
Resource Partitioning among “Ancillary” Pelagic Fishes (Scomber spp., Trachurus spp.) in the Adriatic Sea

 ,
,  , , ,
, , ,  ,
, 
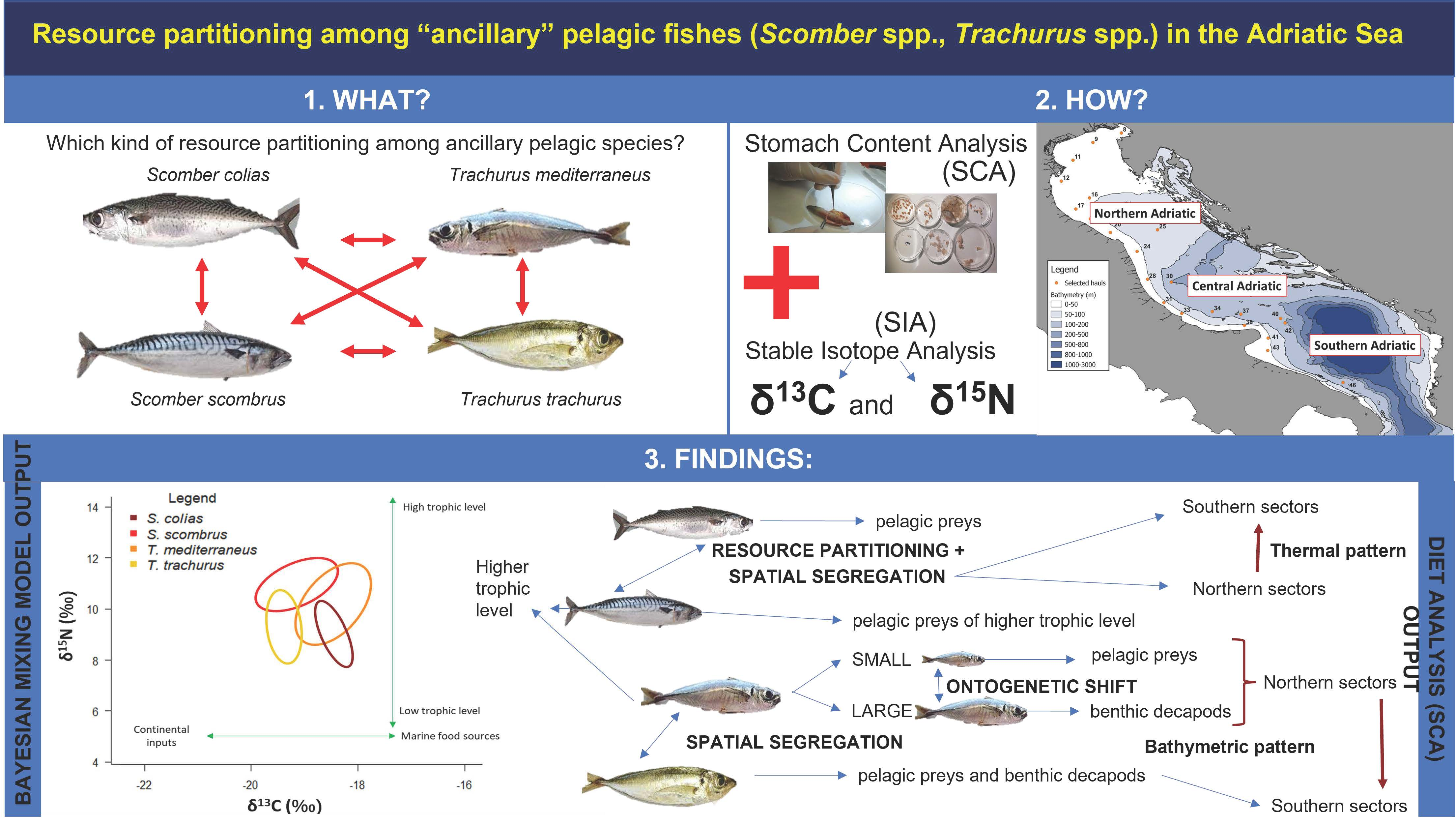
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Stomach Content Analysis
- (i)
- Percentage of frequency of occurrence (%F): where n is the number of guts containing a certain prey and N is the total number of guts examined.
- (ii)
- Percentage of gravimetric composition (%W): [37], where wi is the total weight of individuals of a certain prey i and Wp is the total weight of prey items.
