

The Distribution and Biogenic Origins of Zinc in the Mineralised Tooth Tissues of Modern and Fossil Hominoids: Implications for Life History, Diet and Taphonomy

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Zinc as an Essential Trace Element in the Body
1.2. Zinc in Enamel Secretion and Maturation
1.3. Zinc Absorption and Maternal–Fetal Transfer
1.4. The Distribution of Zinc in Mineralised Tissues

1.5. Zinc Incorporation into Hydroxyapatite
1.6. Zinc Isotopes and Diet in Fossils
1.7. Aims of the Study
2. Materials and Methods
2.1. Samples
2.2. Histological Methods
2.3. SXRF Methods
2.4. Statistical Methods
3. Results
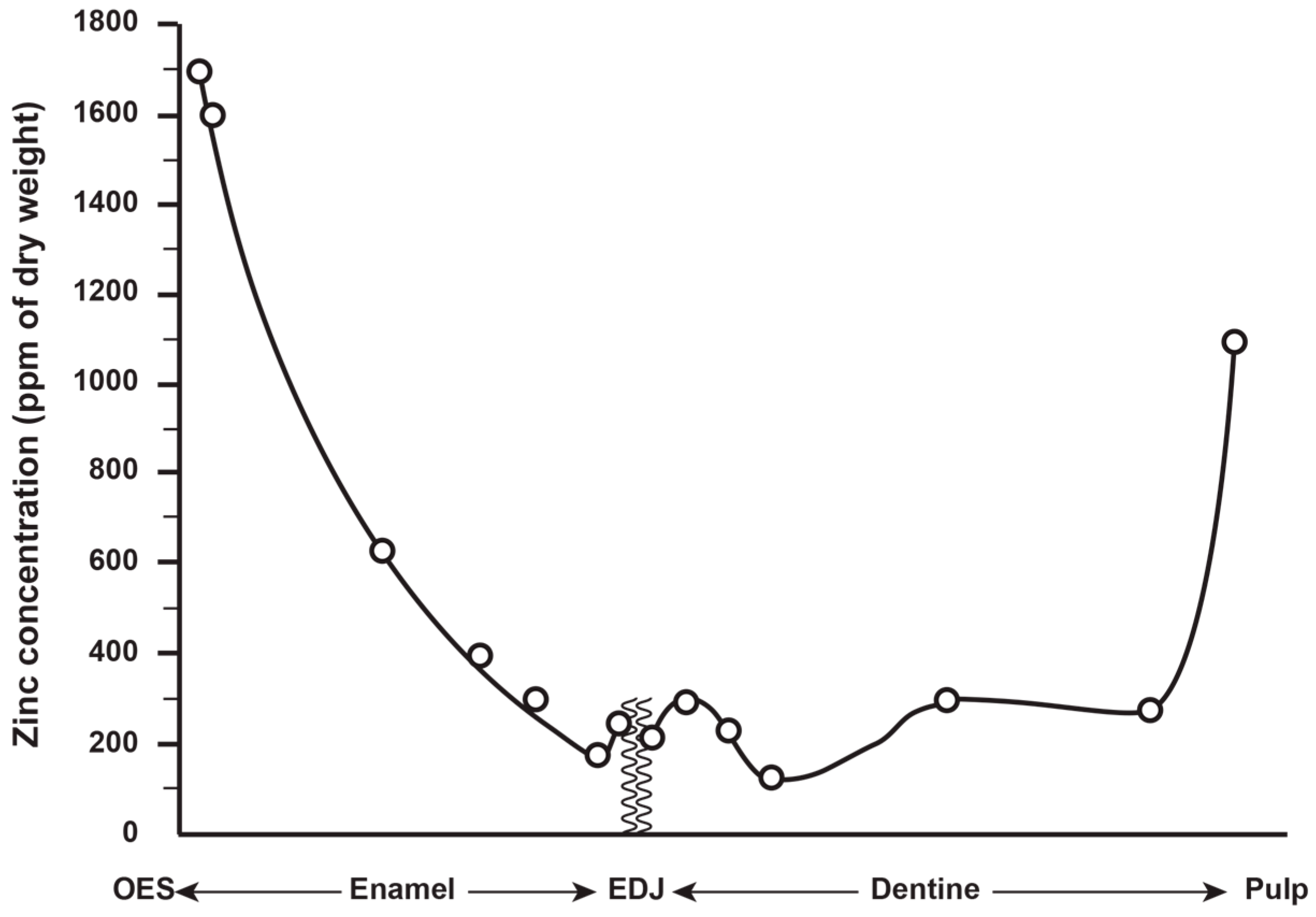
3.1. Zinc Distribution in Modern Human and Great Ape Deciduous Teeth
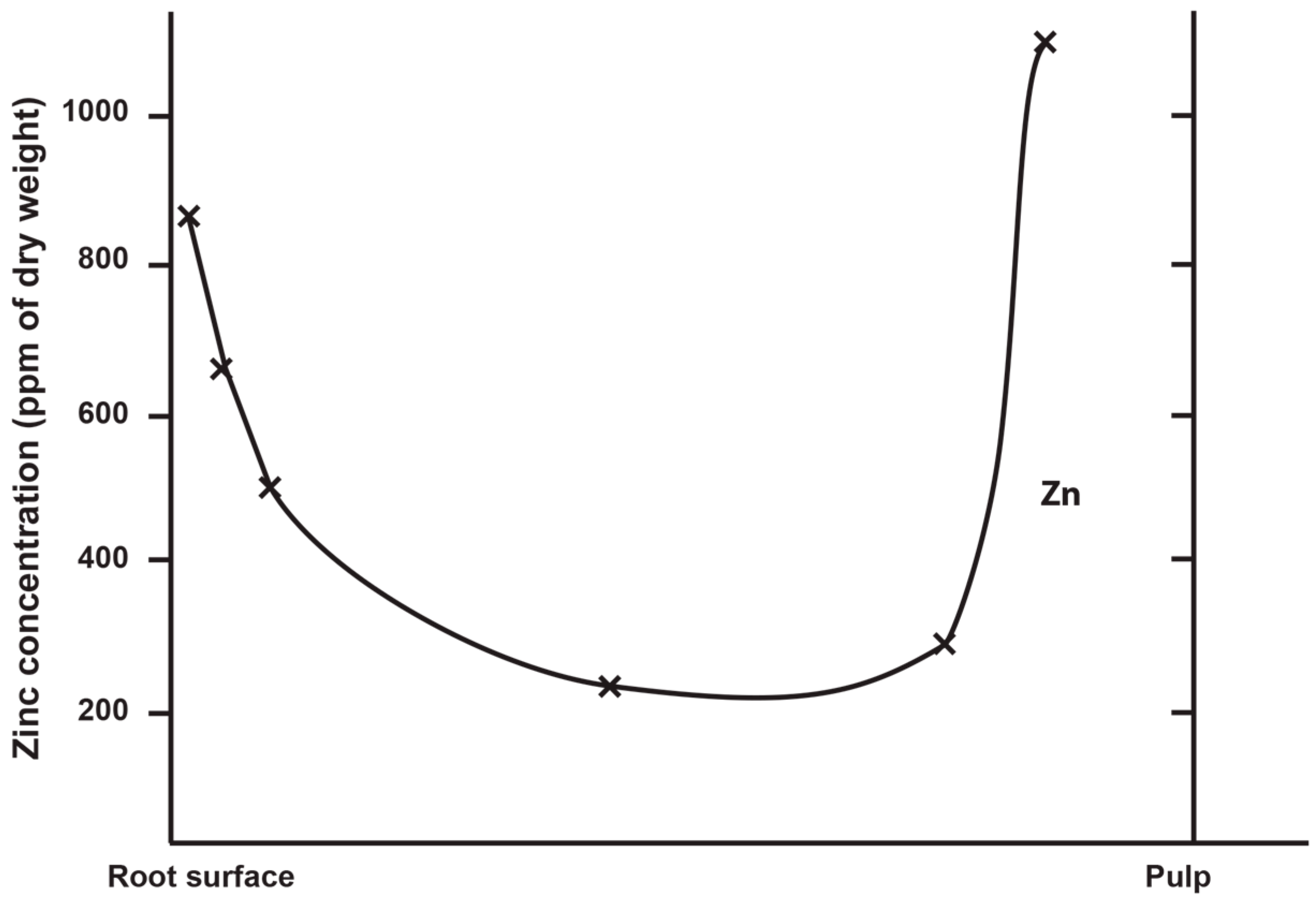
3.2. Zinc Distribution in Modern Human and Great Ape Permanent Teeth
3.3. Zinc Distribution in Permanent Teeth at Higher Resolution
3.4. Zinc Distribution and Diagenesis in Permanent Fossil Teeth




3.5. Zinc Distribution and Diagenesis in Deciduous Fossil Teeth

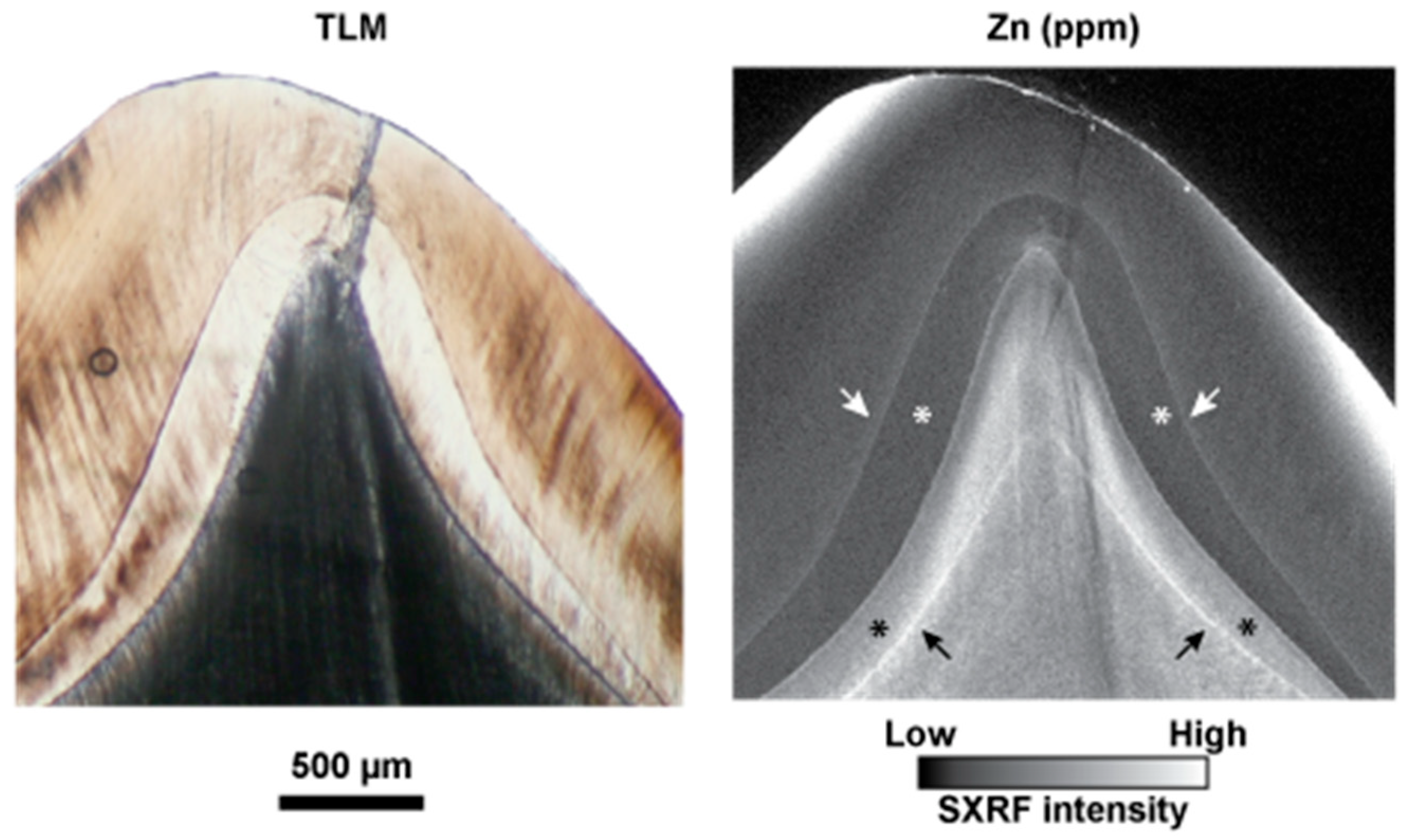
3.6. Zinc Distribution at the Neonatal Line in Fossil Teeth
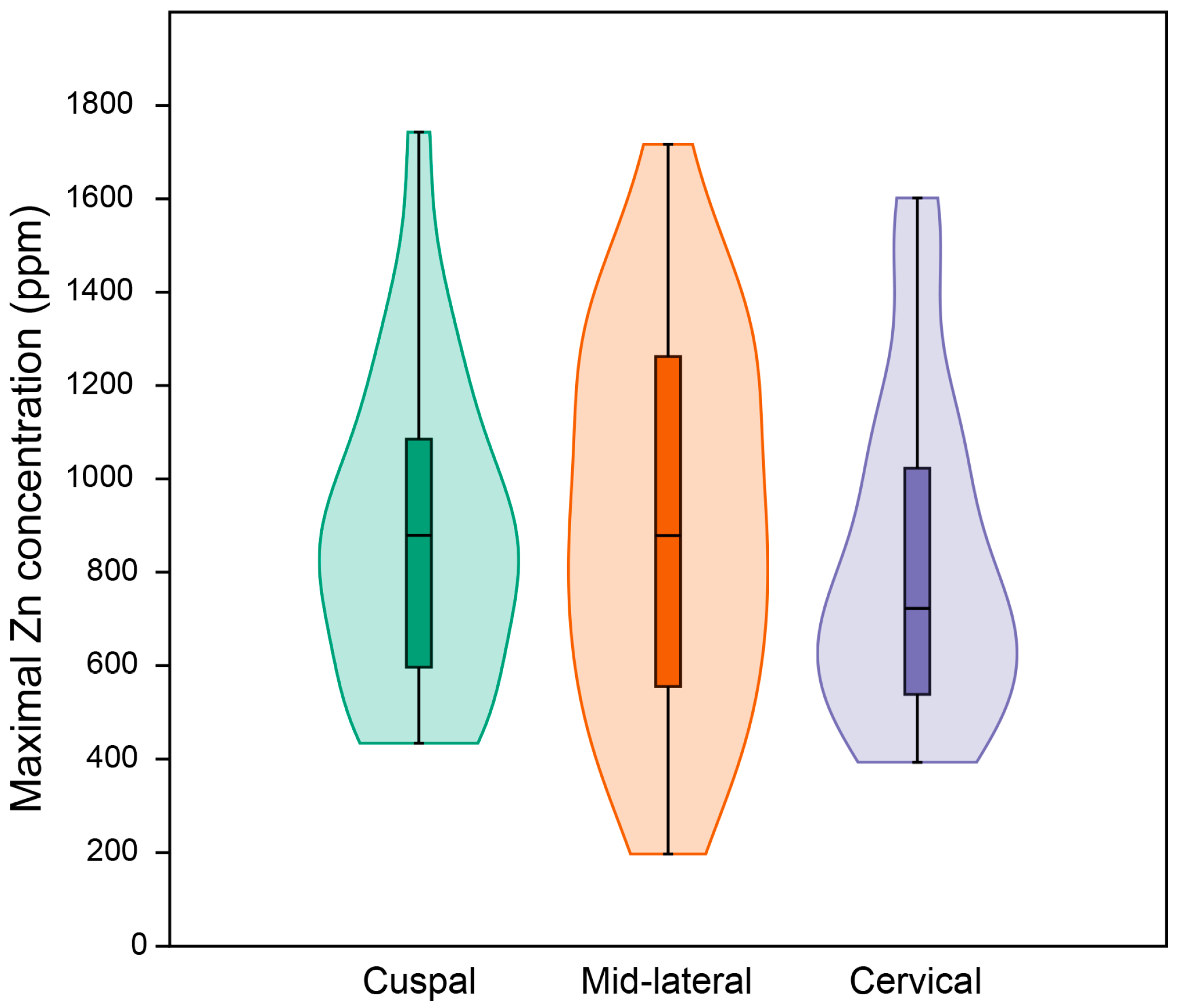
3.7. Quantification of Zinc Distribution in the Outer Enamel



4. Discussion
4.1. Zinc Incorporation into Enamel and Dentine
4.2. Zinc at the NNL and as a Marker of Stress Events
4.3. Quantification of Zinc Distribution in the Outer Enamel
4.4. Diagenetic Changes in Fossil Tooth Tissues
4.5. Sampling Zinc in Tooth Tissues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Terrin, G.; Berni Canani, R.; Di Chiara, M.; Pietravalle, A.; Aleandri, V.; Conte, F.; De Curtis, M. Zinc in Early Life: A Key Element in the Fetus and Preterm Neonate. Nutrients 2015, 7, 10427–10446. [Google Scholar] [CrossRef]
- Greenberg, S.R. The Histopathology of Clinically-Important Metals. A Review. Histol. Histopathol. 1989, 4, 375–380. [Google Scholar]
- Moonga, B.S.; Dempster, D.W. Zinc Is a Potent Inhibitor of Osteoclastic Bone Resorption in Vitro. J. Bone Miner. Res. 1995, 10, 453–457. [Google Scholar] [CrossRef]
- Lynch, R.J.M. Zinc in the Mouth, Its Interactions with Dental Enamel and Possible Effects on Caries; a Review of the Literature. Int. Dent. J. 2011, 61, 46–54. [Google Scholar] [CrossRef]
- Ganss, B.; Jheon, A. Zinc Finger Transcription Factors in Skeletal Development. Crit. Rev. Oral Biol. Med. 2004, 15, 282–297. [Google Scholar] [CrossRef]
- Maret, W. Escort Proteins for Cellular Zinc Ions. Nature 2022, 608, 38–39. [Google Scholar] [CrossRef]
- Brudevold, F.; Steadman, L.T.; Spinelli, M.A.; Amdur, B.H.; Grøn, P. A Study of Zinc in Human Teeth. Arch. Oral Biol. 1963, 8, 135–144. [Google Scholar] [CrossRef]
- Ten Cate, A.R. The Distribution of Alkaline Phosphatase in the Human Tooth Germ. Arch. Oral Biol. 1962, 7, 195–205. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Inamoto, K.; Suketa, Y. Effect of Essential Trace Metals on Bone Metabolism in Weanling Rats: Comparison with Zinc and Other Metals’ Actions. Res. Exp. Med. (Berl.) 1986, 186, 337–342. [Google Scholar] [CrossRef]
- Balter, V.; Lamboux, A.; Zazzo, A.; Télouk, P.; Leverrier, Y.; Marvel, J.; Moloney, A.P.; Monahan, F.J.; Schmidt, O.; Albarède, F. Contrasting Cu, Fe, and Zn Isotopic Patterns in Organs and Body Fluids of Mice and Sheep, with Emphasis on Cellular Fractionation. Metallomics 2013, 5, 1470–1482. [Google Scholar] [CrossRef]
