
Recognition of Rare Microfossils Using Transfer Learning and Deep Residual Networks

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
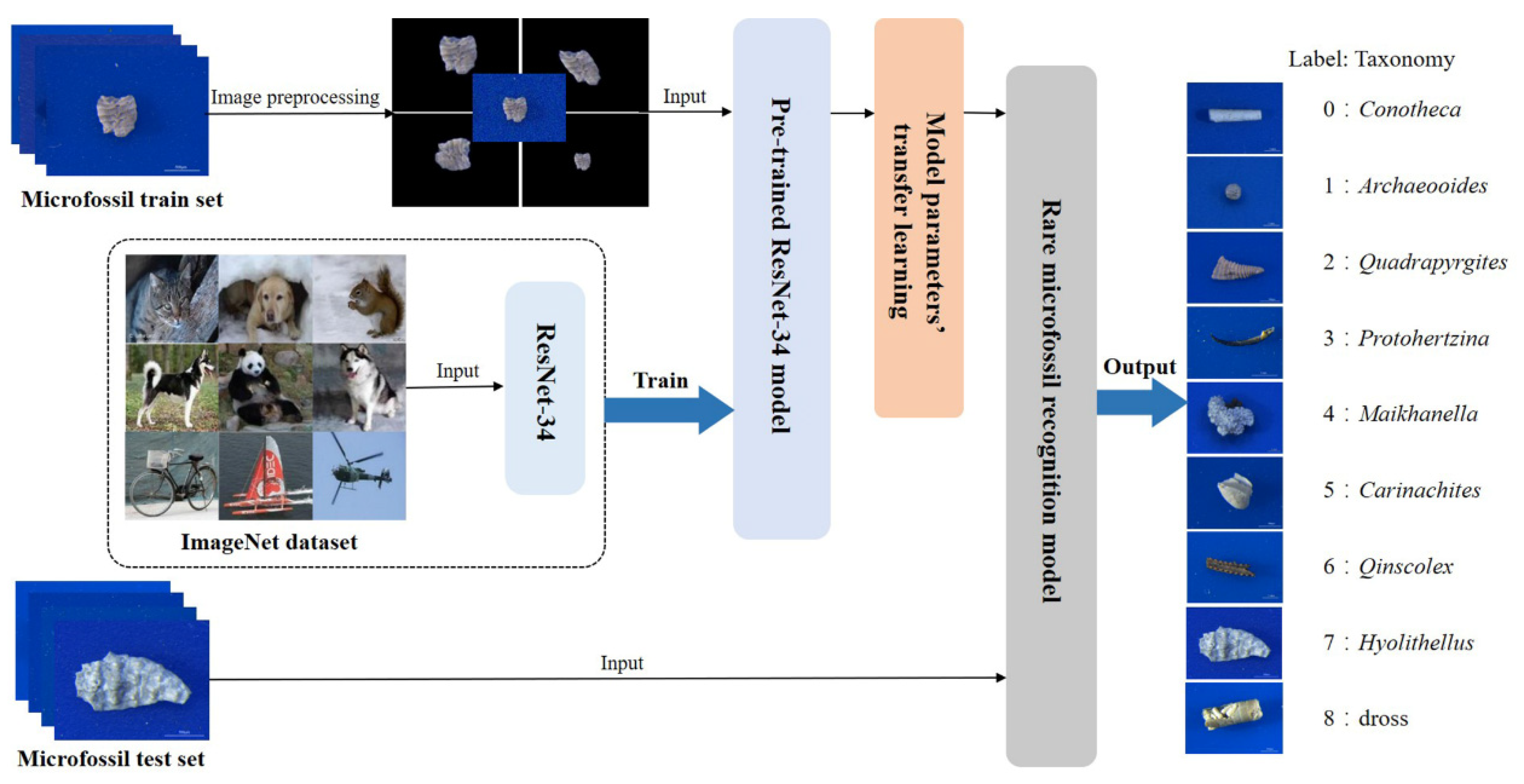
2. Materials and Methods
2.1. Hardware and Software Environment
2.2. Data Acquisition
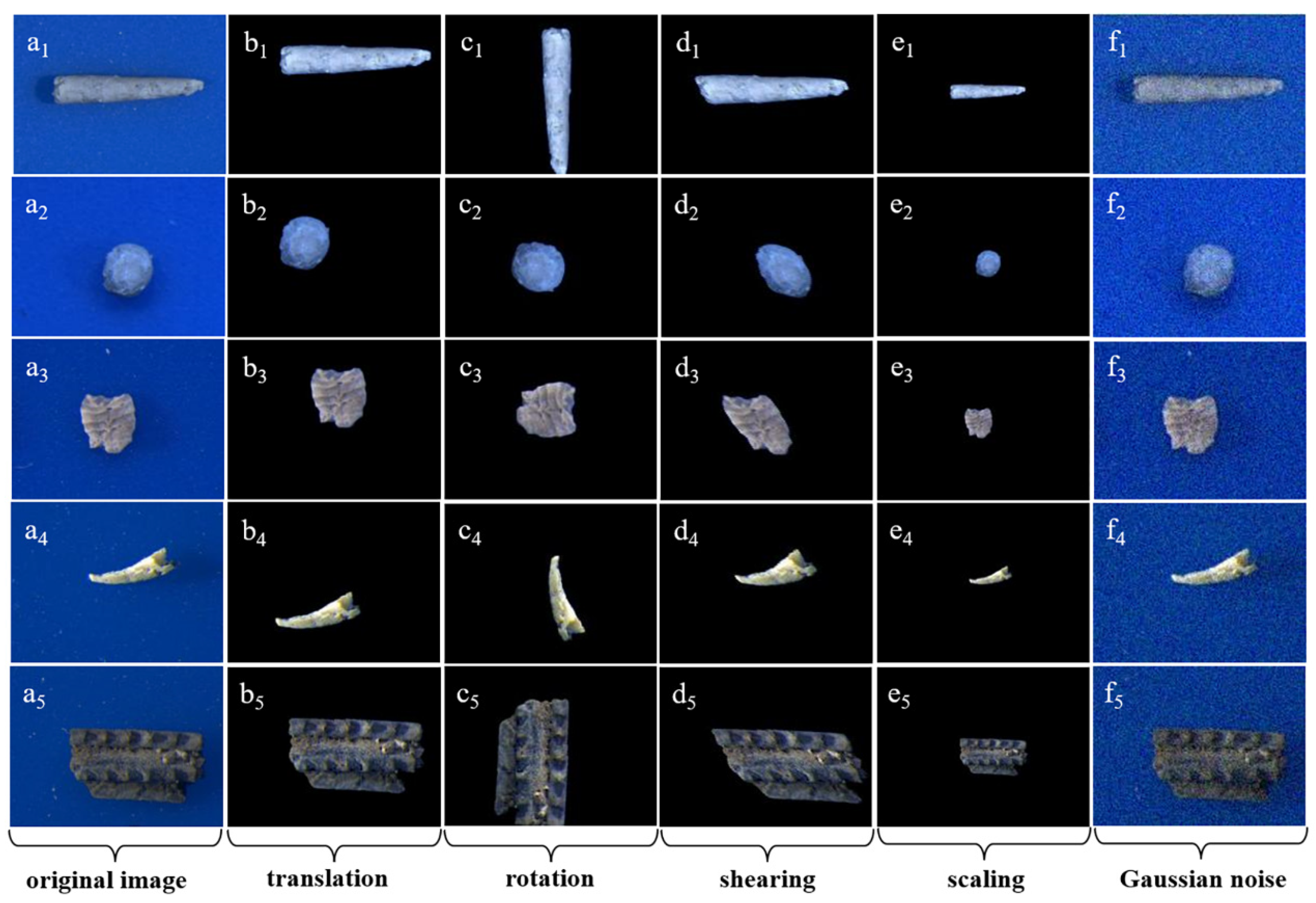
2.3. Data Preprocessing
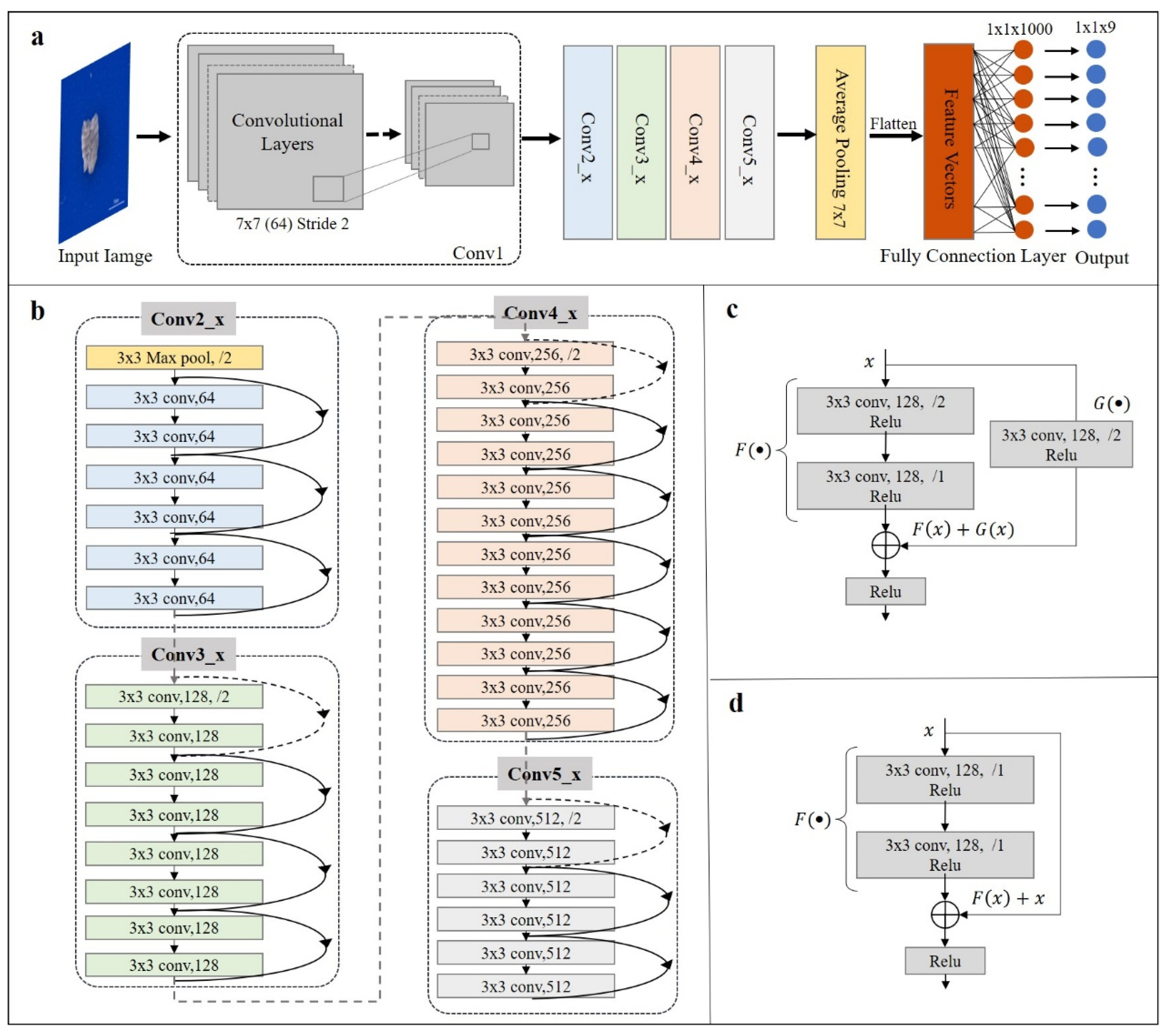
3. Network Structure
- TF-ResNet (ResNet + Train Full Connection Layers): Only the full connection layers are trained and the parameters of the remaining layers of the pre-trained model are retained.
- TS-ResNet (ResNet + Train Specific Characteristic Layers): The feature extraction layer of the pre-trained model is divided into two parts: general feature layers (learning edge, texture and color features) and special feature layers (learning more abstract internal features), keeping the parameters of the bottom general feature layers and retraining the parameters of the top special feature layers.
- TA-ResNet (ResNet + Train All Layers): A pre-trained model structure is used and all the entire network parameters are retrained based on the microfossil dataset.