- (iii)
- The index of relative importance (%IRI): %IRI = IRI/∑IRI × 100, where IRI = (%N + %W) × %F, as described in [38].
- (iv)
- Diet diversity within the year was calculated for each sex based on the Shannon–Wiener H′ index, calculated as H’ = −∑pi × lnpi, where pi is the ratio between the number of individuals (e.g., density) of each prey items (the ith species) and N is the total number of individuals of all prey species found in the stomach contents (pi = ni/N).
2.3. Stable Isotope Analysis
2.4. Data Treatment
3. Results
3.1. Sampling Data
3.2. Condition Indexes and Fullness of “Ancillary” Pelagic Fishes
3.3. Diets of “Ancillary” Pelagic Fishes
3.3.1. Scomber colias
3.3.2. Scomber scombrus
3.3.3. Trachurus mediterraneus
3.3.4. Trachurus trachurus
3.4. Overlap of the Diet of “Ancillary” Pelagic Fishes
3.5. Stable Isotope Composition of “Ancillary” Pelagic Fishes
3.6. Tropic Niches of “Ancillary” Pelagic Fishes
4. Discussion
4.1. Overall Feeding Ecology of the Pelagic “Ancillary” Species
4.1.1. Scomber colias
4.1.2. Scomber scombrus
4.1.3. Trachurus mediterraneus
4.1.4. Trachurus trachurus
4.2. Resource Partitioning of the Pelagic “Ancillary” Species
4.3. Role of Scomber spp. and Trachurus spp. in the Pelagic Food Web
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cushing, D.H.; Shipley, O.N.; Siskey, M.R. Pelagic fishes. In Encyclopedia of Ocean Sciences; Elsevier: Amsterdam, The Netherlands, 2019; Volume 2, pp. 290–296. [Google Scholar]
- Fanelli, E.; Da Ros, Z.; Menicucci, S.; Malavolti, S.; Biagiotti, I.; Canduci, G.; De Felice, A.; Leonori, I. The pelagic food web of the Adriatic Sea: A focus on the role of small pelagics. Front. Mar. Sci. Under Review.
- Bas, C. Ecological structures: Expansion and replacement. Sci. Mar. 1995, 59, 373–380. [Google Scholar]
- Rice, J. Food web theory, marine food webs, and what climate change may do to northern marine fish populations: Climate change and northern fish populations. Can. Spec. Publ. Fish. Aquat. Sci. 1995, 121, 561–568. [Google Scholar]
- Fréon, P.; Cury, P.; Shannon, L.; Roy, C. Sustainable exploitation of small pelagic fish stocks challenged by environmental and ecosystem changes: A review. Bullettin Mar. Sci. 2005, 76, 385–462. [Google Scholar]
- Santojanni, A.; Cingolani, N.; Arneri, E.; Kirkwood, G.; Belardinelli, A.; Giannetti, G.; Colella, S.; Donato, F.; Barry, C. Stock assessment of sardine (Sardina pilchardus, Walb.) in the Adriatic Sea with an estimate of discards. Sci. Mar. 2005, 69, 603–617. [Google Scholar] [CrossRef]
- FAO Fisheries and Aquaculture Department. Review of the State of World Marine Fishery Resources; FAO: Rome, Italy, 2011. [Google Scholar]
- FAO. The State of Mediterranean and Black Sea Fisheries 2020; General Fisheries Commission for the Mediterranean: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- IREPA (Istituto Ricerche Economia Pesca ed Acquicoltura). Osservatorio Economico sulle Strutture Produttive della Pesca Marittima in Italia; Edizioni Scientifiche Italiane: Napoli, Italy, 2011. [Google Scholar]
- Cikeš Keč, V.; Zorica, B. The reproductive traits of Scomber japonicus (Houttuyn, 1782) in the Eastern Adriatic Sea. J. Appl. Ichthyol. 2012, 28, 15–21. [Google Scholar] [CrossRef]
- Meneghesso, C.; Riginella, E.; La Mesa, M.; Donato, F.; Mazzoldi, C. Life-history traits and population decline of the Atlantic mackerel Scomber scombrus in the Adriatic Sea. J. Fish Biol. 2013, 83, 1249–1267. [Google Scholar] [CrossRef]