- Klimuszko, E.; Orywal, K.; Sierpinska, T.; Sidun, J.; Golebiewska, M. The Evaluation of Zinc and Copper Content in Tooth Enamel without Any Pathological Changes—An in Vitro Study. Int. J. Nanomed. 2018, 13, 1257–1264. [Google Scholar] [CrossRef]
- Gomez, S.; Rizzo, R.; Pozzi-Mucelli, M.; Bonucci, E.; Vittur, F. Zinc Mapping in Bone Tissues by Histochemistry and Synchrotron Radiation–Induced X-ray Emission: Correlation with the Distribution of Alkaline Phosphatase. Bone 1999, 25, 33–38. [Google Scholar] [CrossRef]
- Kim, T.H.; Bae, C.H.; Lee, J.C.; Kim, J.E.; Yang, X.; de Crombrugghe, B.; Cho, E.S. Osterix Regulates Tooth Root Formation in a Site-Specific Manner. J. Dent. Res. 2015, 94, 430–438. [Google Scholar] [CrossRef]
- Lu, Y.; Papagerakis, P.; Yamakoshi, Y.; Hu, J.C.-C.; Bartlett, J.D.; Simmer, J.P. Functions of KLK4 and MMP-20 in Dental Enamel Formation. Biol. Chem. 2008, 389, 695–700. [Google Scholar] [CrossRef]
- Hubbard, M.J. Calcium Transport across the Dental Enamel Epithelium. Crit. Rev. Oral Biol. Med. 2000, 11, 437–466. [Google Scholar] [CrossRef]
- Reynard, B.; Balter, V. Trace Elements and Their Isotopes in Bones and Teeth: Diet, Environments, Diagenesis, and Dating of Archeological and Paleontological Samples. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 416, 4–16. [Google Scholar] [CrossRef]
- Müller, W.; Nava, A.; Evans, D.; Rossi, P.F.; Alt, K.W.; Bondioli, L. Enamel Mineralization and Compositional Time-Resolution in Human Teeth Evaluated via Histologically-Defined LA-ICPMS Profiles. Geochim. Cosmochim. Acta 2019, 255, 105–126. [Google Scholar] [CrossRef]
- Goldberg, M.; Septier, D.; Bourd, K.; Hall, R.; Jeanny, J.-C.; Jonet, L.; Colin, S.; Tager, F.; Chaussain-Miller, C.; Garabédian, M.; et al. The Dentino-Enamel Junction Revisited. Connect. Tissue Res. 2002, 43, 482–489. [Google Scholar] [CrossRef]
- Stock, S.R.; Finney, L.A.; Telser, A.; Maxey, E.; Vogt, S.; Okasinski, J.S. Cementum Structure in Beluga Whale Teeth. Acta Biomater. 2017, 48, 289–299. [Google Scholar] [CrossRef]
- Jeejeebhoy, K. Zinc: An Essential Trace Element for Parenteral Nutrition. Gastroenterology 2009, 137, S7–S12. [Google Scholar] [CrossRef]
- Jaouen, K.; Pouilloux, L.; Balter, V.; Pons, M.-L.; Hublin, J.-J.; Albarède, F. Dynamic Homeostasis Modeling of Zn Isotope Ratios in the Human Body. Metallomics 2019, 11, 1049–1059. [Google Scholar] [CrossRef]
- Jaouen, K.; Trost, M.; Bourgon, N.; Colleter, R.; Le Cabec, A.; Tütken, T.; Elias Oliveira, R.; Pons, M.L.; Méjean, P.; Steinbrenner, S.; et al. Zinc Isotope Variations in Archeological Human Teeth (Lapa Do Santo, Brazil) Reveal Dietary Transitions in Childhood and No Contamination from Gloves. PLoS ONE 2020, 15, e0232379. [Google Scholar] [CrossRef]
- Donangelo, C.M.; King, J.C. Maternal Zinc Intakes and Homeostatic Adjustments during Pregnancy and Lactation. Nutrients 2012, 4, 782–798. [Google Scholar] [CrossRef]
- Takeuchi, K.; Nakagaki, H.; Toyama, Y.; Kimata, N.; Ito, F.; Robinson, C.; Weatherell, J.A.; Stösser, L.; Künzel, W. Fluoride Concentrations and Distribution in Premolars of Children from Low and Optimal Fluoride Areas. Caries Res. 1996, 30, 76–82. [Google Scholar] [CrossRef]
- Ide-Ektessabi, A.; Shirasawa, K.; Koizumi, A.; Azechi, M. Application of Synchrotron Radiation Microbeams to Environmental Monitoring. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2004, 213, 761–765. [Google Scholar] [CrossRef]
