Upregulation of Sarcolemmal Hemichannels and Inflammatory Transcripts with Neuromuscular Junction Instability during Lower Limb Unloading in Humans

 ,
,  ,
,  ,
,  ,
,  , ,
, ,  ,
,  ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. ULLS Model and Recruitment
2.2. Human Blood Sampling and Biopsies
2.3. ELISA
2.4. Immunostaining
2.5. Immunoblotting
2.6. SIMOA Analysis
2.7. RNA Extraction, Sequencing and Transcriptomic Analysis
2.8. Statistical Analysis and Graphics
2.9. Data Availability
3. Results
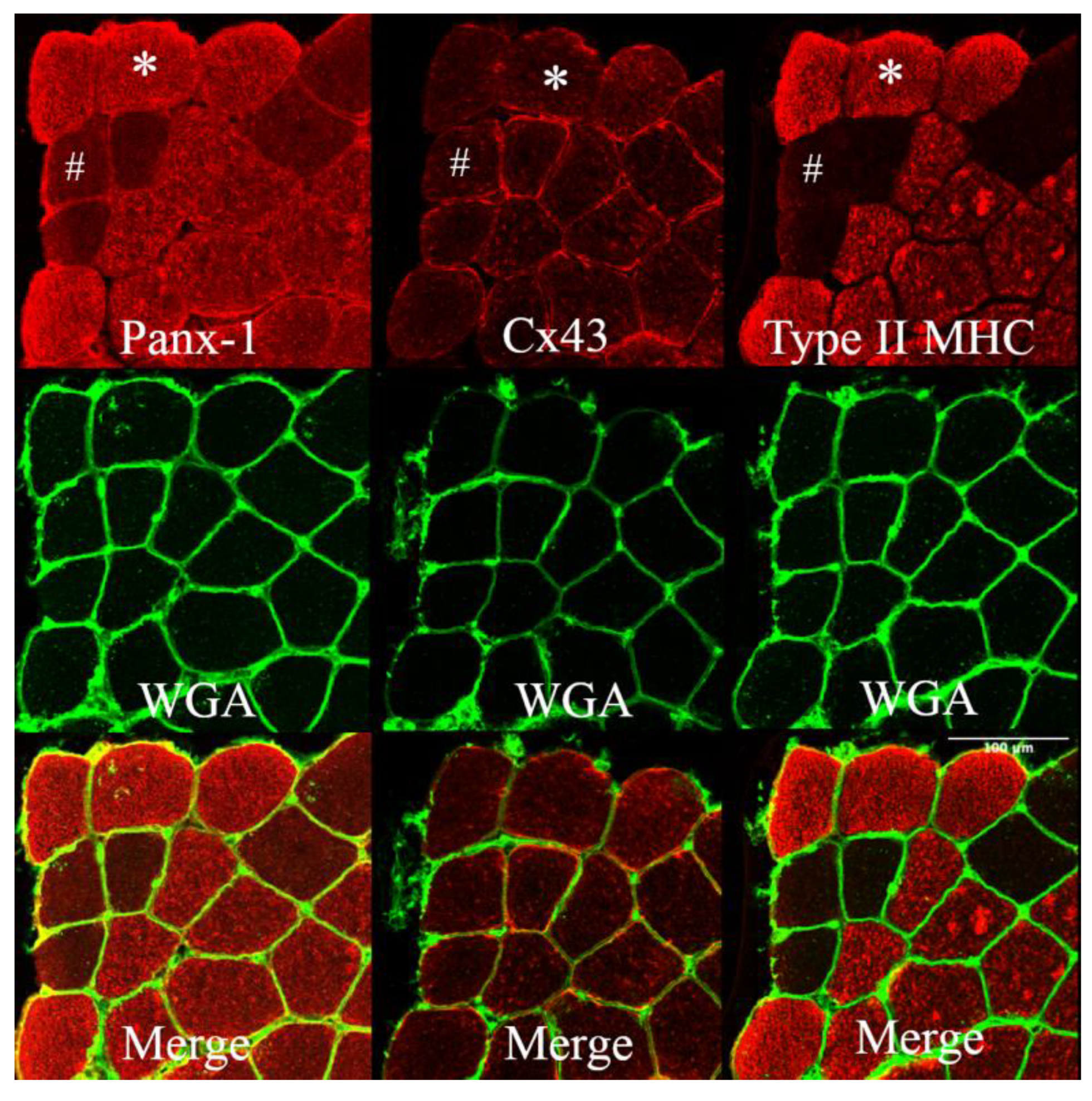
3.1. Connexin 43 and Pannexin-1 Expression in Human Vastus Lateralis
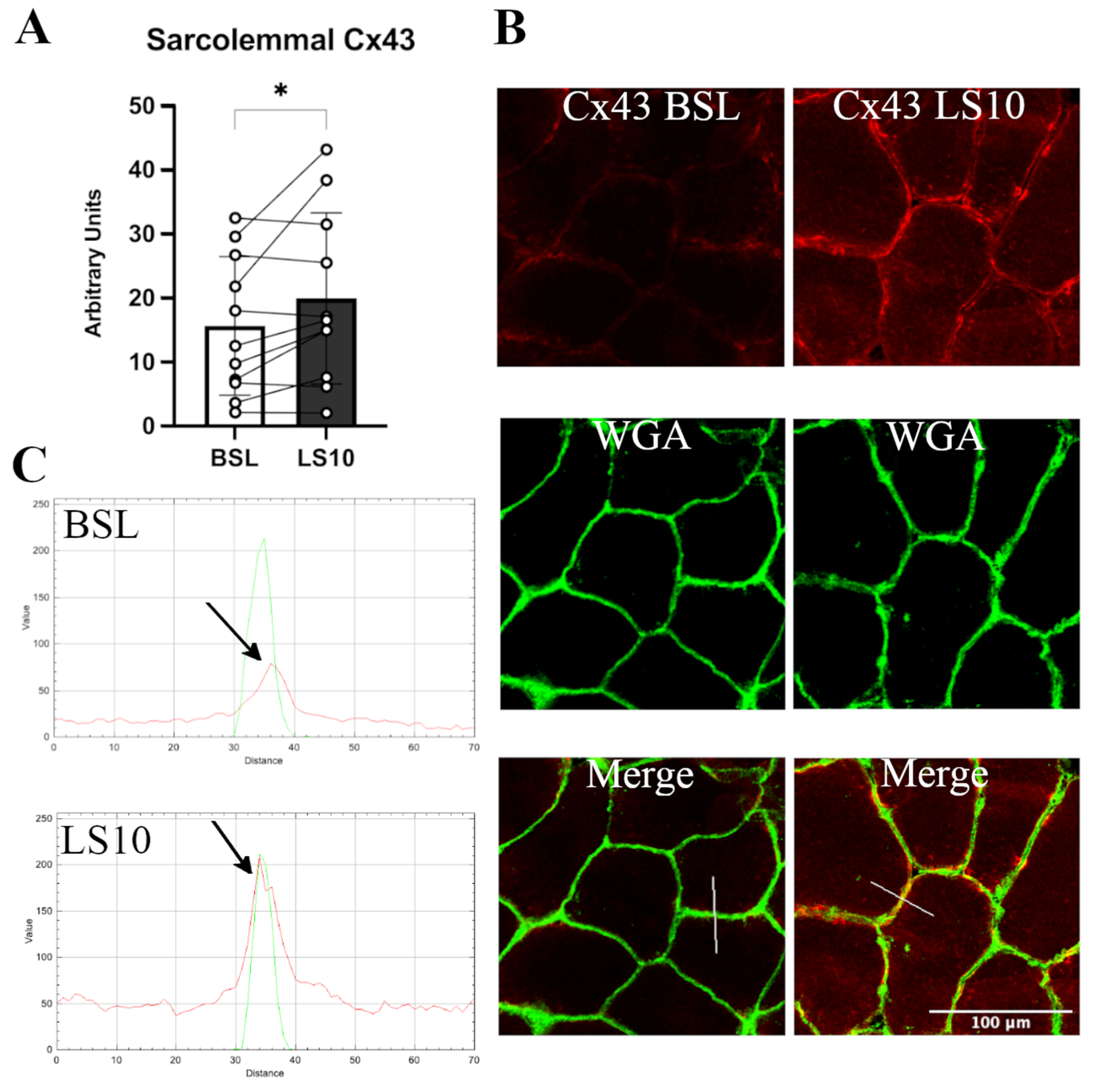
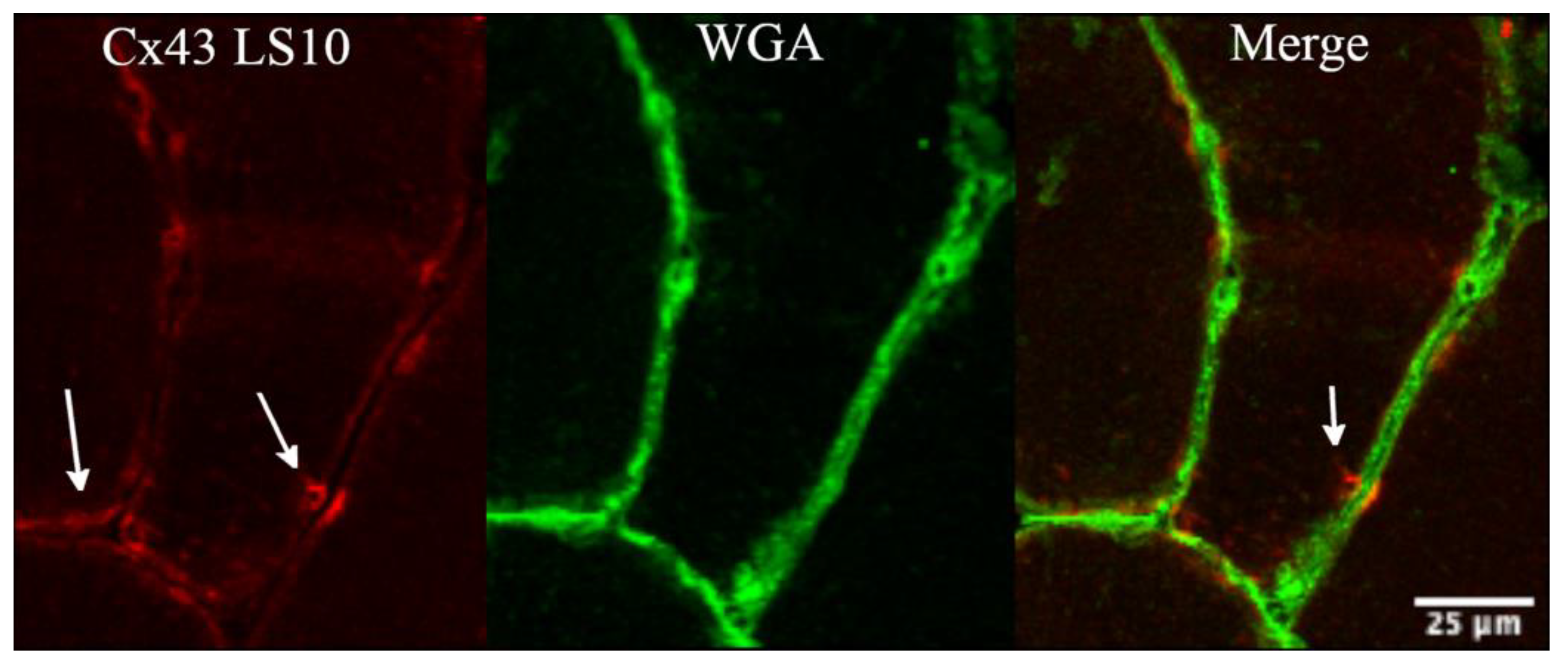
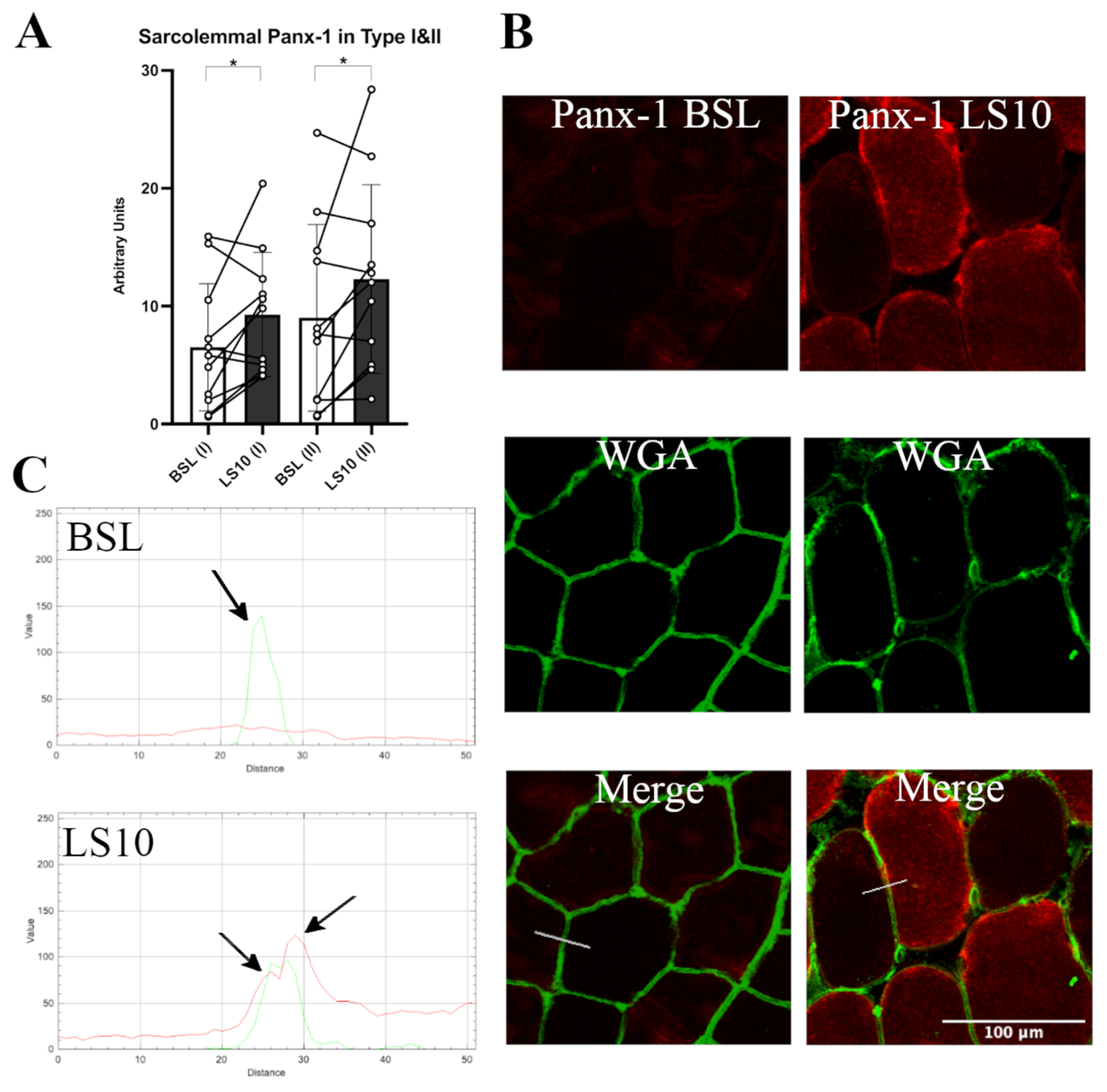
3.2. Sarcolemmal Upregulation of Connexin 43 and Pannexin-1
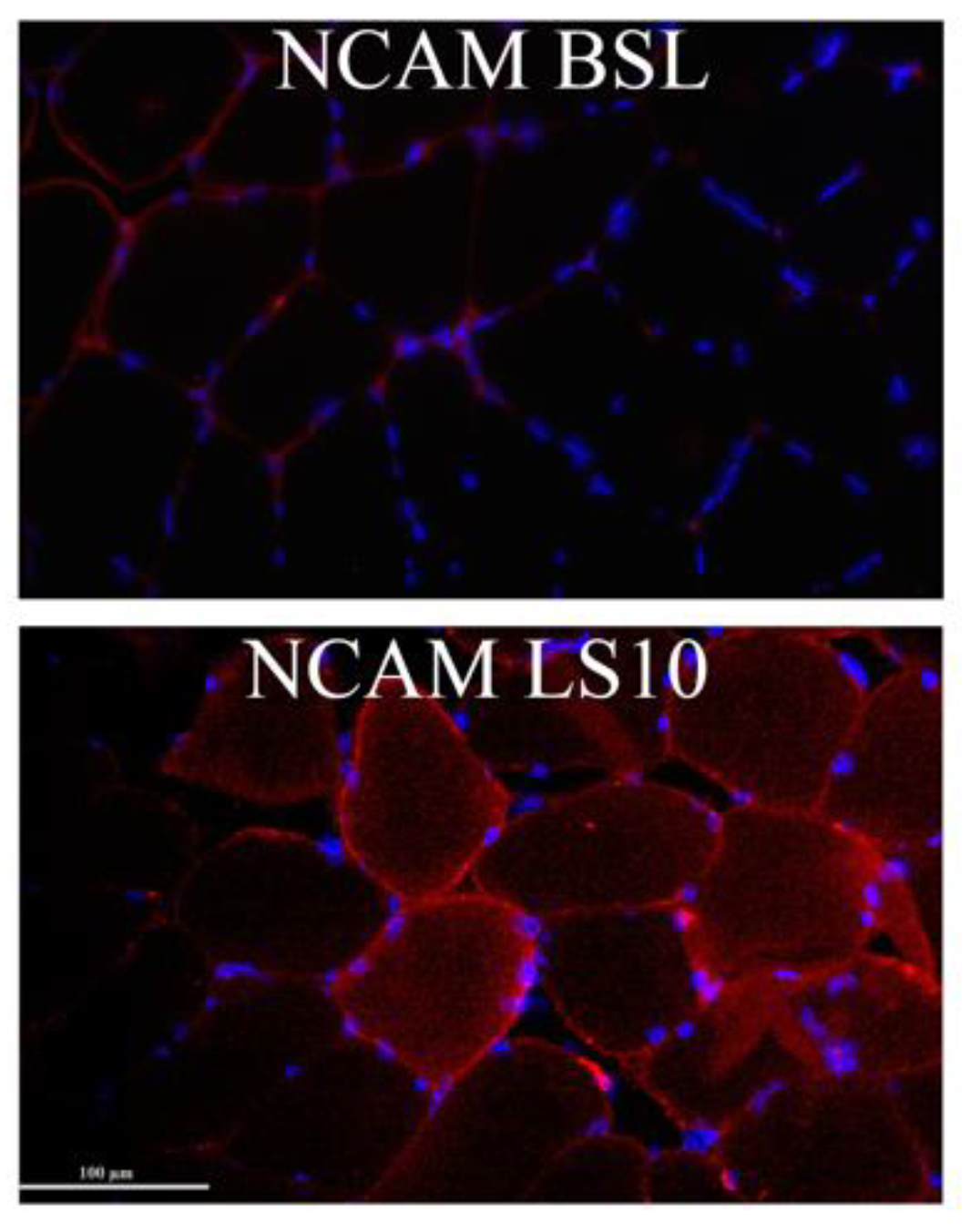
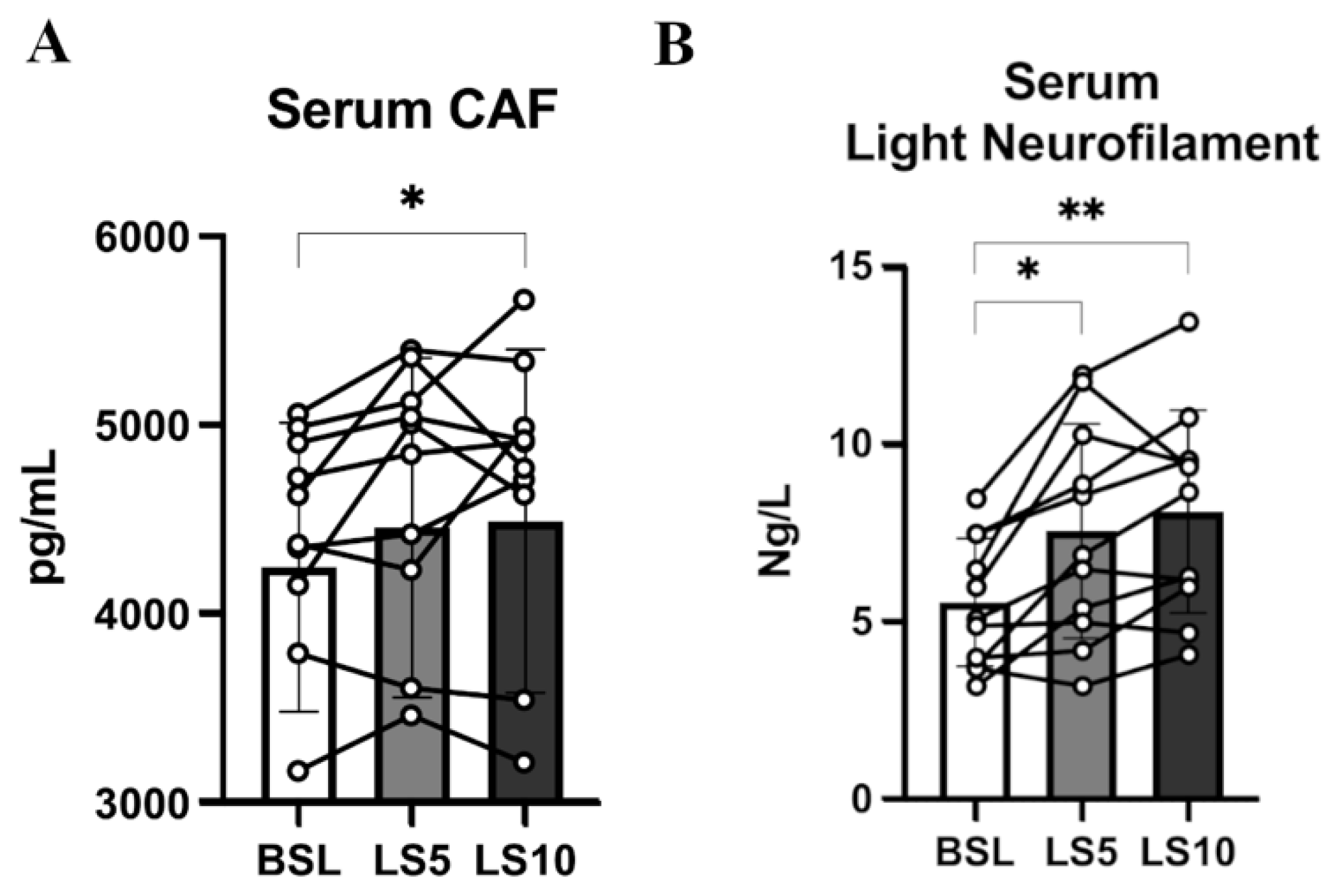
3.3. Muscle Fibre Denervation Events, Axonal Damage and Agrin Cleavage Processes
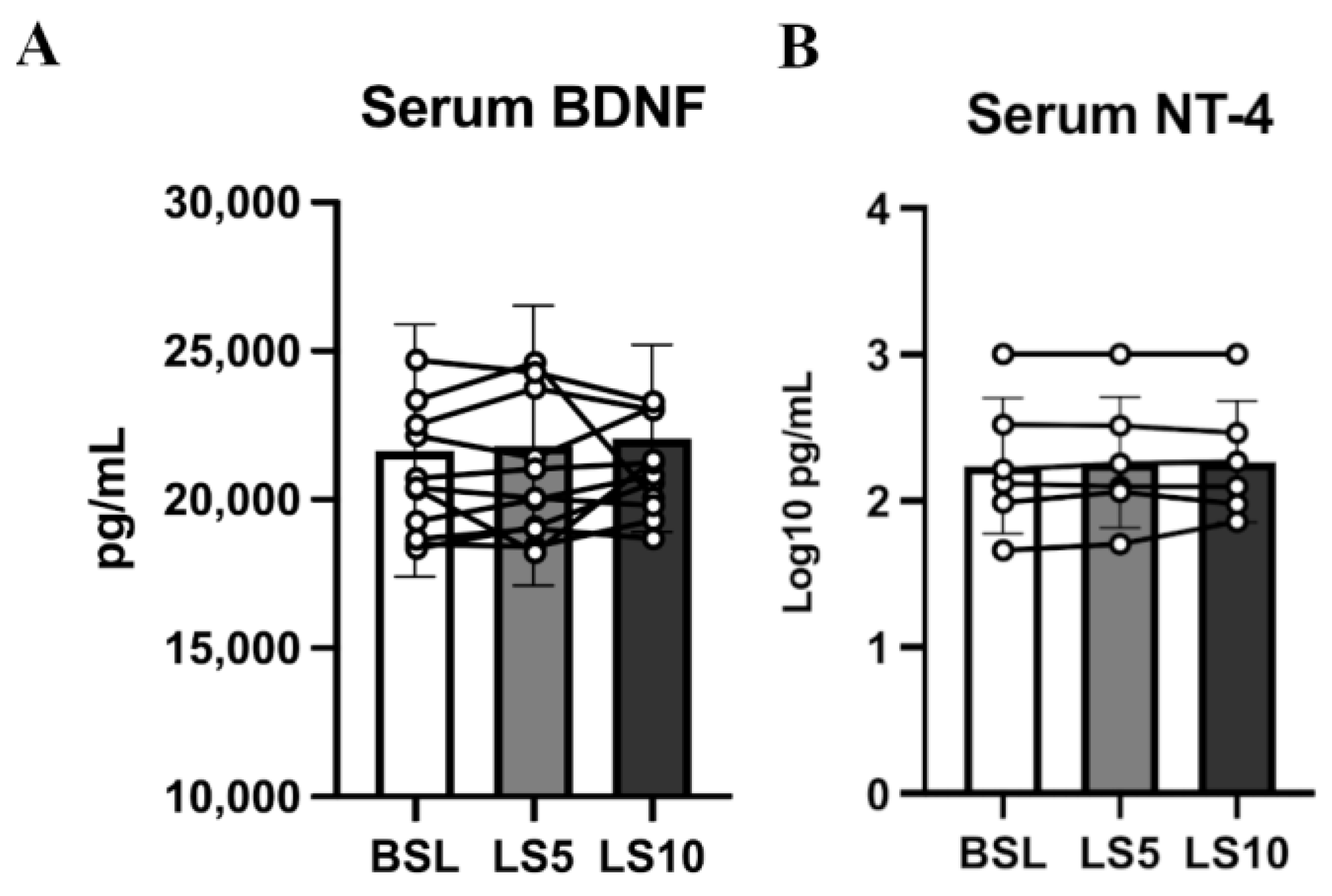
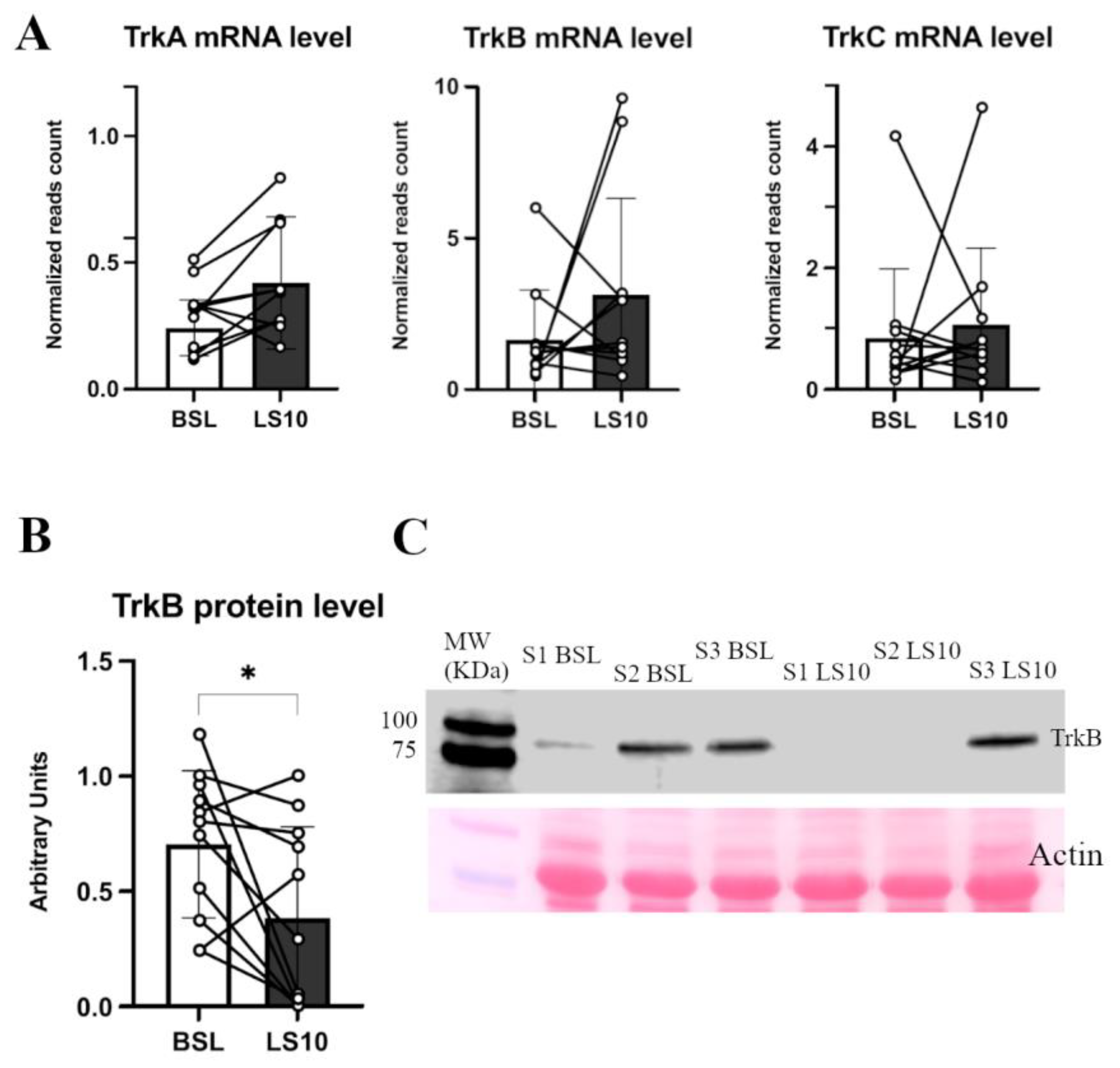
3.4. Circulating Neurotrophins and Neurotrophin Receptors
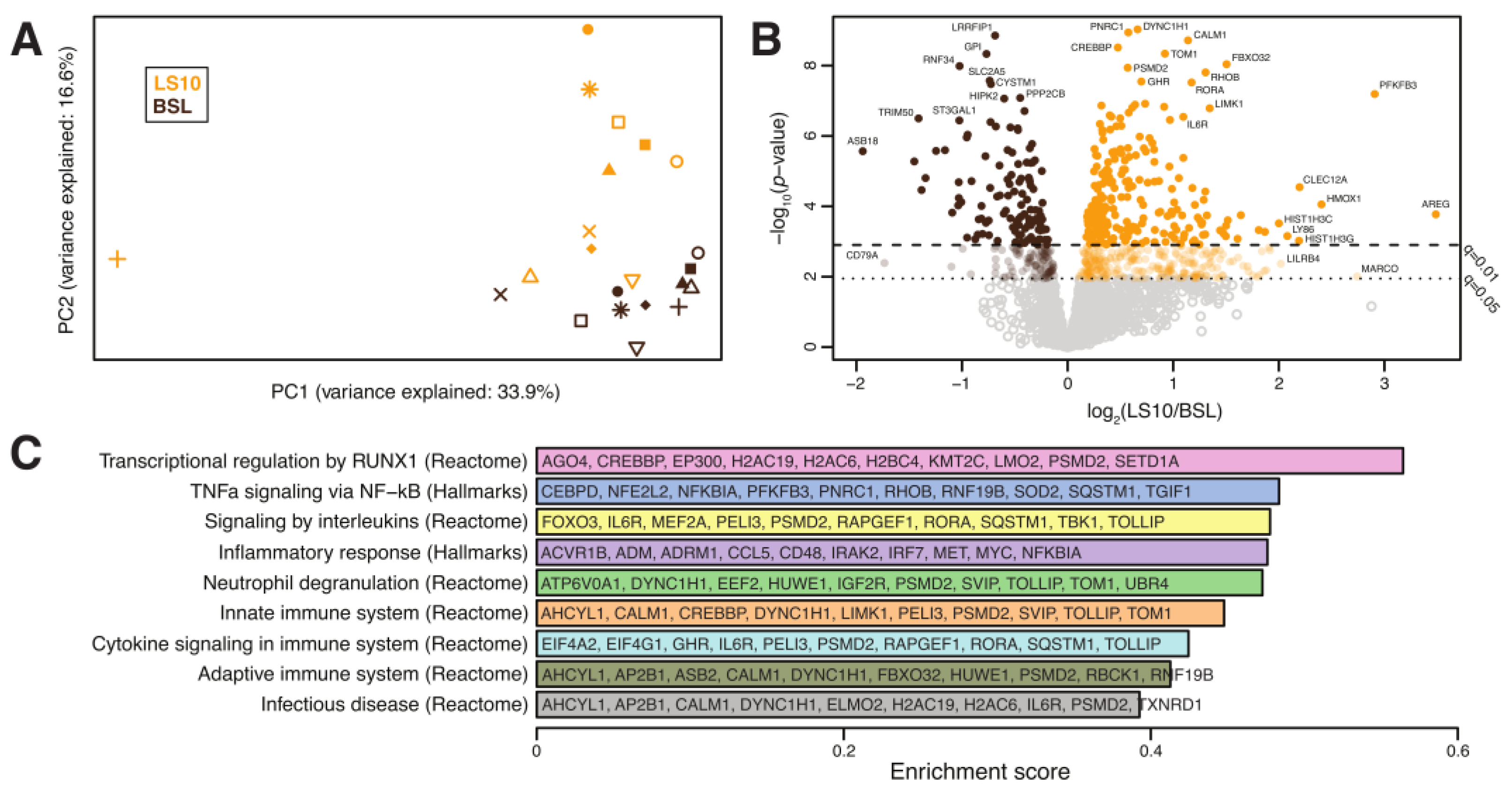
3.5. Transcriptomics Analysis of ULLS vs. Baseline Differences
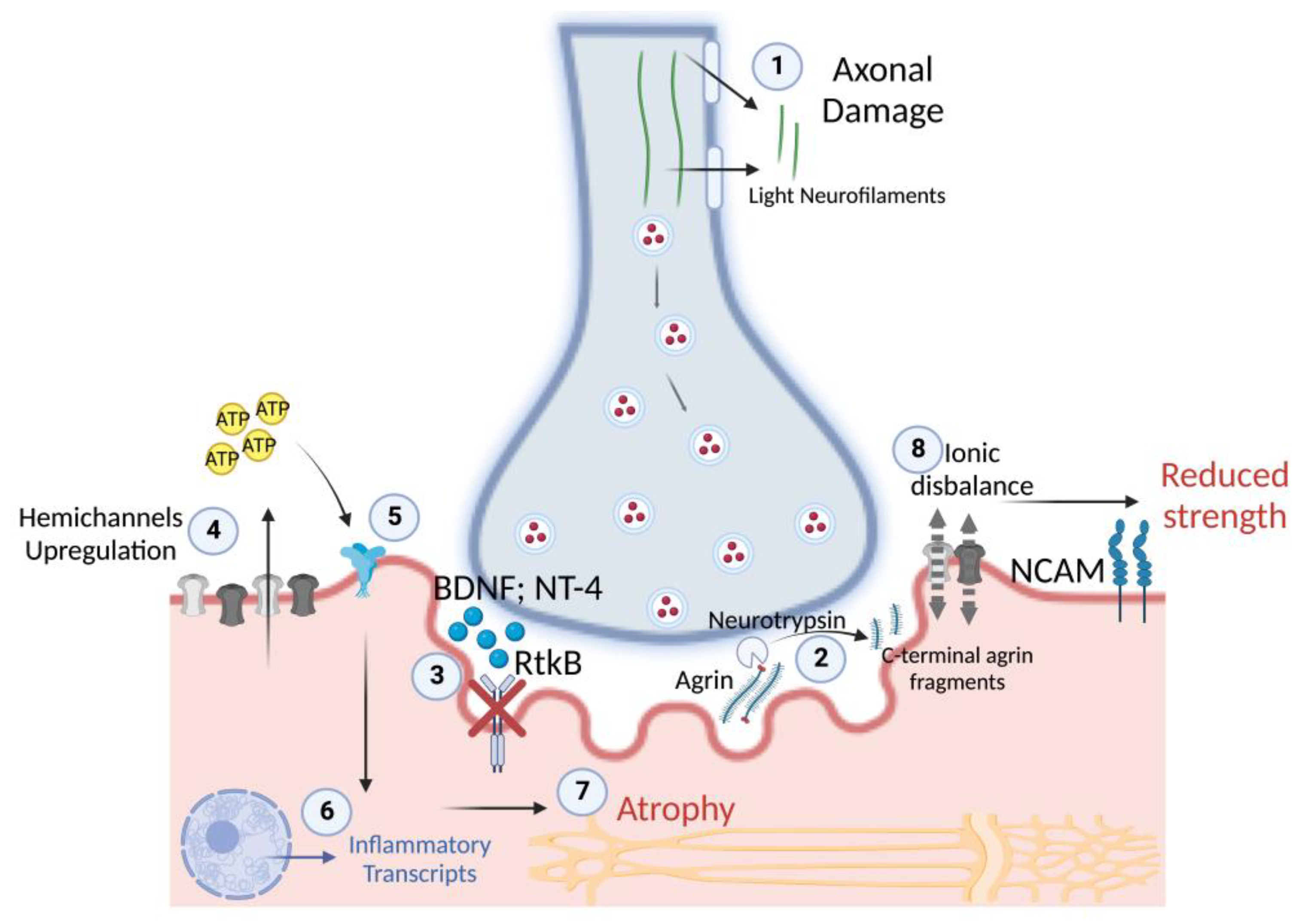
4. Discussion
4.1. Circulatory and Transcriptional Signatures of Muscle Denervation upon Disuse
4.2. Sarcolemmal Hemichannels Upregulation and Inflammatory Transcriptional Signatures Response upon Muscle Disuse
4.3. Neurotrophins’ Secretome, TrkB Receptor and Cxs Expression upon Muscle Disuse
