinez, A.; Baggaley, M.; Giandolini, M.; Horvais, N.; Millet, G.Y.; Edwards, W.B. Biomechanics of graded running: Part II—Joint kinematics and kinetics. Scand. J. Med. Sci. Sport. 2020, 30, 1642–1654. [Google Scholar] [CrossRef]
- Roberts, T.J.; Konow, N. How tendons buffer energy dissipation by muscle. Exerc. Sport Sci. Rev. 2013, 41, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Ozone, K.; Kokubun, T.; Takahata, K.; Takahashi, H.; Yoneno, M.; Oka, Y.; Minegishi, Y.; Arakawa, K.; Kano, T.; Murata, K.; et al. Structural and pathological changes in the enthesis are influenced by the muscle contraction type during exercise. J. Orthop. Res. 2022, 40, 2076–2088. [Google Scholar] [CrossRef] [PubMed]
- Ozone, K.; Minegishi, Y.; Takahata, K.; Takahashi, H.; Yoneno, M.; Hattori, S.; Xianglan, L.; Oka, Y.; Murata, K.; Kanemura, N. Eccentric contraction-dominant exercise leads to molecular biological changes in enthesis and enthesopathy-like morphological changes. J. Orthop. Res. 2022, 41, 511–523. [Google Scholar] [CrossRef]
- Soslowsky, L.J.; Thomopoulos, S.; Tun, S.; Flanagan, C.L.; Keefer, C.C.; Mastaw, J.; Carpenter, J.E. Overuse activity injures the supraspinatus tendon in an animal model: A histologic and biomechanical study. J. Shoulder Elb. Surg. 2000, 9, 79–84. [Google Scholar] [CrossRef]
- Archambault, J.M.; Jelinsky, S.A.; Lake, S.P.; Hill, A.A.; Glaser, D.L.; Soslowsky, L.J. Rat supraspinatus tendon expresses cartilage markers with overuse. J. Orthop. Res. 2007, 25, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.B.; Ogilvie, R.W.; Schwane, J.A. Eccentric exercise-induced injury to rat skeletal muscle. J. Appl. Physiol. 1983, 54, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Portier, H.; Benaitreau, D.; Pallu, S. Does physical exercise always improve bone quality in rats? Life 2020, 10, 217. [Google Scholar] [CrossRef]
- Kanazawa, T.; Gotoh, M.; Ohta, K.; Shiba, N.; Nakamura, K.I. Novel characteristics of normal supraspinatus insertion in rats: An ultrastructural analysis using three-dimensional reconstruction using focused ion beam/scanning electron microscope tomography. Muscles. Ligaments Tendons J. 2014, 4, 182–187. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- JASP Team JASP; Version 0.16.3; JASP Team: Online, 2022.