- Brozou, A.; Mannino, M.A.; Van Malderen, S.J.M.; Garrevoet, J.; Pubert, E.; Fuller, B.T.; Dean, M.C.; Colard, T.; Santos, F.; Lynnerup, N.; et al. Using SXRF and LA-ICP-TOFMS to Explore Evidence of Treatment and Physiological Responses to Leprosy in Medieval Denmark. Biology 2023, 12, 184. [Google Scholar] [CrossRef]
- Obtel, N.; Le Cabec, A.; Nguyen, T.N.; Giabicani, E.; Van Malderen, S.J.M.; Garrevoet, J.; Percot, A.; Paris, C.; Dean, C.; Hadj-Rabia, S.; et al. Impact of Claudin-10 Deficiency on Amelogenesis: Lesson from a HELIX Tooth. Ann. N. Y. Acad. Sci. 2022, 1516, 197–211. [Google Scholar] [CrossRef]
- Sánchez-Quevedo, M.; Crespo, P.; García, J.; Campos, A. X-ray Histochemistry of Zinc in Dental Tissues. Eur. Arch. Biol. 1992, 103, 47–49. [Google Scholar]
- Humphrey, L.T.; Jeffries, T.E.; Dean, M.C. Micro Spatial Distributions of Lead and Zinc in Human Deciduous Tooth Enamel. In Technique and Application in Dental Anthropology; Irish, J.D., Nelson, G.C., Eds.; Studies in Biological Anthropology; Cambridge University Press: Cambridge, UK, 2008; Volume 53, pp. 87–110. [Google Scholar]
- Humphrey, L.T.; Hassett, B.R.; Jeffries, T.E.; Dean, C.M. Trace Element Distributions in Early and Late Forming Tooth Enamel. In Proceedings of the 4th Meeting of the European Society for the Study of Human Evolution, Florence, Italy, 18–20 September 2014; p. 88. [Google Scholar]
- Humphrey, L.T.; Hassett, B.R.; Jeffries, T.E.; Dean, C.M. Micro-spatial Patterning of Lead and Zinc in Deciduous Tooth Enamel. In Proceedings of the 17th International Symposium on Dental Morphology, Bordeaux, France, 4–7 October 2017; p. 170. [Google Scholar]
- Bourgon, N.; Jaouen, K.; Bacon, A.-M.; Jochum, K.P.; Dufour, E.; Duringer, P.; Ponche, J.-L.; Joannes-Boyau, R.; Boesch, Q.; Antoine, P.-O.; et al. Zinc Isotopes in Late Pleistocene Fossil Teeth from a Southeast Asian Cave Setting Preserve Paleodietary Information. Proc. Natl. Acad. Sci. USA 2020, 117, 4675. [Google Scholar] [CrossRef]
- Kang, D.; Amarasiriwardena, D.; Goodman, A.H. Application of Laser Ablation-Inductively Coupled Plasma-Mass Spectrometry (LA-ICP-MS) to Investigate Trace Metal Spatial Distributions in Human Tooth Enamel and Dentine Growth Layers and Pulp. Anal. Bioanal. Chem. 2004, 378, 1608–1615. [Google Scholar] [CrossRef]
- Dean, M.C.; Spiers, K.M.; Garrevoet, J.; Le Cabec, A. Synchrotron X-ray Fluorescence Mapping of Ca, Sr and Zn at the Neonatal Line in Human Deciduous Teeth Reflects Changing Perinatal Physiology. Arch. Oral Biol. 2019, 104, 90–102. [Google Scholar] [CrossRef]
- Stock, S.R.; Deymier-Black, A.C.; Veis, A.; Telser, A.; Lux, E.; Cai, Z. Bovine and Equine Peritubular and Intertubular Dentin. Biomineralization 2014, 10, 3969–3977. [Google Scholar] [CrossRef]
- Dean, M.C.; Le Cabec, A.; Spiers, K.; Zhang, Y.; Garrevoet, J. Incremental Distribution of Strontium and Zinc in Great Ape and Fossil Hominin Cementum Using Synchrotron X-ray Fluorescence Mapping. J. R. Soc. Interface 2018, 15, 20170626. [Google Scholar] [CrossRef]
- Brudevold, F.; Steadman, L.T.; Smith, F.A. Inorganic and Organic Components of Tooth Structure. Ann. N. Y. Acad. Sci. 1960, 85, 110–132. [Google Scholar] [CrossRef]
- Martin, R.R.; Naftel, S.J.; Nelson, A.J.; Feilen, A.B.; Narvaez, A. Synchrotron X-ray Fluorescence and Trace Metals in the Cementum Rings of Human Teeth. J. Environ. Monit. 2004, 6, 783–786. [Google Scholar] [CrossRef]
