
Copepods as Indicators of Different Water Masses during the Northeast Monsoon Prevailing Period in the Northeast Taiwan

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
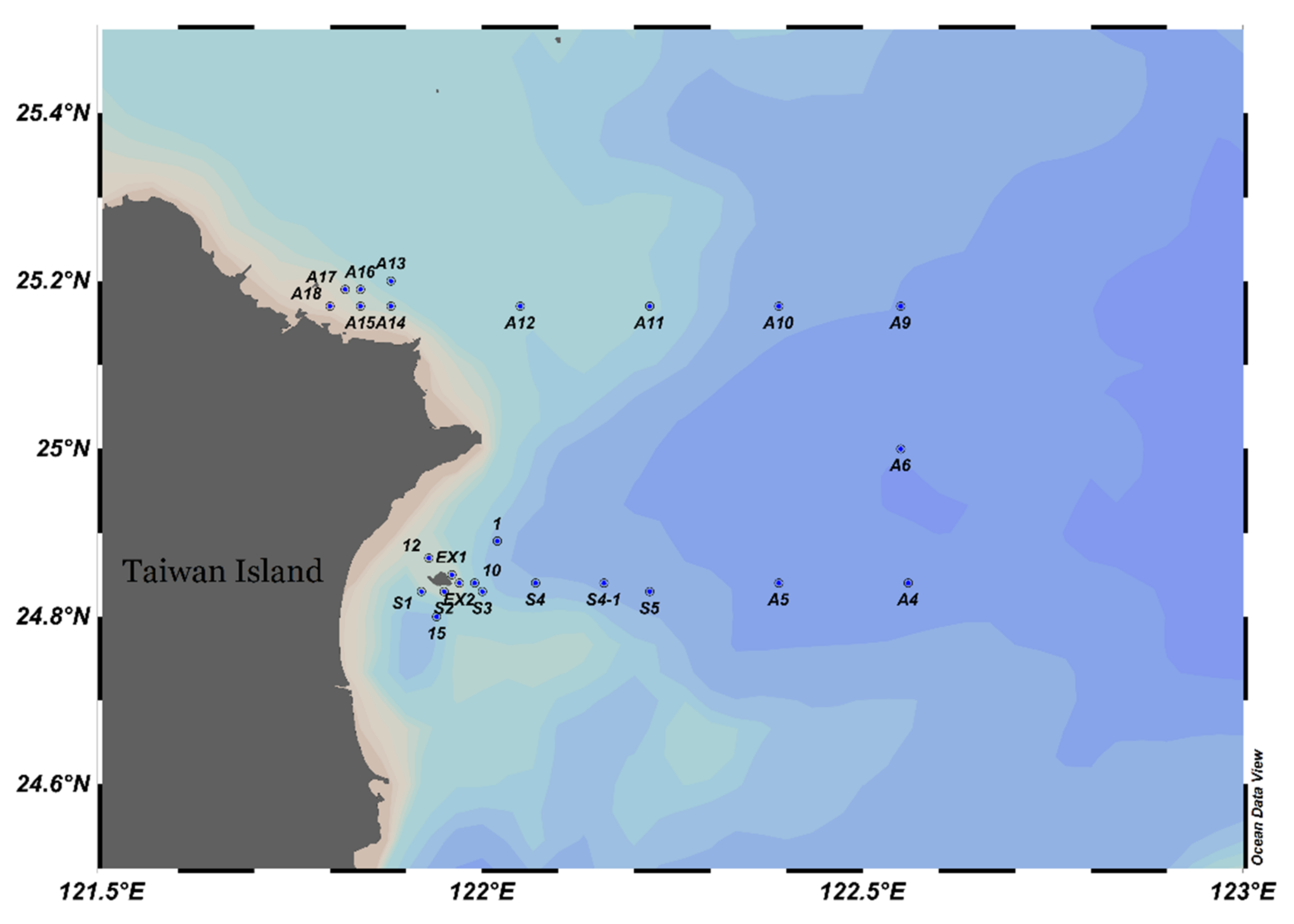
2. Materials and Methods
3. Results
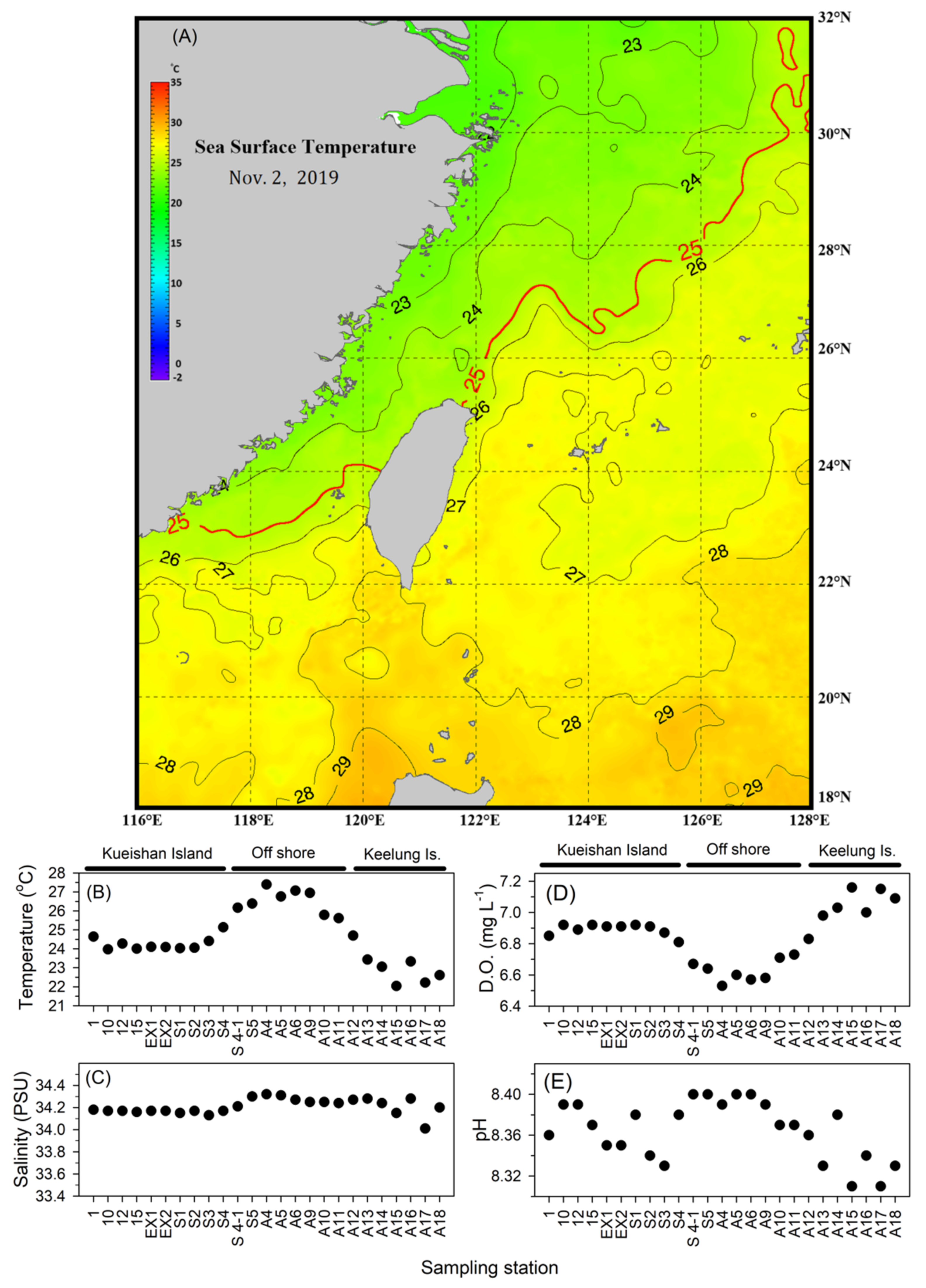
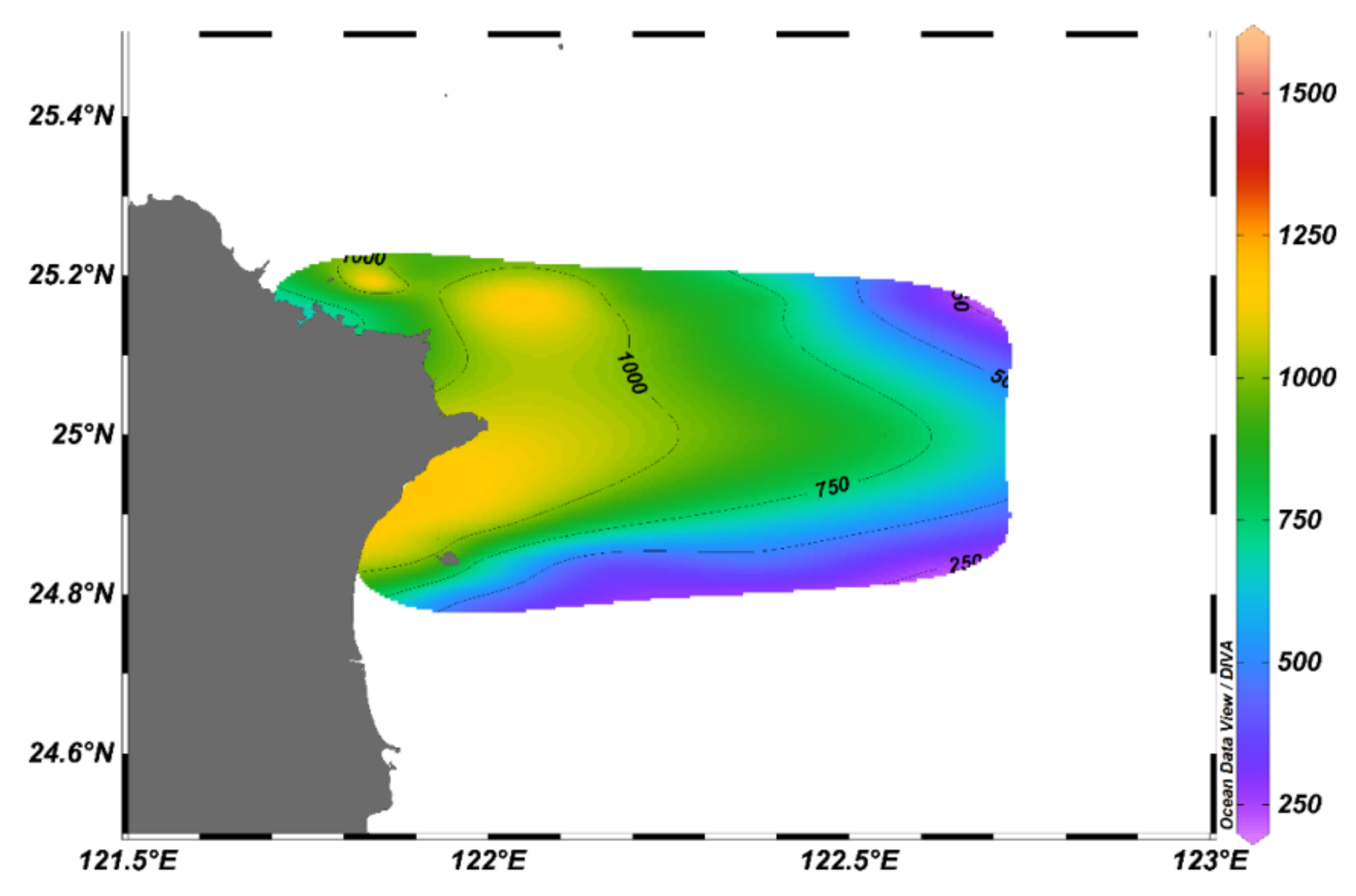
3.1. Spatial Changes in Hydrographic Conditions
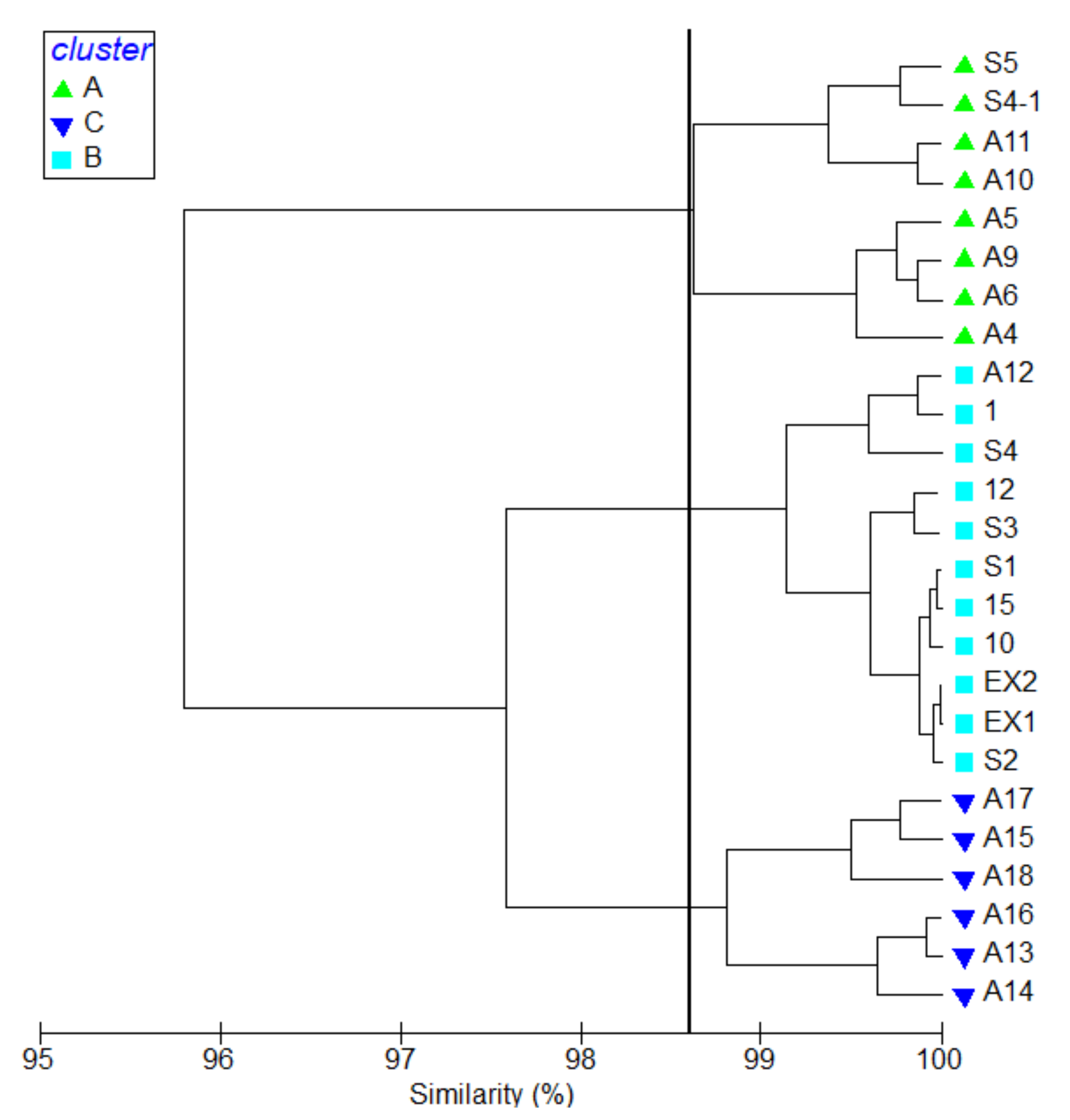
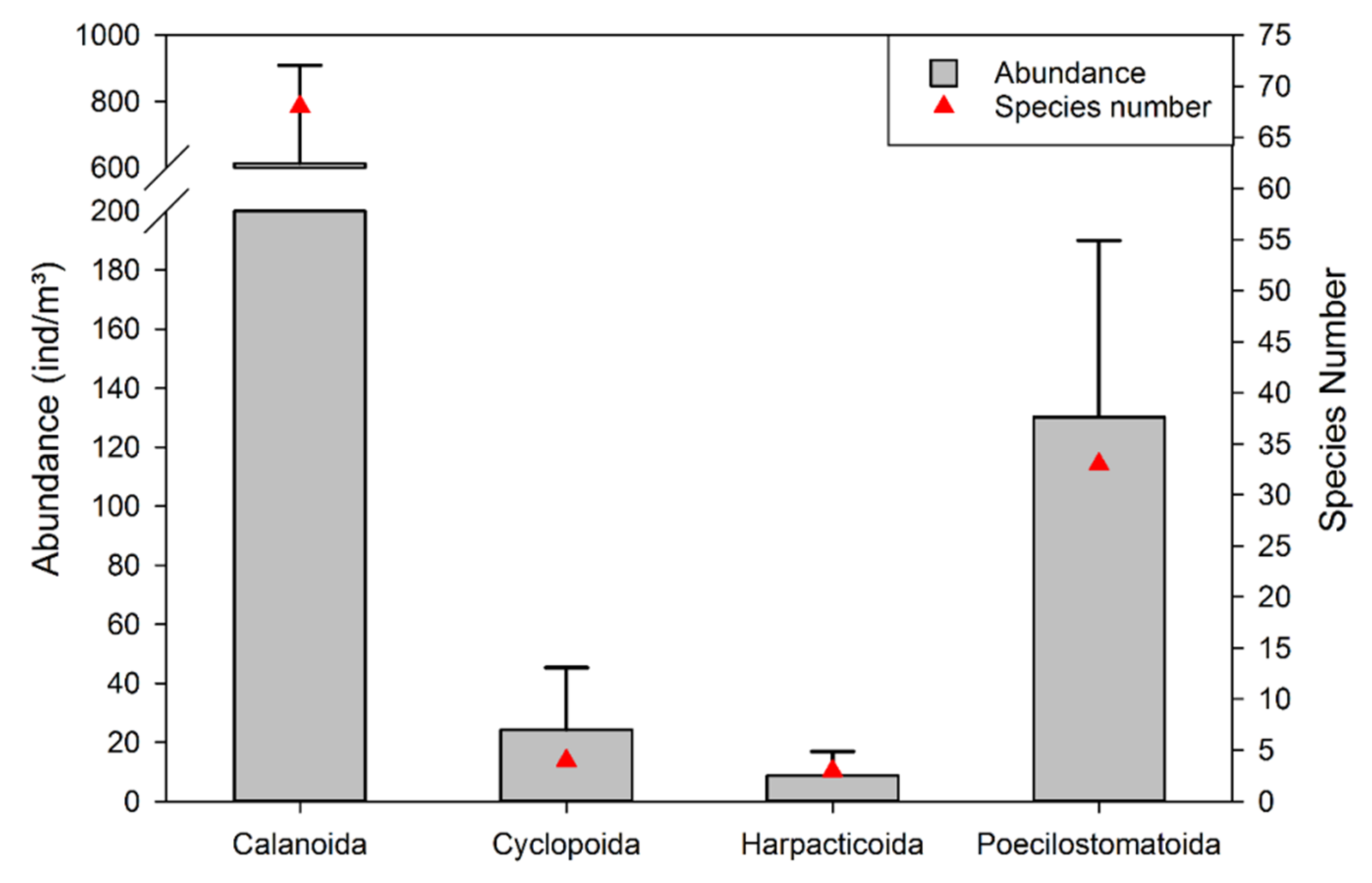
3.2. Species Composition and Spatial Characteristics of Copepod Communities
3.3. Dominant Species Composition and Community Indicator Species
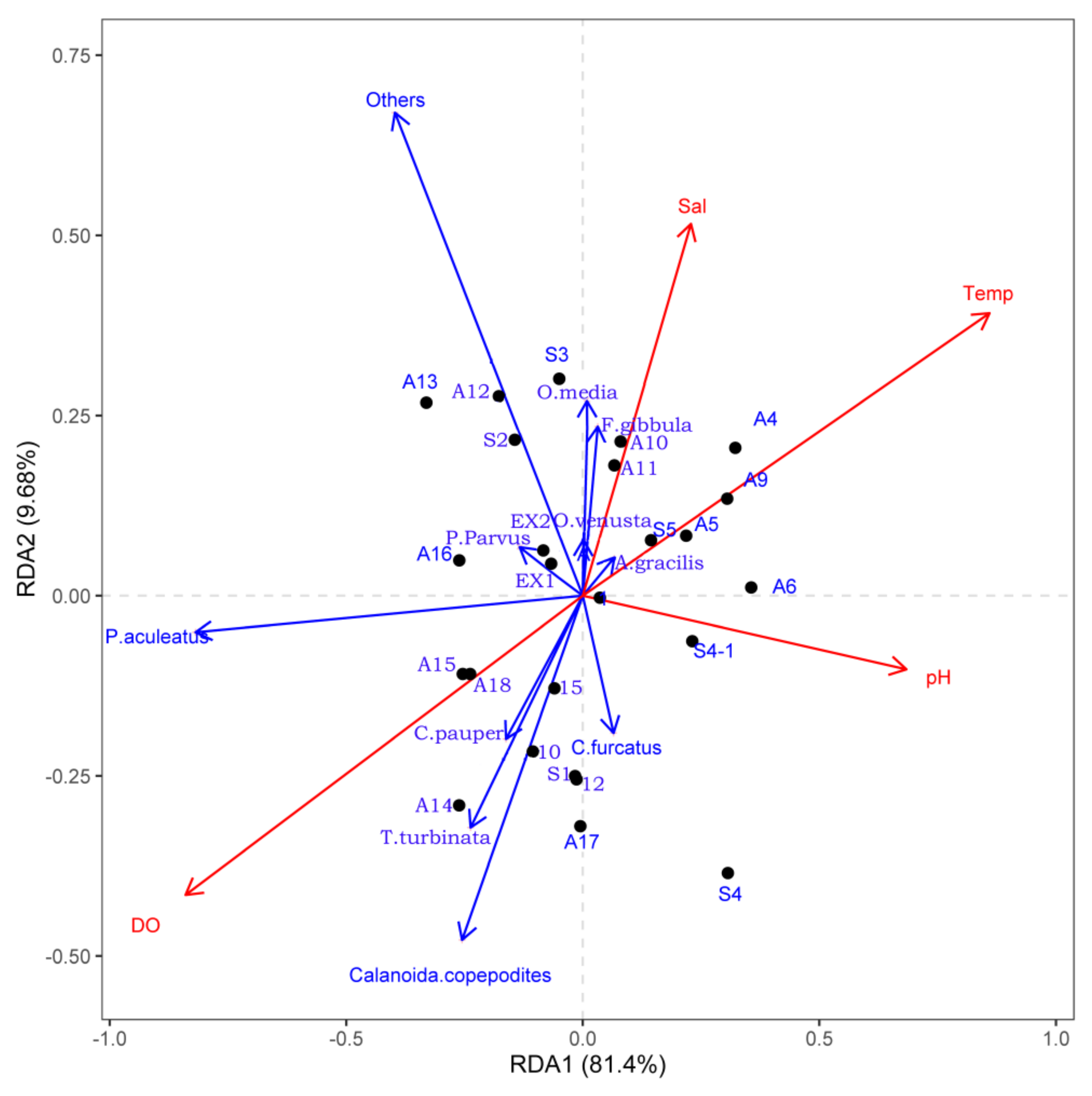
3.4. RDA Analysis of Dominant Species with Environmental Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uriarte, I.; Villate, F. Effects of pollution on zooplankton abundance and distribution in two estuaries of