4. Experiments and Results
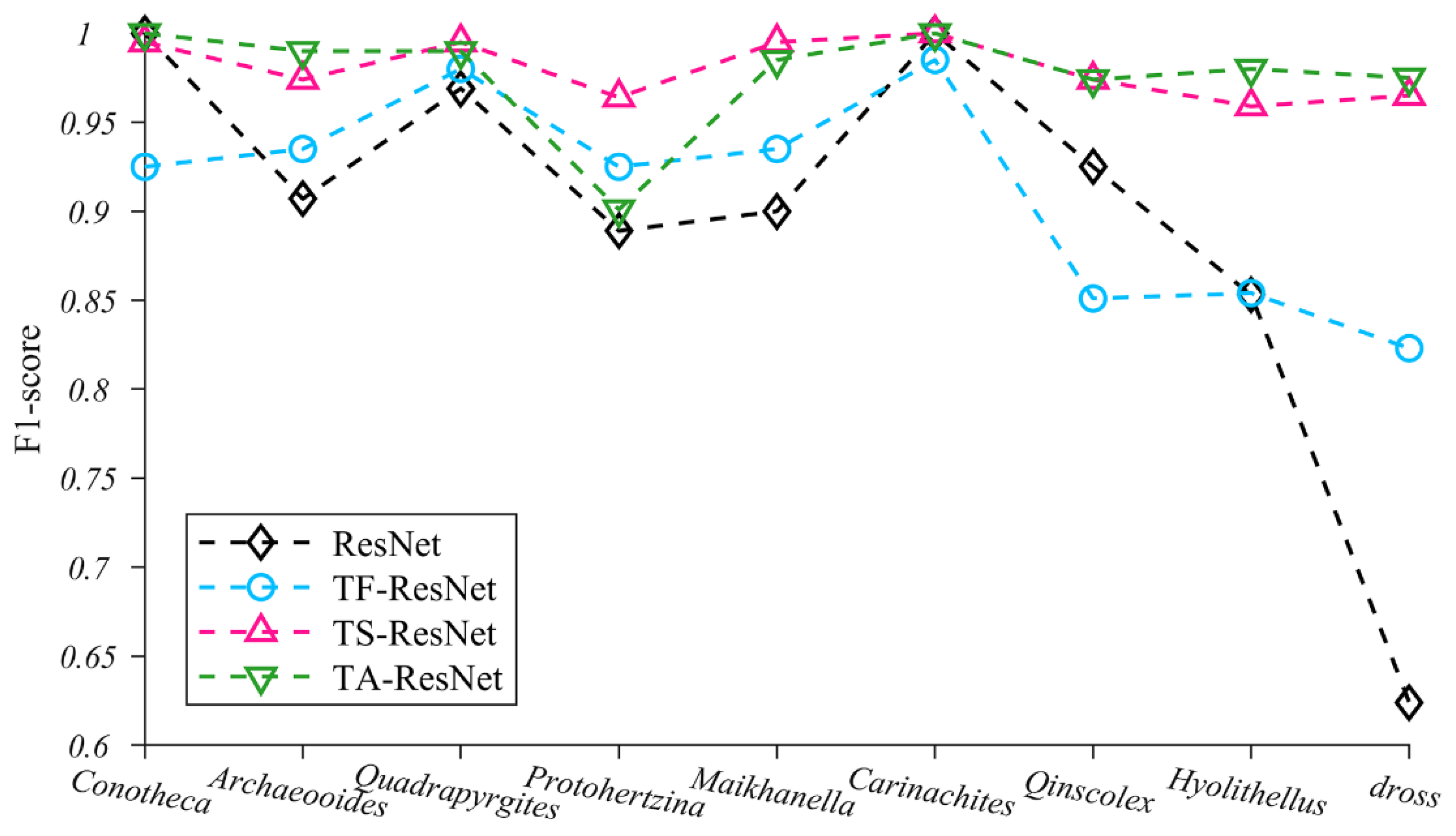
4.1. Evaluation of Transfer Strategies
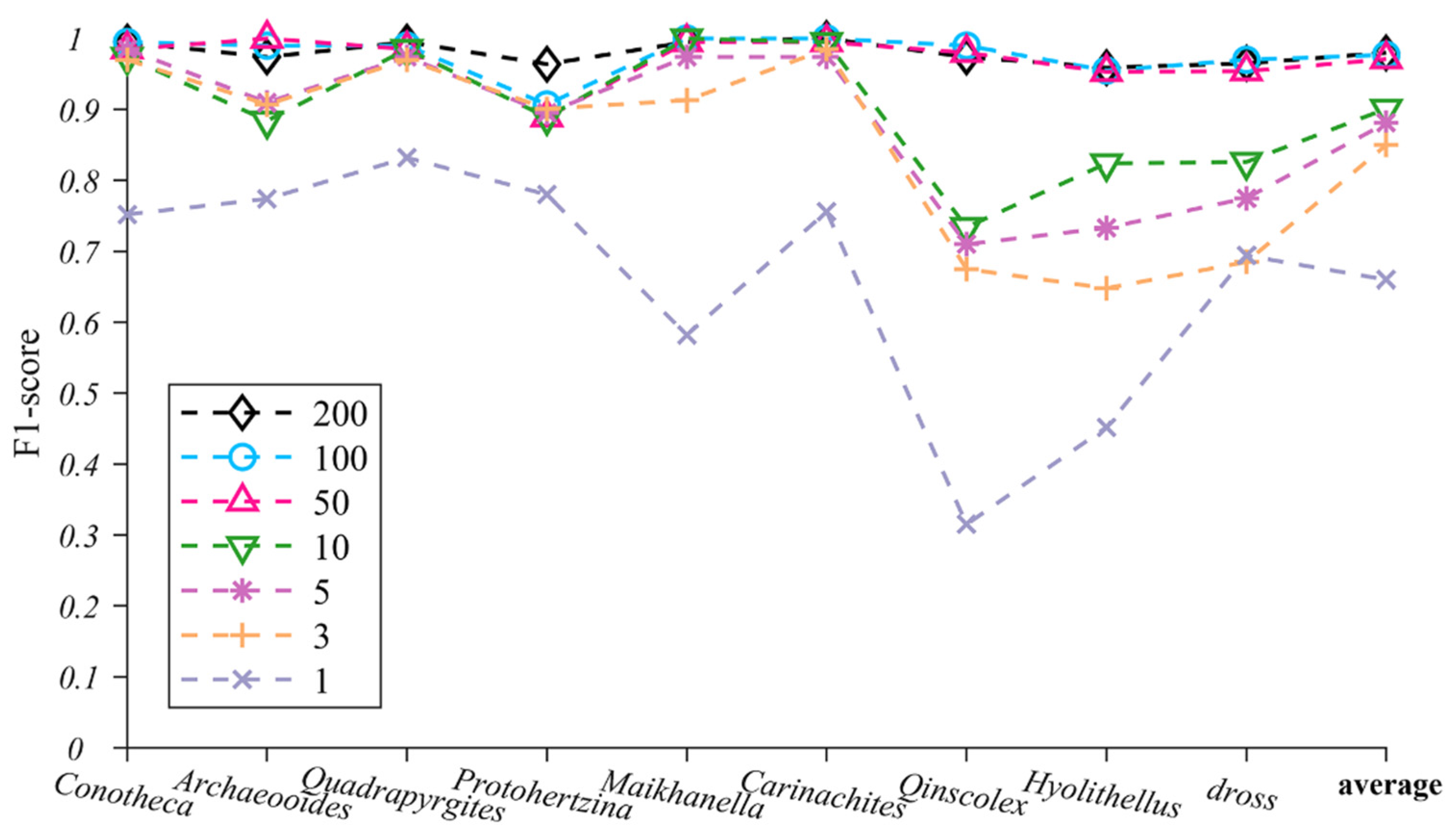
4.2. TS-ResNet Performance on Training Datasets of Varied Sizes
4.3. Performance Comparison of TS-ResNet with Other Models
5. Discussion
5.1. Paleontological Significance of Rare Taxa Recognition
5.2. Necessity and Feasibility of Transferring Pre-Trained Models Based on Natural Images
5.3. Performance Analysis of TS-ResNet
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Budd, G.E.; Jackson, I.S. Ecological innovations in the Cambrian and the origins of the crown group phyla. Philos. T. R. Soc. B 2016, 371, 20150287. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Liu, W.; Zhang, X. Phosphatized microbial fossils from the lowest Cambrian of South China and their ecological and environmental implications for the Kuanchuanpu biota. Precambrian Res. 2018, 338, 105560. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Tang, Q.Q.; Liu, P.; Sun, J.; Yao, X.Y.; Hao, W.J.; Yang, X.G.; Guo, J.F.; Uesugi, K.; Komiya, T.; et al. Characterization of the Multicellular Membrane-Bearing Algae From the Kuanchuanpu Biota (Cambrian: Terreneuvian). J. Geophys. Res. Biogeo. 2021, 126, e2020JG006102. [Google Scholar] [CrossRef]
- Zhang, J.; Hao, X.; Hu, Y.; Yang, X.G.; Wang, X.; Yao, X.Y.; Yang, S.; Han, J. A spherical hat-shaped organism from the lower Cambrian Kuanchuanpu formation. Acta Palaeontol. Sin. 2017, 56, 415–424. (In Chinese) [Google Scholar]
- Dong, X.P.; Cunningham, J.A.; Bengtson, S.; Thomas, C.W.; Liu, J.B.; Stampanoni, M.; Donohue, P.C.J. Embryos, polyps and medusae of the Early Cambrian scyphozoan Olivooides. Proc. Roy. Soc. B-Biol. Sci. 2013, 280, 20130071. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Kubota, S.; Li, G.; Yao, X.; Yang, X.; Shu, D.; Li, Y.; Kinoshita, S.; Sasaki, O.; Komiya, T.; et al. Early Cambrian pentamerous cubozoan embryos from South China. PLoS ONE 2013, 8, e70741. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. The new record of phosphatized Cambricodium fossils from the basal Cambrian of the southern Shaanxi, China. Acta Micropalaeontol. Sin. 2014, 31, 98–103. [Google Scholar]