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, P.H.U.; Chung, M.; Ren, Z.; Mair, D.B.; Kim, D.-H. Factors Mediating Spaceflight-Induced Skeletal Muscle Atrophy. Am. J. Physiol. Physiol. 2022, 322, C567–C580. [Google Scholar] [CrossRef]
- Franchi, M.V.; Sarto, F.; Simunič, B.; Pišot, R.; Narici, M.V. Early Changes of Hamstrings Morphology and Contractile Properties during 10 d of Complete Inactivity. Med. Sci. Sports Exerc. 2022, 54, 1346–1354. [Google Scholar] [CrossRef]
- Demangel, R.; Treffel, L.; Py, G.; Brioche, T.; Pagano, A.F.; Bareille, M.P.; Beck, A.; Pessemesse, L.; Candau, R.; Gharib, C.; et al. Early Structural and Functional Signature of 3-Day Human Skeletal Muscle Disuse Using the Dry Immersion Model. J. Physiol. 2017, 595, 4301–4315. [Google Scholar] [CrossRef]
- Monti, E.; Reggiani, C.; Franchi, M.V.; Toniolo, L.; Sandri, M.; Armani, A.; Zampieri, S.; Giacomello, E.; Sarto, F.; Sirago, G.; et al. Neuromuscular Junction Instability and Altered Intracellular Calcium Handling as Early Determinants of Force Loss during Unloading in Humans. J. Physiol. 2021, 599, 3037–3061. [Google Scholar] [CrossRef]
- Arentson-Lantz, E.J.; English, K.L.; Paddon-Jones, D.; Fry, C.S. Fourteen Days of Bed Rest Induces a Decline in Satellite Cell Content and Robust Atrophy of Skeletal Muscle Fibers in Middle-Aged Adults. J. Appl. Physiol. 2016, 120, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Sarto, F.; Stashuk, D.W.; Franchi, M.V.; Monti, E.; Zampieri, S.; Valli, G.; Sirago, G.; Candia, J.; Hartnell, L.M.; Paganini, M.; et al. Effects of Short-term Unloading and Active Recovery on Human Motor Unit Properties, Neuromuscular Junction Transmission and Transcriptomic Profile. J. Physiol. 2022, 600, 4731–4751. [Google Scholar] [CrossRef] [PubMed]