- Azzurro, E.; Moschella, P.; Maynou, F. Tracking signals of change in Mediterranean fish diversity based on local ecological knowledge. PLoS ONE 2011, 6, e24885. [Google Scholar] [CrossRef]
- Barausse, A.; Michieli, A.; Riginella, E.; Palmeri, L.; Mazzoldi, C. Long-term changes in community composition and life-history traits in a highly exploited basin (northern Adriatic Sea): The role of environment and anthropogenic pressures. J. Fish Biol. 2011, 79, 1453–1486. [Google Scholar] [CrossRef]
- Zardoya, R.; Castilho, R.; Grande, C.; Favre-Krey, L.; Caetano, S.; Marcato, S.; Krey, G.; Patarnello, T. Differential population structuring of two closely related fish species, the mackerel (Scomber scombrus) and the chub mackerel (Scomber japonicus), in the Mediterranean Sea. Mol. Ecol. 2004, 13, 1785–1798. [Google Scholar] [CrossRef]
- Šantić, M.; Jardas, I.; Pallaoro, A. Feeding Habits of Mediterranean Horse Mackerel, Trachurus mediterraneus (Carangidae), in the central Adriatic Sea. Cybium 2003, 27, 247–253. [Google Scholar]
- Collette, B.B.; Nauen, C.E. FAO Species Catalogue. In FAO Fisheries Synopsis 125; FAO: Rome, Italy, 1983; Volume 2. [Google Scholar]
- Smith-Vaniz, W.F. Carangidae. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volume 2, pp. 815–844. [Google Scholar]
- Nielsen, J.M.; Clare, E.L.; Hayden, B.; Brett, M.T.; Kratina, P.; Gilbert, M.T.P. Diet tracing in ecology: Method comparison and selection. Methods Ecol. Evol. 2017, 9, 278–291. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology 1999, 80, 1395–1404. [Google Scholar] [CrossRef]
- Post, D. Using stable isotopes to estimate trophic position: Models, methods and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Pinnegar, J.K.; Polunin, N.V.C. Contributions of stable-isotope data to elucidating food webs of Mediterranean rocky littoral fishes. Oecologia 2000, 122, 399–409. [Google Scholar] [CrossRef]
- France, R.L. Carbon-13 enrichment in benthic compared to planktonic algae: Foodweb implications. Mar. Ecol. Prog. Ser. 1995, 124, 307–312. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Et Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Fanelli, E.; Badalamenti, F.; D’Anna, G.; Pipitone, C.; Riginella, E.; Azzurro, E. Food partitioning and diet temporal variation in two coexisting sparids, Pagellus erythrinus and Pagellus acarne. J. Fish Biol. 2011, 78, 869–900. [Google Scholar] [CrossRef]
- Rumolo, P.; Fanelli, E.; Barra, M.; Basilone, G.; Genovese, S.; Gherardi, S.; Ferreri, R.; Gargano, A.; Mazzola, S.; Bonanno, A. Trophic relationships between anchovy (Engraulis encrasicolus) and zooplankton in the Strait of Sicily (Central Mediterranean sea): A stable isotope approach. Hydrobiologia 2018, 821, 41–56. [Google Scholar] [CrossRef]
- Leonori, I.; Ticina, V.; De Felice, A.; Vidjak, O.; Grubisic, L.; Pallaoro, A. Comparisons of two research vessels’ properties in the acoustic surveys of small pelagic fish. Acta Adriat. 2012, 53, 389–398. [Google Scholar]
- Leonori, I.; Tičina, V.; Giannoulaki, M.; Hattab, T.; Iglesias, M.; Bonanno, A.; Costantini, I.; Canduci, G.; Machias, A.; Ventero, A.; et al. History of hydroacoustic surveys of small pelagic fish species in the European Mediterranean Sea. Mediterr. Mar. Sci. 2021, 22, 751–768. [Google Scholar] [CrossRef]
- Simmonds, E.J.; Maclennan, D.N. Fisheries Acoustics: Theory and Practice; Blackwell Publishing: Oxford, UK, 2005; p. 456. [Google Scholar]
- MEDIAS. Common Protocol for the Pan-MEDiterranean Acoustic Survey. MEDIAS Handbook. Available online: http://www.medias-project.eu/medias/website (accessed on 6 October 2022).