- Shu, D.; Han, J. Core value of the Chengjiang fauna:formation of the animal kingdom and the birth of basic human organs. Earth Sci. Front. 2020, 27, 31. [Google Scholar]
- Han, J.; Shin, K.; Li, G.X.; Yao, X.Y.; Yang, X.G.; Shu, D.G.; Yong, L.; Kinoshita, S.; Sasaki, O.; Komiya, T.; et al. Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China). Nature 2017, 542, 228–231. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, X.P. The oldest known larva and its implications for the plesiomorphy of metazoan development. Sci. Bull. 2015, 60, 1947–1953. [Google Scholar] [CrossRef] [Green Version]
- Shu, D.G.; Morris, S.C.; Han, J.; Zhang, Z.F.; Liu, J.N. Ancestral echinoderms from the Chengjiang deposits of China. Nature 2004, 430, 422–428. [Google Scholar] [CrossRef]
- Huang, Z.; Bai, Z.Q.; Chai, H. Identification of the ‘Ambiguity Features’ of the Conodont by the Artificial Neural Network. Geol. Sci. Technol. Inf. 2009, 28, 94–98. [Google Scholar]
- Tcheng, D.K.; Nayak, A.K.; Fowlkes, C.C.; Punyasena, S.W.; Peter, W. Visual recognition doftware for binary classification and its application to spruce pollen identification. PLoS ONE 2016, 11, e0148879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, B.; Dedong, L. Artificial Intelligence Identification of Multiple Microfossils from the Cambrian Kuanchuanpu Formation in Southern Shaanxi, China. Acta Geol. Sin-Engl. 2020, 94, 189–197. [Google Scholar] [CrossRef]
- Lecun, Y.; Bottou, L. Gradient-based learning applied to document recognition. Proc. IEEE 1998, 86, 2278–2324. [Google Scholar] [CrossRef] [Green Version]
- Szegedy, C.; Liu, W.; Jia, Y.; Sermanet, P.; Reed, S.; Anguelov, D.; Rabinovich, A.; Erhan, D.; Vanhoucke, V.; Rabinovich, A. Going deeper with convolutions. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Boston, MA, USA, 7–12 June 2015. [Google Scholar]
- Szegedy, C.; Vanhoucke, V.; Ioffe, S.; Shlens, J.; Wojna, Z. Rethinking the inception architecture for computer vision. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016. [Google Scholar]
- Simonyan, K.; Zisserman, A. Very deep convolutional networks for large-scale image recognition. arXiv 2014, arXiv:1409.1556. [Google Scholar]