- Suetta, C.; Frandsen, U.; Jensen, L.; Jensen, M.M.; Jespersen, J.G.; Hvid, L.G.; Bayer, M.; Petersson, S.J.; Schrøder, H.D.; Andersen, J.L.; et al. Aging Affects the Transcriptional Regulation of Human Skeletal Muscle Disuse Atrophy. PLoS ONE 2012, 7, e51238. [Google Scholar] [CrossRef]
- Tesch, P.A.; Lundberg, T.R.; Fernandez-Gonzalo, R. Unilateral Lower Limb Suspension: From Subject Selection to “Omic” Responses. J. Appl. Physiol. 2016, 120, 1207–1214. [Google Scholar] [CrossRef]
- Cea, L.A.; Cisterna, B.A.; Puebla, C.; Frank, M.; Figueroa, X.F.; Cardozo, C.; Willecke, K.; Latorre, R.; Sáez, J.C. De Novo Expression of Connexin Hemichannels in Denervated Fast Skeletal Muscles Leads to Atrophy. Proc. Natl. Acad. Sci. USA 2013, 110, 16229–16234. [Google Scholar] [CrossRef]
- Finol, H.J.; Lewis, D.M.; Owens, R. The Effects of Denervation on Contractile Properties or Rat Skeletal Muscle. J. Physiol. 1981, 319, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Kotsias, B.A.; Venosa, R.A. Sodium Influx during Action Potential in Innervated and Denervated Rat Skeletal Muscles. Muscle Nerve 2001, 24, 1026–1033. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Davis, H.M.; Cisterna, B.A.; Sáez, J.C. Connexins and Pannexins in Bone and Skeletal Muscle. Curr. Osteoporos. Rep. 2017, 15, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Catoe, H.; Werner, R. MIR-206 Regulates Connexin43 Expression during Skeletal Muscle Development. Nucleic Acids Res. 2006, 34, 5863–5871. [Google Scholar] [CrossRef] [PubMed]