- Walsh, M.; Hopkins, P.; Witthames, P.R.; Greer-Walker, M.; Watson, J. Estimation of the Potential Fecundity and Atresia in the Western Mackerel Stock 1989; ICES (International Council for the Exploration of the Sea) Conference and Meeting Document: 1990/H:31; ICES: Copenhagen, Denmark, 1990. [Google Scholar]
- ICES. Report of the Workshop on Maturity Staging of Mackerel and Horse Mackerel (WKMSMAC2); 28 September–2 October 2015; ICES: Lisbon, Portugal, 2015. [Google Scholar]
- Fanelli, E.; Rumolo, P.; Barra, M.; Basilone, G.; Genovese, S.; Bonanno, A. Mesoscale variability in the trophic ecology of the European hake Merluccius merluccius in the Strait of Sicily. Hydrobiologia 2018, 821, 57–72. [Google Scholar] [CrossRef]
- Rumolo, P.; Basilone, G.; Fanelli, E.; Barra, M.; Calabrò, M.; Genovese, S.; Gherardi, S.; Ferreri, R.; Mazzola, S.; Bonanno, A. Linking spatial distribution and feeding behavior of Atlantic horse mackerel (Trachurus trachurus) in the Strait of Sicily (Central Mediterranean Sea). J. Sea Res. 2017, 121, 47–58. [Google Scholar] [CrossRef]
- Olaso, I.; Sànchez, F.; Rodrìguez-Cabello, C.; Velasco, F. The feeding behaviour of some demersal fish species in response to artificial discarding. Sci. Mar. 2002, 66, 301–311. [Google Scholar] [CrossRef]
- Hyslop, E. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Pinkas, L.; Oliphant, M.S.; Iverson, I.L.K. Food Habits of Albacore, Bluefin Tuna, and Bonito In California Waters. Fish Bull. 1971, 152, 1–139. [Google Scholar]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Logan, M.J.; Jardine, T.D.; Miller, T.J.; Bunn, S.E.; Cunjak, R.A.; Lutcavage, M.E. Lipid Corrections in Carbon and Nitrogen Stable Isotope Analyses: Comparison of Chemical Extraction and Modelling Methods. J. Anim. Ecol. 2008, 77, 838–846. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 20 December 2022).
- Clarke, K.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Layman, C.A.; Quattrochi, J.P.; Peyer, C.M.; Allgeier, J.E. Niche width collapse in a resilient top predator following ecosystem fragmentation. Ecol. Lett. 2007, 10, 937–944. [Google Scholar] [CrossRef]
- Sever, T.M.; Bayhan, B.; Bilecenoglu, M.; Mavili, S. Diet composition of the juvenile chub mackerel (Scomber japonicus) in the Aegean Sea (Izmir Bay, Turkey). J. Appl. Ichthyol. 2006, 22, 145–148. [Google Scholar] [CrossRef]
- Castro, J. Mysids and euphausiids in the diet of Scomber japonicus Houttuyn, 1782 off the Canary Islands. Boletín-Inst. Español De Oceanogr. 1995, 11, 77–86. [Google Scholar]
- Gushchin, A.V.; Corten, A. Feeding of pelagic fish in waters of Mauritania: Atlantic Chub mackerel Scomber colias, Atlantic horse mackerel Trachurus trachurus, Cunene horse mackerel Trachurus trecae. J. Ichthyol. 2017, 57, 410–423. [Google Scholar] [CrossRef]
- Ménard, F.; Dallot, S.; Thomas, S.; Braconnot, J.C. Temporal fluctuations of two Mediterranean salp populations from 1967 to 1990. Analysis of the influence of environmental variables using a Markov chain model. Mar. Ecol. Prog. Ser. 1994, 104, 139–152. [Google Scholar] [CrossRef]
- Ursella, L.; Cardin, V.; Batistić, M.; Garić, R.; Gačić, M. Evidence of zooplankton vertical migration from continuous Southern Adriatic buoy current-meter records. Prog. Oceanogr. 2018, 167, 78–96. [Google Scholar] [CrossRef]
- Fanelli, E.; Cartes, J.E.; Papiol, V. Food web structure of deep-sea macrozooplankton and micronekton off the Catalan slope: Insight from stable isotopes. J. Mar. Syst. 2011, 87, 79–89. [Google Scholar] [CrossRef]