- Martin, R.R.; Naftel, S.J.; Nelson, A.J.; Sapp III, W.D. Comparison of the Distributions of Bromine, Lead, and Zinc in Tooth and Bone from an Ancient Peruvian Burial Site by X-ray Fluorescence. Can. J. Chem. 2007, 85, 831–836. [Google Scholar] [CrossRef]
- Stock, S.R.; Veis, A.; Telser, A.; Cai, Z. Near Tubule and Intertubular Bovine Dentin Mapped at the 250 Nm Level. J. Struct. Biol. 2011, 176, 203–211. [Google Scholar] [CrossRef]
- Djomehri, S.I.; Candell, S.; Case, T.; Browning, A.; Marshall, G.W.; Yun, W.; Lau, S.H.; Webb, S.; Ho, S.P. Mineral Density Volume Gradients in Normal and Diseased Human Tissues. PLoS ONE 2015, 10, e0121611. [Google Scholar] [CrossRef]
- Klevezal, G.A.; Kirillova, I.V.; Shishlina, N.I.; Sokolov, A.A.; Trunova, Y.E. Growth Layers in Tooth Dentin and Cementum: Problems and Perspectives of Their Use in the Study of Fossil and Subfossil Mammal Remains Including Humans. Doc. Archaeobiol. 2006, 4, 113–124. [Google Scholar]
- Le Cabec, A.; Tang, N.K.; Ruano Rubio, V.; Hillson, S. Nondestructive Adult Age at Death Estimation: Visualizing Cementum Annulations in a Known Age Historical Human Assemblage Using Synchrotron X-ray Microtomography. Am. J. Phys. Anthropol. 2019, 168, 25–44. [Google Scholar] [CrossRef]
- Cerrito, P.; Bailey, S.E.; Hu, B.; Bromage, T.G. Parturitions, Menopause and Other Physiological Stressors Are Recorded in Dental Cementum Microstructure. Sci. Rep. 2020, 10, 5381. [Google Scholar] [CrossRef]
- Seo, H.-J.; Cho, Y.-E.; Kim, T.; Shin, H.-I.; Kwun, I.-S. Zinc May Increase Bone Formation through Stimulating Cell Proliferation, Alkaline Phosphatase Activity and Collagen Synthesis in Osteoblastic MC3T3-E1 Cells. Nutr. Res. Pract. 2010, 4, 356. [Google Scholar] [CrossRef]
- Mansell, R.E.; Hendershot, L.C. The Spectrochemical Analysis of Metals in Rat Molar Enamel, Femurs and Incisors. Arch. Oral Biol. 1960, 2, 31–37. [Google Scholar] [CrossRef]
- Sasaki, T.; Takagi, M.; Yanagisawa, T. Structure and Function of Secretory Ameloblasts in Enamel Formation. In Proceedings of the Ciba Foundation Symposium 205; Chadwick, D.J., Cardew, G., Eds.; Novartis Foundation Symposia; John Wiley & Sons, Ltd.: Chichester, UK, 1997; pp. 32–53. ISBN 978-0-470-51530-3. [Google Scholar]
- Featherstone, J.D.B.; Nelson, D.G.A. The Effect of Fluoride, Zinc, Strontium, Magnesium and Iron on the Crystal-Structural Disorder in Synthetic Carbonated Apatites. Aust. J. Chem. 1980, 33, 2363. [Google Scholar] [CrossRef]
- Crawford, A.W.; De Bruin, H.J. Concentration Changes in Surface Ca, P, F, Zn, Fe, and Sr During White Spot Formation. J. Dent. Res. 1983, 62, 964–968. [Google Scholar] [CrossRef]
- Mohammed, N.R.; Mneimne, M.; Hill, R.G.; Al-Jawad, M.; Lynch, R.J.M.; Anderson, P. Physical Chemical Effects of Zinc on in Vitro Enamel Demineralization. J. Dent. 2014, 42, 1096–1104. [Google Scholar] [CrossRef]
- Yamaguchi, M. Role of Nutritional Zinc in the Prevention of Osteoporosis. Mol. Cell. Biochem. 2010, 338, 241–254. [Google Scholar] [CrossRef]
- Mayer, I.; Apfelbaum, F.; Featherstone, J.D.B. Zinc Ions in Synthetic Carbonated Hydroxyapatites. Arch. Oral Biol. 1994, 39, 87–90. [Google Scholar] [CrossRef]
- Fuierer, T.A.; LoRe, M.; Puckett, S.A.; Nancollas, G.H. A Mineralization Adsorption and Mobility Study of Hydroxyapatite Surfaces in the Presence of Zinc and Magnesium Ions. Langmuir 1994, 10, 4721–4725. [Google Scholar] [CrossRef]