- Itaki, T.; Taira, Y.; Kuwamori, N.; Saito, H.; Hoshino, T. Innovative microfossil (radiolarian) analysis using a system for automated image collection and AI-based classification of species. Sci. Rep. 2020, 10, 21136. [Google Scholar] [CrossRef]
- Pires de Lima, R.; Welch, K.F.; Barrick, J.E.; Marfurt, K.J.; Burkhalter, R.; Cassel, M.; Soreghan, G.S. Convolutional neural networks as an aid to biostratigraphy and micropaleontology: A test on late paleozoic microfossils. Palaios 2020, 35, 391–402. [Google Scholar] [CrossRef]
- Wang, H.; Li, C.F.; Zhang, Z.F.; Kershaw, S.; Holmer, L.E.; Zhang, Y.; Wei, K.Y.; Liu, P. Fossil Brachiopod identification using a new deep convolutional neural network. Gondwana Res. 2021, 105, 290–298. [Google Scholar] [CrossRef]
- Liu, X.K.; Song, H.J. Automatic identification of fossils and abiotic grains during carbonate microfacies analysis using deep convolutional neural networks. Sediment. Geol. 2020, 410, 105790. [Google Scholar] [CrossRef]
- Hsiang, A.Y.; Brombacher, A.; Rillo, M.C.; Mleneck-Vautravers, M.J.; Conn, S.; Lordsmith, S.; Jenten, A.; Henehan, M.J.; Metcalfe, B.; Fenton, I.S.; et al. Endless forams: > 34,000 modern planktonic foraminiferal images for taxonomic training and automated species recognition using convolutional neural networks. Paleoceanog. Paleocl. 2019, 34, 1157–1177. [Google Scholar] [CrossRef] [Green Version]
- Bourel, B.; Marchant, R.; de Garidel-Thoron, T.; Tetard, M.; Barboni, D.; Gally, Y.; Beaufort, L. Automated recognition by multiple convolutional neural networks of modern, fossil, intact and damaged pollen grains. Comput. Geosci. 2020, 140, 104498. [Google Scholar] [CrossRef]
- He, K.M.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Pan, S.J.; Yang, Q. A Survey on transfer learning. IEEE Trans. Knowl. Data Eng. 2009, 22, 1345–1359. [Google Scholar] [CrossRef]
- Ribani, R.; Marengoni, M. A survey of transfer learning for convolutional neural networks. In Proceedings of the 2019 32nd SIBGRAPI Conference on Graphics, Patterns and Images Tutorials (SIBGRAPI-T), Rio de Janeiro, Brazil, 28–31 October 2019. [Google Scholar]
- Deng, J.; Dong, W.; Socher, R.; Li, L.J.; Li, K.; Fei-Fei, L. ImageNet: A large-scale hierarchical image database. In Proceedings of the 2009 IEEE Conference on Computer Vision and Pattern Recognition, Miami, FL, USA, 20–25 June 2009.