- Zaripova, K.A.; Kalashnikova, E.P.; Belova, S.P.; Kostrominova, T.Y.; Shenkman, B.S.; Nemirovskaya, T.L. Role of Pannexin 1 Atp-Permeable Channels in the Regulation of Signaling Pathways during Skeletal Muscle Unloading. Int. J. Mol. Sci. 2021, 22, 10444. [Google Scholar] [CrossRef]
- The Broad Institute The Picard Toolkit. Available online: https://broadinstitute.github.io/picard/ (accessed on 16 November 2021).
- Daley, T.; Smith, A.D. Predicting the Molecular Complexity of Sequencing Libraries. Nat. Methods 2013, 10, 325–327. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 16 November 2021).
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality Control of RNA-Seq Experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast Metagenomic Sequence Classification Using Exact Alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The Reference Human Genome Annotation for the ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, S.G. Voom: Precision Weights Unlock Linear Model Analysis Tools for RNA-Seq Read Counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1–25. [Google Scholar] [CrossRef]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.; Sergushichev, A. Fast Gene Set Enrichment Analysis. 2016; preprint. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Cisterna, B.A.; Vargas, A.A.; Puebla, C.; Fernández, P.; Escamilla, R.; Lagos, C.F.; Matus, M.F.; Vilos, C.; Cea, L.A.; Barnafi, E.; et al. Active Acetylcholine Receptors Prevent the Atrophy of Skeletal Muscles and Favor Reinnervation. Nat. Commun. 2020, 11, 1073. [Google Scholar] [CrossRef] [PubMed]