- Van Heghe, L.; Engström, E.; Rodushkin, I.; Cloquet, C.; Vanhaecke, F. Isotopic Analysis of the Metabolically Relevant Transition Metals Cu, Fe and Zn in Human Blood from Vegetarians and Omnivores Using Multi-Collector ICP-Mass Spectrometry. J. Anal. At. Spectrom. 2012, 27, 1327. [Google Scholar] [CrossRef]
- Jaouen, K.; Beasley, M.; Schoeninger, M.; Hublin, J.-J.; Richards, M.P. Zinc Isotope Ratios of Bones and Teeth as New Dietary Indicators: Results from a Modern Food Web (Koobi Fora, Kenya). Sci. Rep. 2016, 6, 26281. [Google Scholar] [CrossRef]
- Jaouen, K.; Szpak, P.; Richards, M.P. Zinc Isotope Ratios as Indicators of Diet and Trophic Level in Arctic Marine Mammals. PLoS ONE 2016, 11, e0152299. [Google Scholar] [CrossRef]
- Jaouen, K.; Villalba-Mouco, V.; Smith, G.M.; Trost, M.; Leichliter, J.; Lüdecke, T.; Méjean, P.; Mandrou, S.; Chmeleff, J.; Guiserix, D.; et al. A Neandertal Dietary Conundrum: Insights Provided by Tooth Enamel Zn Isotopes from Gabasa, Spain. Proc. Natl. Acad. Sci. USA 2022, 119, e2109315119. [Google Scholar] [CrossRef]
- Bourgon, N.; Jaouen, K.; Bacon, A.-M.; Dufour, E.; McCormack, J.; Tran, N.-H.; Trost, M.; Fiorillo, D.; Dunn, T.E.; Zanolli, C.; et al. Trophic Ecology of a Late Pleistocene Early Modern Human from Tropical Southeast Asia Inferred from Zinc Isotopes. J. Hum. Evol. 2021, 161, 103075. [Google Scholar] [CrossRef]
- Jaouen, K.; Herrscher, E.; Balter, V. Copper and Zinc Isotope Ratios in Human Bone and Enamel. Am. J. Phys. Anthropol. 2017, 162, 491–500. [Google Scholar] [CrossRef]
- Jaouen, K.; Pons, M.-L. Potential of Non-Traditional Isotope Studies for Bioarchaeology. Archaeol. Anthropol. Sci. 2017, 9, 1389–1404. [Google Scholar] [CrossRef]
- Weber, K.; Weber, M.; Menneken, M.; Kral, A.G.; Mertz-Kraus, R.; Geisler, T.; Vogl, J.; Tütken, T. Diagenetic Stability of Non-Traditional Stable Isotope Systems (Ca, Sr, Mg, Zn) in Teeth—An in-Vitro Alteration Experiment of Biogenic Apatite in Isotopically Enriched Tracer Solution. Chem. Geol. 2021, 572, 120196. [Google Scholar] [CrossRef]
- Dean, M.C.; Elamin, F. Parturition Lines in Modern Human Wisdom Tooth Roots: Do They Exist, Can They Be Characterized and Are They Useful for Retrospective Determination of Age at First Reproduction and/or Inter-Birth Intervals? Ann. Hum. Biol. 2014, 41, 358–367. [Google Scholar] [CrossRef]
- Dean, M.C.; Liversidge, H.M.; Elamin, F. Combining Radiographic and Histological Data for Dental Development to Compare Growth in the Past and the Present. Ann. Hum. Biol. 2014, 41, 336–347. [Google Scholar] [CrossRef]
- Dean, M.C. Hunters, Dealers, Collectors and Scientists and the Origins of the Elliot Smith Primate Collection at University College London. Dent. Hist. 2022, 67, 1–9. [Google Scholar]
- Dean, M.C.; Le Cabec, A.; Van Malderen, S.J.M.; Garrevoet, J. Synchrotron X-ray Fluorescence Imaging of Strontium Incorporated into the Enamel and Dentine of Wild-Shot Orangutan Canine Teeth. Arch. Oral Biol. 2020, 119, 104879. [Google Scholar] [CrossRef]
- Beynon, A.D.; Dean, M.C.; Leakey, M.G.; Reid, D.J.; Walker, A. Comparative Dental Development and Microstructure of Proconsul Teeth from Rusinga Island, Kenya. J. Hum. Evol. 1998, 35, 163–209. [Google Scholar] [CrossRef]
- Dean, M.C.; Kelley, J. Comparative Dental Development in Hispanopithecus laietanus and Pan troglodytes. J. Hum. Evol. 2012, 62, 174–178. [Google Scholar] [CrossRef]
- Ward, C.V.; Leakey, M.G.; Walker, A. Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. J. Hum. Evol. 2001, 41, 255–368. [Google Scholar] [CrossRef]