- Fanelli, E.; Cartes, J.E.; Rumolo, P.; Sprovieri, M. Food-web structure and trophodynamics of mesopelagic–suprabenthic bathyal macrofauna of the Algerian Basin based on stable isotopes of carbon and nitrogen. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 1504–1520. [Google Scholar] [CrossRef]
- Bachiller, E.; Irigoien, X. Trophodynamics and diet overlap of small pelagic fish species in the Bay of Biscay. Mar. Ecol. Prog. Ser. 2015, 534, 179–198. [Google Scholar] [CrossRef]
- Olaso, I.; Gutiérrez, J.L.; Villamor, B.; Carrera, P.; Valdés, L.; Abaunza, P. Seasonal changes in the north-eastern Atlantic mackerel diet (Scomber scombrus) in the north of Spain (ICES Division VIIIc). J. Mar. Biol. Assoc. U. K. 2005, 85, 415–418. [Google Scholar] [CrossRef]
- Cardona, L.; Alvarez de Quevedo, I.; Borrell, A.; Aguilar, A. Massive consumption of gelatinous plankton by Mediterranean apex predators. PLoS ONE 2012, 7, e31329. [Google Scholar] [CrossRef] [PubMed]
- Bjørdal, V.R.; Mørk, H.L.; Utne, K.R.; Fernö, A.; Nøttestad, L. The diet of juvenile Atlantic mackerel (Scomber scombrus) feeding in new northern nursery areas along the Norwegian coast. Mar. Biol. Res. 2022, 18, 415–425. [Google Scholar] [CrossRef]
- Šantić, M.; Rada, B.; Pallaoro, A. Diet of juveniles Mediterranean horse mackerel, Trachurus mediterraneus and horse mackerel, Trachurus trachurus (Carangidae), from the eastern central Adriatic. Cah. De Biol. Mar. 2013, 4, 41–48. [Google Scholar]
- Georgieva, Y.; Daskalov, G.M.; Klayn, S.; Stefanova, K.; Stefanova, E.S. Seasonal Diet and Feeding Strategy of Horse Mackerel Trachurus mediterraneus (Steindachner, 1868) (Perciformes: Carangidae) in the South-Western Black Sea. Acta Zool. Bulg. 2019, 71, 201. [Google Scholar]
- Fanelli, E.; Menicucci, S.; Malavolti, S.; De Felice, A.; Leonori, I. Mesoscale variations in the assemblage structure and trophodynamics of mesozooplankton communities of the Adriatic basin (Mediterranean Sea). Biogeosciences 2021, 1–33. [Google Scholar] [CrossRef]
- Costa, G.; MacKenzie, K.; Oliva, M.E. A Review of the Parasites Infecting Fishes of the Genus Trachurus (Pisces: Carangidae). Rev. Fish. Sci. Aquac. 2017, 25, 297–315. [Google Scholar] [CrossRef]
- Šantić, M.; Jardas, I.; Pallaoro, A. Feeding habits of horse mackerel, Trachurus trachurus (Linneaus, 1758), from the central Adriatic Sea. J. Appl. Ichthyol. 2005, 21, 125–130. [Google Scholar] [CrossRef]
- Jardas, I.; Šantić, M.; Pallaoro, A. Diet composition and feeding intensity of horse mackerel, Trachurus trachurus (Osteichthyes: Carangidae) in the eastern Adriatic. Mar. Biol. 2004, 144, 1051–1056. [Google Scholar] [CrossRef]
- Eigaard, O.R.; Bastardie, F.; Hintzen, N.T.; Buhl-Mortensen, L.; Buhl-Mortensen, P.; Catarino, R.; Dinesen, G.E.; Egekvist, J.; Fock, H.O.; Geitner, K.; et al. The footprint of bottom trawling in European waters: Distribution, intensity, and seabed integrity. ICES J. Mar. Sci. 2017, 74, 847–865. [Google Scholar] [CrossRef]
- Baptista, J.; Martinho, F.; Martins, R.; Carneiro, M.; Azevedo, M.; Vieira, A.R.; Gomes, P.; Pardal, M.A. Water temperature gradient shapes the structure and composition of nearshore marine fish communities in southern Europe. J. Sea Res. 2019, 154, 101807. [Google Scholar] [CrossRef]
- Milisenda, G.; Garofalo, G.; Fezzani, S.; Rjeibi, O.; Jarboui, O.; Chemmam, B.; Ceriola, L.; Bonanno, A.; Genovese, S.; Basilone, G.; et al. Biomass HotSpot distribution model and spatial interaction of two exploited species of horse mackerel in the south-central Mediterranean Sea. Hydrobiologia 2018, 821, 135–150. [Google Scholar] [CrossRef]