- Love, J.; Selker, R.; Marsman, M.; Jamil, T.; Dropmann, D.; Verhagen, J.; Ly, A.; Gronau, Q.F.; Šmíra, M.; Epskamp, S.; et al. JASP: Graphical statistical software for common statistical designs. J. Stat. Softw. 2019, 88, 1–17. [Google Scholar] [CrossRef]
- Ghebes, C.A.; Groen, N.; Cheuk, Y.C.; Fu, S.C.; Fernandes, H.M.; Saris, D.B.F. Muscle-Secreted Factors Improve Anterior Cruciate Ligament Graft Healing: An In Vitro and In Vivo Analysis. Tissue Eng. Part A 2018, 24, 322–334. [Google Scholar] [CrossRef] [PubMed]
- West, D.W.D.; Lee-Barthel, A.; Mcintyre, T.; Shamim, B.; Lee, C.A.; Baar, K. The exercise-induced biochemical milieu enhances collagen content and tensile strength of engineered ligaments. J. Physiol. 2015, 593, 4665–4675. [Google Scholar] [CrossRef] [PubMed]
- Dubin, J.A.; Greenberg, D.R.; Iglinski-Benjamin, K.C.; Abrams, G.D. Effect of micro-RNA on tenocytes and tendon-related gene expression: A systematic review. J. Orthop. Res. 2018, 36, 2823–2829. [Google Scholar] [CrossRef] [PubMed]
- Pingel, J.; Wienecke, J.; Kongsgaard, M.; Behzad, H.; Abraham, T.; Langberg, H.; Scott, A. Increased mast cell numbers in a calcaneal tendon overuse model. Scand. J. Med. Sci. Sport. 2013, 23, e353–e360. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.V. Effects of grade and mass distribution on the mechanics of trotting in dogs. J. Exp. Biol. 2011, 214, 402–411. [Google Scholar] [CrossRef]
- Birn-Jeffery, A.V.; Higham, T.E. The scaling of uphill and downhill locomotion in legged animals. Integr. Comp. Biol. 2014, 54, 1159–1172. [Google Scholar] [CrossRef]
- Huang, C.Y.; Wang, V.M.; Pawluk, R.J.; Bucchieri, J.S.; Levine, W.N.; Bigliani, L.U.; Mow, V.C.; Flatow, E.L. Inhomogeneous mechanical behavior of the human supraspinatus tendon under uniaxial loading. J. Orthop. Res. 2005, 23, 924–930. [Google Scholar] [CrossRef]
- Tsumiyama, W.; Oki, S.; Takamiya, N.; Umei, N.; Shimizu, M.E.; Ono, T.; Otsuka, A. Induction of muscle hypertrophy in rats through low intensity eccentric contraction. J. Phys. Ther. Sci. 2014, 26, 1623–1625. [Google Scholar] [CrossRef]
- Hortobágyi, T. The positives of negatives: Clinical implications of eccentric resistance exercise in old adults. J. Gerontol.-Ser. A Biol. Sci. Med. Sci. 2003, 58, 417–418. [Google Scholar] [CrossRef]
- Flann, K.L.; Lastayo, P.C.; McClain, D.A.; Hazel, M.; Lindstedt, S.L. Muscle damage and muscle remodeling: No pain, no gain? J. Exp. Biol. 2011, 214, 674–679. [Google Scholar] [CrossRef]
- Franchi, M.V.; Reeves, N.D.; Narici, M. V Skeletal muscle remodeling in response to eccentric vs. concentric loading: Morphological, molecular, and metabolic adaptations. Front. Physiol. 2017, 8, 447. [Google Scholar] [CrossRef]
- Birch, H.L. Tendon matrix composition and turnover in relation to functional requirements. Int. J. Exp. Pathol. 2007, 88, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Heinemeier, K.M.; Schjerling, P.; Heinemeier, J.; Magnusson, S.P.; Kjaer, M. Lack of tissue renewal in human adult Achilles tendon is revealed by nuclear bomb 14 C. FASEB J. 2013, 27, 2074–2079. [Google Scholar] [CrossRef] [PubMed]
- Bohm, S.; Mersmann, F.; Arampatzis, A. Human tendon adaptation in response to mechanical loading: A systematic review and meta-analysis of exercise intervention studies on healthy adults. Sport. Med.-Open 2015, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, M.; Kumai, T.; Milz, S.; Boszczyk, B.M.; Boszczyk, A.; Ralphs, J. The skeletal attachment of tendons-tendon “entheses”. Comp. Biochem. Physiol. 2002, 133, 931–945. [Google Scholar] [CrossRef]