- Dean, M.C.; Zanolli, C.; Le Cabec, A.; Tawane, M.; Garrevoet, J.; Mazurier, A.; Macchiarelli, R. Growth and Development of the Third Permanent Molar in Paranthropus robustus from Swartkrans, South Africa. Sci. Rep. 2020, 10, 19053. [Google Scholar] [CrossRef]
- Rink, W.J.; Schwarcz, H.P.; Lee, H.K.; Rees-Jones, J.; Rabinovich, R.; Hovers, E. Electron Spin Resonance (ESR) and Thermal Ionization Mass Spectrometric (TIMS) 230Th/234U Dating of Teeth in Middle Paleolithic Layers at Amud Cave, Israel. Geoarchaeology 2001, 16, 701–717. [Google Scholar] [CrossRef]
- Rak, Y.; Kimbel, W.H.; Hovers, E. A Neandertal Infant from Amud Cave, Israel. J. Hum. Evol. 1994, 26, 313–324. [Google Scholar] [CrossRef]
- Hovers, E.; Rak, Y.; Lavi, R.; Kimbel, W.H. Hominid Remains from Amud Cave in the Context of the Levantine Middle Paleolithic. Paléorient 1995, 21, 47–61. [Google Scholar] [CrossRef]
- Stringer, C.B.; Grün, R.; Schwarcz, H.P.; Goldberg, P. ESR Dates for the Hominid Burial Site of Es Skhul in Israel. Nature 1989, 338, 756–758. [Google Scholar] [CrossRef]
- Mercier, N.; Valladas, H.; Bar-Yosef, O.; Vandermeersch, B.; Stringer, C.; Joron, J.-L. Thermoluminescence Date for the Mousterian Burial Site of Es-Skhul, Mt. Carmel. J. Archaeol. Sci. 1993, 20, 169–174. [Google Scholar] [CrossRef]
- Dean, M.C.; Garrevoet, J.; Spiers, K.M.; Rak, Y.; Lahr, M.M.; Foley, R.A.; Le Cabec, A. The Distribution of Zinc in Modern and Fossil Enamel, Dentine and Cementum. In Proceedings of the 9th Meeting of the European Society for the Study of Human Evolution 8, Liège, Belgium, 19–21 September 2019; p. 39. [Google Scholar]
- Schroer, C.G.; Boye, P.; Feldkamp, J.M.; Patommel, J.; Samberg, D.; Schropp, A.; Schwab, A.; Stephan, S.; Falkenberg, G.; Wellenreuther, G.; et al. Hard X-ray Nanoprobe at Beamline P06 at PETRA III. X-ray Mirror 2010, 616, 93–97. [Google Scholar] [CrossRef]
- Boesenberg, U.; Ryan, C.G.; Kirkham, R.; Siddons, D.P.; Alfeld, M.; Garrevoet, J.; Nunez, T.; Claussen, T.; Kracht, T.; Falkenberg, G. Fast X-ray Microfluorescence Imaging with Submicrometer-Resolution Integrating a Maia Detector at Beamline P06 at PETRA III. J. Synchrotron Radiat. 2016, 23, 1550–1560. [Google Scholar] [CrossRef]
- Kirkham, R.; Dunn, P.A.; Kuczewski, A.J.; Siddons, D.P.; Dodanwela, R.; Moorhead, G.F.; Ryan, C.G.; Geronimo, G.D.; Beuttenmuller, R.; Pinelli, D.; et al. The Maia Spectroscopy Detector System: Engineering for Integrated Pulse Capture, Low-Latency Scanning and Real-Time Processing. AIP Conf. Proc. 2010, 1234, 240–243. [Google Scholar] [CrossRef]
- Weidmann, S.M.; Weatherell, J.A.; Hamm, S.M. Variations of Enamel Density in Sections of Human Teeth. Arch. Oral Biol. 1967, 12, 85–97. [Google Scholar] [CrossRef]
- Solé, V.A.; Papillon, E.; Cotte, M.; Walter, P.; Susini, J. A Multiplatform Code for the Analysis of Energy-Dispersive X-ray Fluorescence Spectra. Spectrochim. Acta Part B At. Spectrosc. 2007, 62, 63–68. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol Electron. 2001, 4, 1–9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Schulte, E.; Davison, D.; Dye, T.; Dominik, C. A Multi-Language Computing Environment for Literate Programming and Reproducible Research. J. Stat. Softw. 2012, 46, 1–24. [Google Scholar] [CrossRef]
- Simonsohn, U.; Gruson, H. Groundhog: Reproducible Scripts via Version-Specific Package Loading 2021. Available online: https://CRAN.R-project.org/package=groundhog (accessed on 1 November 2023).