the Basque coast (Bay of Biscay). Mar. Pollut. Bull. 2004, 49, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Hafferssas, A.; Seridji, R. Relationships between the hydrodynamics and changes in copepod structure on the Algerian coast. Zool. Stud. 2010, 49, 353–366. [Google Scholar]
- Drira, Z.; Kmiha-Megdiche, S.; Sahnoun, H.; Pagano, M.; Tedetti, M.; Ayadi, H. Water quality affects the structure of copepod assemblages in the Sfax Southern coast (Tunisia, Southern Mediterranean Sea). Mar. Freshw. Res. 2017, 69, 220–231. [Google Scholar] [CrossRef]
- Drira, Z.; Kmiha-Megdiche, S.; Sahnoun, H.; Tedetti, M.; Pagano, M.; Ayadi, H. Copepod assemblages as a bioindicator of environmental quality in three coastal areas under contrasted anthropogenic inputs (Gulf of Gabes, Tunisia). J. Mar. Biol. Assoc. UK 2017, 98, 1–17. [Google Scholar] [CrossRef]
- Yoshida, T.; Liong, C.; Majid, A.M.; Toda, T.; Ross, B.H. Temperature effects on the egg development time and hatching success of three Acartia species (Copepoda: Calanoida) from the Strait of Malacca Teruaki. Zool. Stud. 2012, 51, 644–654. [Google Scholar]
- Sommer, U.; Paul, C.; Moustaka, G. Warming and ocean acidification effects on phytoplankton from species shifts to size shifts within species in a mesocosm experiment. PLoS ONE 2015, 10, 17. [Google Scholar]
- Bhattacharya, B.D.; Bhattacharya, A.; Rakshit, D.; Sarkar, S.K. Impact of the tropical cyclonic storm ‘Aila’ on the water quality characteristics and mesozooplankton community structure of Sundarban mangrove wetland. Indian J. Geo-Mar. Sci. 2014, 43, 216–223. [Google Scholar]
- Zheng, Z. Marine Planktology; China Ocean Press: Beijing, China, 1989. [Google Scholar]