- Steiner, M.; Li, G.X.; Qian, Y.; Zhu, M.Y. Lower Cambrian Small Shelly Fossils of northern Sichuan and southern Shaanxi (China), and their biostratigraphic importance. Geobios 2004, 37, 259–275. [Google Scholar] [CrossRef]
- Liu, Y.H.; Qin, J.C.; Wang, Q.; Maas, A.; Duan, B.; Zhang, Y.N.; Zhang, H.; Shao, T.; Zhang, H.Q. New Armoured Scalidophorans (Ecdysozoa, Cycloneuralia) from the Cambrian Fortunian Zhangjiagou Lagerstatte, South China. Pap. Palaeontol. 2019, 5, 241–260. [Google Scholar] [CrossRef]
- Prasad, S.; Prasad, R. Data standardization and scaling technique for the implementation of human bond communication. Wireless Pers. Commun. 2020, 110, 959–972. [Google Scholar] [CrossRef]
- Wu, Z.; Shen, C.; Hengel, A. Wider or deeper: Revisiting the ResNet model for visual recognition. Pattern Recogn. 2016, 90, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Russakovsky, O.; Deng, J.; Su, H.; Krause, J.; Satheesh, S.; Ma, S.; Huang, Z.; Karpathy, A.; Khosla, A.; Bernstein, M.; et al. Imagenet large scale visual recognition challenge. Int. J. Comput. Vision 2015, 115, 211–252. [Google Scholar] [CrossRef] [Green Version]
- Oquab, M.; Bottou, L.; Laptev, I.; Sivic, J. Learning and transferring mid-level image representations using convolutional neural networks. In Proceedings of the 2014 IEEE Conference on Computer Vision and Pattern Recognition, Columbus, OH, USA, 23–28 June 2014. [Google Scholar]
- Tan, C.; Sun, F.; Kong, T.; Zhang, W.; Yang, C.; Liu, C. A survey on deep transfer learning. arXiv 2018, arXiv:1808.01974. [Google Scholar]
- Goutte, C.; Gaussier, E. A probabilistic interpretation of precision, recall and F-score, with implication for evaluation. Proc. ECIR’05 2005, 3408, 345–359. [Google Scholar]
- Yosinski, J.; Clune, J.; Bengio, Y.; Lipson, H. How transferable are features in deep neural networks? In Proceedings of the Advances in Neural Information Processing Systems 27, Montréal, QC, Canada, 8–13 December 2014. [Google Scholar]
- Bengtson, S. Early Cambrian shelly fossils from South Australia. Australas. Assoc. Palaeontol. Mem 1990, 9, 190–216. [Google Scholar]
- Kouchinsky; Bengtson, S.; Feng, W.; Kutygin, R.; Val’kov, A. The Lower Cambrian fossil Anabaritids: Affinities, occurrences and systematics. J. Syst. Palaeontol. 2009, 7, 241–298. [Google Scholar] [CrossRef]
- Hua, H.; Chen, Z.; Yuan, X.; Zhang, L.; Xiao, S. Skeletogenesis and asexual reproduction in the earliest biomineralizing animal Cloudina. Geology 2005, 33, 277–280. [Google Scholar] [CrossRef]
- Han, J.; Cai, Y.; Schiffbauer, J.D.; Hua, H.; Wang, X.; Yang, X.; Uesugi, K.; Komiya, T.; Sun, J. A Cloudina-like fossil with evidence of asexual reproduction from the lowest Cambrian, South China. Geol. Mag. 2017, 154, 1294–1305. [Google Scholar] [CrossRef]
- Erwin, D.H.; Laflamme, M.; Tweedt, S.M.; Sperling, E.A.; Pisani, D.; Peterson, K.J. The Cambrian conundrum: Early divergence and later ecological success in the early history of animals. Science 2011, 334, 1091–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, K.J.; Cotton, J.A.; Gehling, J.G.; Pisani, D. The Ediacaran emergence of bilaterians: Congruence between the genetic and the geological fossil records. Philos. Trans. R Soc. Lond. B Biol. Sci. 2008, 363, 1435–1443. [Google Scholar] [CrossRef]