- Vanden Abeele, F.; Bidaux, G.; Gordienko, D.; Beck, B.; Panchin, Y.V.; Baranova, A.V.; Ivanov, D.V.; Skryma, R.; Prevarskaya, N. Functional Implications of Calcium Permeability of the Channel Formed by Pannexin 1. J. Cell Biol. 2006, 174, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Banzrai, C.; Nodera, H.; Kawarai, T.; Higashi, S.; Okada, R.; Mori, A.; Shimatani, Y.; Osaki, Y.; Kaji, R. Impaired Axonal Na+ Current by Hindlimb Unloading: Implication for Disuse Neuromuscular Atrophy. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.R.; O’Brien, W.T.; Sun, M.; Yamakawa, G.; O’Brien, T.J.; Mychasiuk, R.; Shultz, S.R.; McDonald, S.J.; Brady, R.D. Serum Neurofilament Light as a Biomarker of Traumatic Brain Injury in the Presence of Concomitant Peripheral Injury. Biomark. Insights 2021, 16. [Google Scholar] [CrossRef]
- Khalil, M.; Pirpamer, L.; Hofer, E.; Voortman, M.M.; Barro, C.; Leppert, D.; Benkert, P.; Ropele, S.; Enzinger, C.; Fazekas, F.; et al. Serum Neurofilament Light Levels in Normal Aging and Their Association with Morphologic Brain Changes. Nat. Commun. 2020, 11, 812. [Google Scholar] [CrossRef]
- Głowacka, A.; Ji, B.; Szczepankiewicz, A.A.; Skup, M.; Gajewska-Woźniak, O. BDNF Spinal Overexpression after Spinal Cord Injury Partially Protects Soleus Neuromuscular Junction from Disintegration, Increasing VAChT and AChE Transcripts in Soleus but Not Tibialis Anterior Motoneurons. Biomedicines 2022, 10, 2851. [Google Scholar] [CrossRef]
- Cisterna, B.A.; Cardozo, C.; Sáez, J.C. Neuronal Involvement in Muscular Atrophy. Front. Cell. Neurosci. 2014, 8, 405. [Google Scholar] [CrossRef]