- Birch, W.; Dean, M.C. A Method of Calculating Human Deciduous Crown Formation Times and of Estimating the Chronological Ages of Stressful Events Occurring during Deciduous Enamel Formation. J. Forensic Leg. Med. 2014, 22, 127–144. [Google Scholar] [CrossRef]
- Le Cabec, A.; Garrevoet, J.; Spiers, K.M.; Dean, M.C. Incremental Elemental Distribution in Chimpanzee Cellular Cementum: Insights from Synchrotron X-ray Fluorescence and Implications for Life-History Inferences. In Dental Cementum in Anthropology; Gourichon, L., Naji, S., Rendu, W., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 138–154. ISBN 978-1-108-47708-6. [Google Scholar]
- McNulty, K.P.; Begun, D.R.; Kelley, J.; Manthi, F.K.; Mbua, E.N. A Systematic Revision of Proconsul with the Description of a New Genus of Early Miocene Hominoid. J. Hum. Evol. 2015, 84, 42–61. [Google Scholar] [CrossRef]
- Humphrey, L.T.; Dean, M.C.; Jeffries, T.E.; Penn, M. Unlocking Evidence of Early Diet from Tooth Enamel. Proc. Natl. Acad. Sci. USA 2008, 105, 6834–6839. [Google Scholar] [CrossRef]
- de Winter, N.J.; Snoeck, C.; Claeys, P. Seasonal Cyclicity in Trace Elements and Stable Isotopes of Modern Horse Enamel. PLoS ONE 2016, 11, e0166678. [Google Scholar] [CrossRef]
- Anczkiewicz, R.; Nava, A.; Bondioli, L.; Müller, W.; Spötl, C.; Koziarska, M.; Boczkowska, M.; Wojtal, P.; Wilczyński, J. High Spatial Resolution Sr Isotope and Trace Element Record of Dental Enamel Mineralization in a Woolly Mammoth Tooth: Implications for Paleoecological Reconstructions. Quat. Sci. Rev. 2023, 313, 108191. [Google Scholar] [CrossRef]
- Green, D.R.; Smith, T.M.; Green, G.M.; Bidlack, F.B.; Tafforeau, P.; Colman, A.S. Quantitative Reconstruction of Seasonality from Stable Isotopes in Teeth. Geochim. Cosmochim. Acta 2018, 235, 483–504. [Google Scholar] [CrossRef]
- Kohn, M.J.; Morris, J.; Olin, P. Trace Element Concentrations in Teeth—A Modern Idaho Baseline with Implications for Archeometry, Forensics, and Palaeontology. J. Archaeol. Sci. 2013, 40, 1689–1699. [Google Scholar] [CrossRef]
- Brown, C.J.; Chenery, S.R.N.; Smith, B.; Mason, C.; Tomkins, A.; Roberts, G.J.; Sserunjogi, L.; Tiberindwa, J.V. Environmental Influences on the Trace Element Content of Teeth—Implications for Disease and Nutritional Status. Arch. Oral Biol. 2004, 49, 705–717. [Google Scholar] [CrossRef]
- Stoorvogel, J.J.; Van Breemen, N.; Jassen, B.H. The Nutrient Input by Harmattan Dust to a Forest Ecosystem in Côte d’Ivoire, Africa. Biogeochemistry 1997, 37, 145–157. [Google Scholar] [CrossRef]
- Ziegler, J.L. Geophagy: A Vestige of Palaeonutrition? Trop. Med. Int. Health 1997, 2, 609–611. [Google Scholar] [CrossRef]
- Boyde, A. Carbonate Concentration, Crystal Centers, Core Dissolution, Caries, Cross Striations, Circadian Rhythms, and Compositional Contrast in the SEM. J. Dent. Res. 1979, 58, 981–983. [Google Scholar] [CrossRef]
- Boyde, A. Enamel. In Teeth; Berkovitz, B.K.B., Boyde, A., Frank, R.M., Höhling, H.J., Moxham, B.J., Nalbandian, J., Tonge, C.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 309–473. ISBN 978-3-642-83496-7. [Google Scholar]
- Sasaki, T.; Otsu, K.; Fujiwara, N.; Xu, L.; Taniguchi, A.; Harada, H. New Hypothesis of Cross-Striation Formation Mechanism. In Proceedings of the 10th Meeting of the TDM: Tooth Morphogenesis and Differentiation—Ameloblasts and Amelogenesis, Berlin, Germany, 1–4 September 2010. [Google Scholar]
- Weatherell, J.A.; Deutsch, D.; Robinson, C.; Hallsworth, A.S. Assimilation of Fluoride by Enamel throughout the Life of the Tooth. Caries Res. 1977, 11, 85–115. [Google Scholar] [CrossRef]
- Robinson, C. Enamel Maturation: A Brief Background with Implications for Some Enamel Dysplasias. Front. Physiol. 2014, 5, 388. [Google Scholar] [CrossRef]
- Robinson, C.; Kirkham, J.; Brookes, S.J.; Bonass, W.A.; Shore, R.C. The Chemistry of Enamel Development. Int. J. Dev. Biol. 1995, 39, 145–152. [Google Scholar]
- Robinson, C.; Brookes, S.J.; Bonass, W.A.; Shore, R.C.; Kirkham, J. Enamel Maturation. In Proceedings of the Ciba Foundation Symposium 205; Chadwick, D.J., Cardew, G., Eds.; Novartis Foundation Symposia; John Wiley & Sons, Ltd.: Chichester, UK, 1997; pp. 156–174. ISBN 978-0-470-51530-3. [Google Scholar]
- Nelson, D.G.A.; Featherstone, J.D.B.; Duncan, J.F.; Cutress, T.W. Paracrystalline Disorder of Biological and Synthetic Carbonate-Substituted Apatites. J. Dent. Res. 1982, 61, 1274–1281. [Google Scholar] [CrossRef]
- Lippert, F.; Hara, A.T. Strontium and Caries: A Long and Complicated Relationship. Caries Res. 2013, 47, 34–49. [Google Scholar] [CrossRef]
- Ressler, A.; Žužić, A.; Ivanišević, I.; Kamboj, N.; Ivanković, H. Ionic Substituted Hydroxyapatite for Bone Regeneration Applications: A Review. Open Ceram. 2021, 6, 100122. [Google Scholar] [CrossRef]
- Arora, M.; Kennedy, B.J.; Ryan, C.G.; Boadle, R.A.; Walker, D.M.; Harland, C.L.; Lai, B.; Cai, Z.; Vogt, S.; Zoellner, H.; et al. The Application of Synchrotron Radiation Induced X-ray Emission in the Measurement of Zinc and Lead in Wistar Rat Ameloblasts. Arch. Oral Biol. 2007, 52, 938–944. [Google Scholar] [CrossRef]
- Kuzawa, C.W. Adipose Tissue in Human Infancy and Childhood: An Evolutionary Perspective. Am. J. Phys. Anthropol. 1998, 107, 177–209. [Google Scholar] [CrossRef]
- Foley, R.A.; Lee, P.C. Ecology and Energetics of Encephalization in Hominid Evolution. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1991, 334, 223–232. [Google Scholar] [CrossRef]