- Deudero, S. Interspecific trophic relationships among pelagic fish species underneath FADs. J. Fish Biol. 2001, 58, 53–67. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Community | |
|---|---|---|
| 1 | S. colias, S. scombrus, T. mediterraneus, T. trachurus | 1 unique community |
| 2 | North, Central and South Adriatic | S. colias, S. scombrus, T. mediterraneus, T. trachurus |
| Area | Position | Sex | Species | |||
|---|---|---|---|---|---|---|
| Scomber colias | Scomber scombrus | Trachurus mediterraneus | Trachurus trachurus | |||
| North Adriatic | Inshore | M | 32 | 3 | ||
| F | 2 | 37 | ||||
| ND | 9 | 3 | 6 | |||
| Offshore | M | 5 | 3 | |||
| F | ||||||
| ND | 1 | |||||
| Total | 5 (0) | 11 (5) | 76 (11) | 9 (2) | ||
| Central Adriatic | Inshore | M | 3 | |||
| F | 1 | 8 | ||||
| ND | 19 | 4 | 3 | |||
| Offshore | M | 2 | 1 | |||
| F | ||||||
| ND | 1 | 1 | 20 | |||
| Total | 22 (0) | 5 (1) | 13 (5) | 23 (4) | ||
| South Adriatic | Inshore | M | ||||
| F | ||||||
| ND | 13 | 4 | ||||
| Offshore | M | 1 | ||||
| F | 3 | |||||
| ND | 18 | 10 | ||||
| Total | 35 (0) | 0 | 4 (0) | 10 (2) | ||
| Total males | 8 | 0 | 39 | 3 | ||
| Total females | 3 | 3 | 45 | 0 | ||
| Total juveniles | 51 | 13 | 9 | 39 | ||
| Total individuals | 62 (0) | 16 (6) | 93 (16) | 42 (8) | ||
| SPECIES | Scomber colias | Scomber scombrus | Trachurus mediterraneus | Trachurus trachurus | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TA | 3.34 | 5.12 | 11.3 | 4.72 | |||||||
| SEAc | 1.24 | 2.26 | 3.3 | 1.65 | |||||||
| NR | 2.09 | 0.07 | 3.28 | 2.10 | |||||||
| CR | 0.35 | 0.71 | 0.55 | 0.33 | |||||||
| CD | 0.83 | 0.36 | 1.37 | 0.77 | |||||||
| MNND | 0.88 | 0.71 | 1.30 | 1.07 | |||||||
| SDNND | 0.63 | <0.05 | 1.31 | 0.08 | |||||||
| AREA | North | Central | South | North | Central | North | Central | South | North | Central | South |
| TA | 1.09 | 0.33 | 1.93 | 4.64 | 0.30 | 5.81 | 6.17 | 0.25 | 0.55 | 0.88 | 2.88 |
| SEAc | 1.42 | 0.32 | 1.21 | 2.73 | 0.39 | 2.21 | 5.92 | 0.33 | 0.45 | 1.03 | 1.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Ros, Z.; Fanelli, E.; Cassatella, S.; Biagiotti, I.; Canduci, G.; Menicucci, S.; De Felice, A.; Malavolti, S.; Leonori, I. Resource Partitioning among “Ancillary” Pelagic Fishes (Scomber spp., Trachurus spp.) in the Adriatic Sea. Biology 2023, 12, 272. https://doi.org/10.3390/biology12020272
Da Ros Z, Fanelli E, Cassatella S, Biagiotti I, Canduci G, Menicucci S, De Felice A, Malavolti S, Leonori I. Resource Partitioning among “Ancillary” Pelagic Fishes (Scomber spp., Trachurus spp.) in the Adriatic Sea. Biology. 2023; 12(2):272. https://doi.org/10.3390/biology12020272
Chicago/Turabian StyleDa Ros, Zaira, Emanuela Fanelli, Sacha Cassatella, Ilaria Biagiotti, Giovanni Canduci, Samuele Menicucci, Andrea De Felice, Sara Malavolti, and Iole Leonori. 2023. "Resource Partitioning among “Ancillary” Pelagic Fishes (Scomber spp., Trachurus spp.) in the Adriatic Sea" Biology 12, no. 2: 272. https://doi.org/10.3390/biology12020272
APA StyleDa Ros, Z., Fanelli, E., Cassatella, S., Biagiotti, I., Canduci, G., Menicucci, S., De Felice, A., Malavolti, S., & Leonori, I. (2023). Resource Partitioning among “Ancillary” Pelagic Fishes (Scomber spp., Trachurus spp.) in the Adriatic Sea. Biology, 12(2), 272. https://doi.org/10.3390/biology12020272