- Skaper, S.D. The Neurotrophin Family of Neurotrophic Factors: An Overview. Methods Mol. Biol. 2012, 846, 1–12. [Google Scholar] [PubMed]
- Sakuma, K.; Watanabe, K.; Sano, M.; Uramoto, I.; Nakano, H.; Li, Y.J.; Kaneda, S.; Sorimachi, Y.; Yoshimoto, K.; Yasuhara, M.; et al. A Possible Role for BDNF, NT-4 and TrkB in the Spinal Cord and Muscle of Rat Subjected to Mechanical Overload, Bupivacaine Injection and Axotomy. Brain Res. 2001, 907, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Blagden, C.; Fan, J.; Nowak, S.J.; Taniuchi, I.; Littman, D.R.; Burden, S.J. Runx1 Prevents Wasting, Myofibrillar Disorganization, and Autophagy of Skeletal Muscle. Genes Dev. 2005, 19, 1715–1722. [Google Scholar] [CrossRef]
- Sirago, G.; Pellegrino, M.A.; Bottinelli, R.; Franchi, M.V.; Narici, M.V. Loss of Neuromuscular Junction Integrity and Muscle Atrophy in Skeletal Muscle Disuse. Ageing Res. Rev. 2023, 83, 101810. [Google Scholar] [CrossRef]
- Clark, J.A.; Southam, K.A.; Blizzard, C.A.; King, A.E.; Dickson, T.C. Axonal Degeneration, Distal Collateral Branching and Neuromuscular Junction Architecture Alterations Occur Prior to Symptom Onset in the SOD1G93A Mouse Model of Amyotrophic Lateral Sclerosis. J. Chem. Neuroanat. 2016, 76, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, P.A.; Laird, D.W. Connexins in Skeletal Muscle Development and Disease. Semin. Cell Dev. Biol. 2016, 50, 67–73. [Google Scholar] [CrossRef]
- Graham, Z.A. Mini-Review: Local and Downstream Responses to the Neuromuscular Junction: Potential Roles for Integrins, Connexins and Ephrins in Altering Muscle Characteristics and Function. Neurosci. Lett. 2022, 768, 136359. [Google Scholar] [CrossRef]