- Murdock, D.J.E. The ‘biomineralization toolkit’ and the origin of animal skeletons. Biol. Rev. 2020, 95, 1372–1392. [Google Scholar] [CrossRef]
- Budd, G.E. The Cambrian fossil record and the origin of the Phyla. Integr. Comp. Biol. 2003, 43, 157–165. [Google Scholar] [CrossRef]
- Wang, D.; Vannier, J.; Schumann, I.; Wang, X.; Yang, X.G.; Komiya, T.; Uesugi, K.; Sun, J.; Han, J. Origin of ecdysis: Fossil evidence from 535-million-year-old scalidophoran worms. Proc. Roy. Soc. B-Biol. Sci. 2019, 286, 20190791. [Google Scholar] [CrossRef] [Green Version]
- Butterfield, N.J. Exceptional fossil preservation and the Cambrian explosion. Integr. Comp. Biol. 2003, 43, 166–177. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomy | Shape | Size | Total Amounts |
|---|---|---|---|
| ? Cnidaria: Conotheca | cone-shaped tube | 3 mm (±2 mm) | 858 |
| Animals: Archaeooides | sphere | 1.5 mm (±1 mm) | 968 |
| Cnidaria: Quadrapyrgites | cone-shaped tower | 1 mm (±0.75 mm) | 357 |
| Chaetognatha: Protohertzina | curved spines | 1.5 mm (±1 mm) | 905 |
| Mollusca: Maikhanella | shell | 0.6 mm (±0.2 mm) | 210 |
| Cnidaria: Carinachites | tube with four slots | 3 mm (±2 mm) | 138 |
| Scalidophora: Qinscolex | worms | 1 mm (±0.5 mm) | 56 |
| ? Annelida: Hyolithellus | straight, segmented tube | 1.5 mm (±1 mm) | 83 |
| null | Irregular-shaped dross | 3 mm (±2 mm) | 932 |
| Strategies | Conv1 | Conv2_x | Conv3_x | Conv4_x | Conv5_x | Fc (Fully Connected Layer) |
|---|---|---|---|---|---|---|
| TF-ResNet | ✕ | ✕ | ✕ | ✕ | ✕ | ✓ |
| TS-ResNet | ✕ | ✕ | ✕ | ✕ | ✓ | ✓ |
| ✕ | ✕ | ✕ | ✓ | ✓ | ✓ | |
| ✕ | ✕ | ✓ | ✓ | ✓ | ✓ | |
| ✕ | ✓ | ✓ | ✓ | ✓ | ✓ | |
| TA-ResNet | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Confusion Matrix | True Labels | Evaluation | ||
|---|---|---|---|---|
| Positive | Negative | |||
| Predicted | Positive | True Positive | False Positive | |
| Negative | False Negative | True Negative | ||
| Evaluation | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Sun, R.; Yang, X.; Niu, B.; Zhang, T.; Zhao, Y.; Zhang, Y.; Zhang, Y.; Han, J. Recognition of Rare Microfossils Using Transfer Learning and Deep Residual Networks. Biology 2023, 12, 16. https://doi.org/10.3390/biology12010016
Wang B, Sun R, Yang X, Niu B, Zhang T, Zhao Y, Zhang Y, Zhang Y, Han J. Recognition of Rare Microfossils Using Transfer Learning and Deep Residual Networks. Biology. 2023; 12(1):16. https://doi.org/10.3390/biology12010016
Chicago/Turabian StyleWang, Bin, Ruyue Sun, Xiaoguang Yang, Ben Niu, Tao Zhang, Yuandi Zhao, Yuanhui Zhang, Yiheng Zhang, and Jian Han. 2023. "Recognition of Rare Microfossils Using Transfer Learning and Deep Residual Networks" Biology 12, no. 1: 16. https://doi.org/10.3390/biology12010016
APA StyleWang, B., Sun, R., Yang, X., Niu, B., Zhang, T., Zhao, Y., Zhang, Y., Zhang, Y., & Han, J. (2023). Recognition of Rare Microfossils Using Transfer Learning and Deep Residual Networks. Biology, 12(1), 16. https://doi.org/10.3390/biology12010016