- Uye, S.; Nagano, N.; Tamaki, T. Geographical and seasonal variations in abundance, biomass and estimated production rates of microzooplankton in the Inland Sea of Japan. J. Oceanogr. 1996, 52, 689–703. [Google Scholar] [CrossRef]
- Tseng, L.C.; Hung, J.J.; Chen, Q.C.; Hwang, J.S. Seasonality of the copepod assemblages associated with interplay waters off northeastern Taiwan. Helgol. Mar. Res. 2013, 67, 507–520. [Google Scholar] [CrossRef]
- Longhurst, A.R. Relationship between diversity and the vertical structure of the upper ocean. Deep. Sea Res. Part A Oceanogr. Res. Pap. 1985, 32, 1535–1570. [Google Scholar] [CrossRef]
- Kâ, S.; Hwang, J.S. Mesozooplankton distribution and composition on the northeastern coast of Taiwan during autumn: Effects of the Kuroshio Current and hydrothermal vents. Zool. Stud. 2011, 50, 155–163. [Google Scholar]
- Yasuhara, M.; Hunt, G.; Dowsett, H.J.; Robinson, M.M.; Stoll, D.K. Latitudinal species diversity gradient of marine zooplankton for the last three million years. Ecol. Lett. 2012, 15, 1174–1179. [Google Scholar] [CrossRef]
- Pepin, P.; Johnson, C.L.; Harvey, M.; Casault, B.; Joel, C.; Colbourne, E.B.; Galbraith, P.S.; Hebert, D.; Lazin, G.; Maillet, G. A multivariate evaluation of environmental effects on zooplankton community structure in the western North Atlantic. Prog. Oceanogr. 2015, 134, 197–220. [Google Scholar] [CrossRef]
- Erondu, C.J.; Solomon, R.J. Identification of planktons (zooplankton and phytoplankton) behind Girls’ Hostel University of Abuja, Nigeria. Direct Res. J. Public Health Environ. Technol. 2017, 2, 21–29. [Google Scholar]
- Nakajima, R.; Yamazaki, H.; Lewis, L.S.; Khen, A.; Smith, J.E.; Nakatomi, N.; Kurihara, H. Planktonic trophic structure in a coral reef ecosystem—Grazing versus microbial food webs and the production of mesozooplankton. Prog. Oceanogr. 