- Richards, A.; Fejerskov, O.; Baelum, V. Enamel Fluoride in Relation To Severity of Human Dental Fluorosis. Adv. Dent. Res. 1989, 3, 147–153. [Google Scholar] [CrossRef]
- Iijima, Y.; Katayama, T. Fluoride Uptake by Deciduous Enamel from Fluoride Mouthrinse and Natural Fluoride in the Water. Stud. Environ. Sci. 1986, 27, 369–378. [Google Scholar] [CrossRef]
- Dean, M.C.; Garrevoet, J.; Van Malderen SJ, M.; Santos, F.; Mirazón Lahr, M.; Foley, R.; Le Cabec, A. The Distribution and Biogenic Origins of Zinc in the Mineralised Tooth Tissues of Modern and Fossil Hominoids: Implications for Life History, Diet and Taphonomy. Zenodo 2023. [Google Scholar] [CrossRef]
- Diedenhofen, B.; Musch, J. Cocor: A Comprehensive Solution for the Statistical Comparison of Correlations. PLOS ONE 2015, 10, e0121945. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Specimen | Specimen Code (Graphs) | Site/Collection | Tooth | Thin Section Thickness (µm) | Resolution (µm) | Dwell Time (ms) | Total Scan Time |
|---|---|---|---|---|---|---|---|---|
| Fossils | ||||||||
| Ekembo nyanzae | KNM-RU 1721 | - | Rusinga Island, Kenya | URM1 | 120 | 15 | 10 | 55 min |
| Ekembo heseloni | KNM-RU individual IV | S89 | Rusinga Island, Kenya | LRdm2 | 120 | 15 1.5 | 10 | 36 min 1.9 h |
| Ekembo heseloni | KNM-RU individual IV | S90 | Rusinga Island, Kenya | LLM1 | 110 | 10 1.5 | 4 10 | 28 min 1.9 h |
| Hispanopithecus laietanus | IPS 1781 | S46 | Can Llobateres, Spain | UM1 | 100 | 15 | 10 | 2.3 h |
| Australopithecus anamensis | KNM-KP 30748 | S205 | Kanapoi, Kenya | LP4 (frag.) | 100 | 10 | 10 | 3.1 h |
| Paranthropus robustus | SK 835 | S40 | Swartkrans, South Africa | UM3 | 100 | 10 | 10 | 2.3 h |
| Neanderthal | Amud 7 | S44 | Amud Cave, Israel | Udm1 | 100 | 10 2.7 | 10 3 | 2.3 h 2.2 h |
| early Homo sapiens | Skhūl IV | S45 | Skhūl, Wadi el-Mughara, Mount Carmel, Israel | LLM3 | 100 | 15 | 10 | 2.8 h |
| Modern specimens | ||||||||
| Pongo | UCL-CA28JS7 | S16 | UCL Elliot Smith | URdm2 | 80–100 | 15 | 10 | 1.9 h |
| Pongo | UCL-CA28JS7 | S18 | UCL Elliot Smith | URdm2 | 80–100 | 12 | 10 | 1.9 h |
| Pongo | UCL-CA28JS3 | S23 | UCL Elliot Smith | ULC | 100 | 25 | 10 | 3.0 h |
| Gorilla | UCL-CA1F1472 | S19 | UCL Elliot Smith | LRdm2 | 80–100 | 15 | 10 | 2.1 h |
| Gorilla | UCL-CA1F1472 | S20 | UCL Elliot Smith | LLdc | 100 | 15 | 10 | 1.5 h |
| Gorilla | UCL-CA4 | S27 | UCL Elliot Smith | LRM2 | 100 | 20 | 10 | 2.6 h |
| Gorilla | UCL-CA4 | S28 | UCL Elliot Smith | LRM2 | 100 | 20 | 10 | 2.6 h |
| Gorilla | UCL-CA4 | S29 | UCL Elliot Smith | LRM3 | 100 | 20 | 10 | 2.0 h |
| Pan | UCL-CA11 | S22 | UCL Elliot Smith | LLM1 | 100 | 15 | 10 | 2.2 h |
| Pan | UCL-CA19B | S31 | UCL Elliot Smith | LLM3 | 100 | 20 | 10 | 1.5 h |
| Pan | UCL-CA198 | S30 | UCL Elliot Smith | LLM1 | 100 | 20 | 10 | 1.4 h |
| Pan | UCL-CA11D | S34 | UCL Elliot Smith | LM3 | 100 | 10 | 10 | 1.3 h |
| Pan | UCL-CA14E | UCL Elliot Smith | UI1 | 100 | 5 | 25 | 2.0 h | |
| Homo sapiens | NCL 2 | S3 | NCL Anat | Udm2 | 120 | 2.5 | 10 | 2.1 h |
| Homo sapiens | modern | S6 | UCL Anat | Ldc | 80–100 | 10 | 10 | 1.3 h |
| Homo sapiens | modern | S7 | UCL Anat | Udm1 | 80–100 | 10 1 | 10 10 | 1.6 h 50 min |
| Homo sapiens | modern | S8 | UCL Anat | Udm2 | 80–100 | 10 | 10 | 2.5 h |
| Homo sapiens | modern | S9 | UCL Anat | Udm2 | 80–100 | 15 | 10 | 1.0 h |
| Homo sapiens | modern | S50 | UCL Anat | LLdc | 80–100 | 1 | 15 | 1.1 h |
| Homo sapiens | modern | S51 | UCL Anat | URdm1 | 80–100 | 1 | 15 | 1.1 h |
| Homo sapiens | modern | S209 | UCL Anat | Udm2 | 80–100 | 2.5 | 4 | 3.2 h |
| Homo sapiens | modern | Tr | UCL Anat | Udm2 | 80–100 | 10 | 10 | 1.0 h |
| Homo sapiens | modern | B40 | UCL Anat | LM3 | 80–100 | 4 | 10 | 1.8 h |
| Homo sapiens | modern | B11 | UCL Anat | UC | 80–100 | 11 | 7 | 4.3 h |
| Homo sapiens | modern | B9CD2 | UCL Anat | UC | 80–100 | 5 3 | 2 8 | 23 min 2.6 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dean, M.C.; Garrevoet, J.; Van Malderen, S.J.M.; Santos, F.; Mirazón Lahr, M.; Foley, R.; Le Cabec, A. The Distribution and Biogenic Origins of Zinc in the Mineralised Tooth Tissues of Modern and Fossil Hominoids: Implications for Life History, Diet and Taphonomy. Biology 2023, 12, 1455. https://doi.org/10.3390/biology12121455
Dean MC, Garrevoet J, Van Malderen SJM, Santos F, Mirazón Lahr M, Foley R, Le Cabec A. The Distribution and Biogenic Origins of Zinc in the Mineralised Tooth Tissues of Modern and Fossil Hominoids: Implications for Life History, Diet and Taphonomy. Biology. 2023; 12(12):1455. https://doi.org/10.3390/biology12121455
Chicago/Turabian StyleDean, M. Christopher, Jan Garrevoet, Stijn J. M. Van Malderen, Frédéric Santos, Marta Mirazón Lahr, Robert Foley, and Adeline Le Cabec. 2023. "The Distribution and Biogenic Origins of Zinc in the Mineralised Tooth Tissues of Modern and Fossil Hominoids: Implications for Life History, Diet and Taphonomy" Biology 12, no. 12: 1455. https://doi.org/10.3390/biology12121455
APA StyleDean, M. C., Garrevoet, J., Van Malderen, S. J. M., Santos, F., Mirazón Lahr, M., Foley, R., & Le Cabec, A. (2023). The Distribution and Biogenic Origins of Zinc in the Mineralised Tooth Tissues of Modern and Fossil Hominoids: Implications for Life History, Diet and Taphonomy. Biology, 12(12), 1455. https://doi.org/10.3390/biology12121455