- Eggelbusch, M.; Shi, A.; Broeksma, B.C.; Vázquez-cruz, M.; Soares, M.N.; Wüst, R.C.I.; De Wit, G.M.J.; Everts, B.; Jaspers, R.T. The NLRP3 Inflammasome Contributes to Inflammation-Induced Morphological and Metabolic Alterations in Skeletal Muscle. J. Cachexia. Sarcopenia Muscle 2022, 13, 3048–3061. [Google Scholar] [CrossRef] [PubMed]
- Caiozzo, V.J.; Haddad, F.; Baker, M.J.; Herrick, R.E.; Prietto, N.; Baldwin, K.M. Microgravity-Induced Transformations of Myosin Isoforms and Contractile Properties of Skeletal Muscle. J. Appl. Physiol. 1996, 81, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.G.; Mckechnie, B.A.; Kelkar, S.; Fallon, J.R. Neurotrophins Regulate Agrin-Induced Postsynaptic Differentiation. Proc. Natl. Acad. Sci. USA 1999, 96, 1112–1117. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Code | Type | Dilution | Reaction Type |
|---|---|---|---|---|
| Anti-Connexin 43 | sc-271837 | Mouse Monoclonal | 1:50 | Unconjugated |
| Anti-Pannexin 1 | sc-515941 | Mouse Monoclonal | 1:30 | Unconjugated |
| Type II MHC | M 4276 Sigma | Mouse Monoclonal | 1:1000 | Unconjugated |
| Goat anti-Mouse | A-11031 Invitrogen | Alexa 568 | 1:300 | Alexa 568 |
| WGA | W11261 TF | Alexa 488 | 1:1000 | Alexa 488 |
| NCAM | AB5032 Chemicon | Rabbit Polyclonal | 1:200 | Unconjugated |
| Goat anti-Rabbit | A-11012 Invitrogen | Alexa 594 | 1:200 | Alexa 594 |
| Anti-TrkB | AB9872 Millipore | Rabbit Polyclonal | 1:5000 | Unconjugated |