- Benjamin, M.; Ralphs, J.R. Fibrocartilage in tendons and ligaments—An adaptation to compressive load. J. Anat. 1998, 193, 481–494. [Google Scholar] [CrossRef]
- Vogel, K.G. Tendon structure and response to changing mechanical load. J. Musculoskelet. Neuronal Interact. 2003, 3, 323–325. [Google Scholar]
- Robbins, J.R.; Evanko, S.P.; Vogel, K.G. Mechanical loading and TGF-β regulate proteoglycan synthesis in tendon. Arch. Biochem. Biophys. 1997, 342, 203–211. [Google Scholar] [CrossRef]
- Evanko, S.P.; Vogel, K.G. Proteoglycan synthesis in fetal tendon is differentially regulated by cyclic compression in vitro. Arch. Biochem. Biophys. 1993, 307, 153–164. [Google Scholar] [CrossRef]
- Seki, N.; Itoi, E.; Shibuya, Y.; Wakabayashi, I.; Sano, H.; Sashi, R.; Minagawa, H.; Yamamoto, N.; Abe, H.; Kikuchi, K.; et al. Mechanical environment of the supraspinatus tendon: Three-dimensional finite element model analysis. J. Orthop. Sci. 2008, 13, 348–353. [Google Scholar] [CrossRef]
- Benjamin, M.; Toumi, H.; Ralphs, J.R.; Bydder, G.; Best, T.M.; Milz, S. Where tendons and ligaments meet bone: Attachment sites (‘entheses’) in relation to exercise and/or mechanical load. J. Anat. 2006, 208, 471–490. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Lories, R.J.; D’Agostino, M.A.; Elewaut, D.; Kirkham, B.; Soriano, E.R.; McGonagle, D. Enthesitis: From pathophysiology to treatment. Nat. Rev. Rheumatol. 2017, 13, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Kehl, A.S.; Corr, M.; Weisman, M.H. Review: Enthesitis: New insights into pathogenesis, diagnostic modalities, and treatment. Arthritis Rheumatol. 2016, 68, 312–322. [Google Scholar] [CrossRef]
- Watad, A.; Cuthbert, R.J.; Amital, H.; McGonagle, D. Enthesitis: Much more than focal insertion point inflammation. Curr. Rheumatol. Rep. 2018, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- El Jamal, A.; Briolay, A.; Mebarek, S.; Le Goff, B.; Blanchard, F.; Magne, D.; Brizuela, L.; Bougault, C. Cytokine-induced and stretch-induced sphingosine 1-phosphate production by enthesis cells could favor abnormal ossification in spondyloarthritis. J. Bone Miner. Res. 2019, 34, 2264–2276. [Google Scholar] [CrossRef]
- Boyer, M.I.; Hastings, H. Lateral tennis elbow: “Is there any science out there?” . J. Shoulder Elb. Surg. 1999, 8, 481–491. [Google Scholar] [CrossRef]
- Lipman, K.; Wang, C.; Ting, K.; Soo, C.; Zheng, Z. Tendinopathy: Injury, repair, and current exploration. Drug Des. Devel. Ther. 2018, 12, 591–603. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozone, K.; Minegishi, Y.; Oka, Y.; Sato, M.; Kanemura, N. The Effects of Downhill Running and Maturation on Histological and Morphological Properties of Tendon and Enthesis in Mice. Biology 2023, 12, 456. https://doi.org/10.3390/biology12030456
Ozone K, Minegishi Y, Oka Y, Sato M, Kanemura N. The Effects of Downhill Running and Maturation on Histological and Morphological Properties of Tendon and Enthesis in Mice. Biology. 2023; 12(3):456. https://doi.org/10.3390/biology12030456
Chicago/Turabian StyleOzone, Kaichi, Yuki Minegishi, Yuichiro Oka, Michiaki Sato, and Naohiko Kanemura. 2023. "The Effects of Downhill Running and Maturation on Histological and Morphological Properties of Tendon and Enthesis in Mice" Biology 12, no. 3: 456. https://doi.org/10.3390/biology12030456
APA StyleOzone, K., Minegishi, Y., Oka, Y., Sato, M., & Kanemura, N. (2023). The Effects of Downhill Running and Maturation on Histological and Morphological Properties of Tendon and Enthesis in Mice. Biology, 12(3), 456. https://doi.org/10.3390/biology12030456