| Goat anti-Rabbit | ab205718 Abcam | Goat Polyclonal | 1:10,000 | HRP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirago, G.; Candia, J.; Franchi, M.V.; Sarto, F.; Monti, E.; Toniolo, L.; Reggiani, C.; Giacomello, E.; Zampieri, S.; Hartnell, L.M.; et al. Upregulation of Sarcolemmal Hemichannels and Inflammatory Transcripts with Neuromuscular Junction Instability during Lower Limb Unloading in Humans. Biology 2023, 12, 431. https://doi.org/10.3390/biology12030431
Sirago G, Candia J, Franchi MV, Sarto F, Monti E, Toniolo L, Reggiani C, Giacomello E, Zampieri S, Hartnell LM, et al. Upregulation of Sarcolemmal Hemichannels and Inflammatory Transcripts with Neuromuscular Junction Instability during Lower Limb Unloading in Humans. Biology. 2023; 12(3):431. https://doi.org/10.3390/biology12030431
Chicago/Turabian StyleSirago, Giuseppe, Julián Candia, Martino V. Franchi, Fabio Sarto, Elena Monti, Luana Toniolo, Carlo Reggiani, Emiliana Giacomello, Sandra Zampieri, Lisa M. Hartnell, and et al. 2023. "Upregulation of Sarcolemmal Hemichannels and Inflammatory Transcripts with Neuromuscular Junction Instability during Lower Limb Unloading in Humans" Biology 12, no. 3: 431. https://doi.org/10.3390/biology12030431
APA StyleSirago, G., Candia, J., Franchi, M. V., Sarto, F., Monti, E., Toniolo, L., Reggiani, C., Giacomello, E., Zampieri, S., Hartnell, L. M., De Vito, G., Sandri, M., Ferrucci, L., & Narici, M. V. (2023). Upregulation of Sarcolemmal Hemichannels and Inflammatory Transcripts with Neuromuscular Junction Instability during Lower Limb Unloading in Humans. Biology, 12(3), 431. https://doi.org/10.3390/biology12030431