2017, 156, 104–120. [Google Scholar] [CrossRef]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 3, 279–295. [Google Scholar] [CrossRef]
- Hwang, J.S.; Souissi, S.; Dahms, H.U.; Tseng, L.C.; Schmitt, F.G.; Chen, Q.C. Rank-abundance allocations as a tool to analyze planktonic copepod assemblages off the Danshuei estuary (Taiwan). Zool. Stud. 2009, 48, 49–62. [Google Scholar]
- Dam, H.G. Evolutionary adaptation of marine zooplankton to global change. Annu. Rev. Mar. Sci. 2013, 5, 349–370. [Google Scholar] [CrossRef]
- Wang, Y.G.; Tseng, L.C.; Lin, M.; Hwang, J.S. Vertical and geographic distribution of copepod communities at late summer in the Amerasian Basin, Arctic Ocean. PLoS ONE 2019, 14, e0219319. [Google Scholar] [CrossRef]
- Kusk, K.O.; Wollenberger, L. Towards an internationally harmonized test method for reproductive and developmental effects of endocrine disrupters in marine copepods. Ecotoxicology 2007, 16, 183–195. [Google Scholar] [CrossRef]
- Raisuddin, S.; Kwok, K.W.H.; Leung, K.M.Y.; Schlenk, D.; Lee, J.S. The copepod Tigriopus: A promising marine model organism for ecotoxicology and environmental genomics. Aquat. Toxicol. 2007, 83, 161–173. [Google Scholar] [CrossRef]
- Daufresne, M.; Lengfellner, K.; Sommer, U. Global warming benefits the small in aquatic ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12788–12793. [Google Scholar] [CrossRef]
- Murphy, E.J.; Hofmann, E.E.; Watkins, J.L.; Johnston, N.M.; Piñones, A.; Ballerini, T.; Hill, S.L.; Trathan, P.N.; Tarling, G.A.; Cavanagh, R.A. Comparison of the structure and function of Southern Ocean regional ecosystems: The Antarctic Peninsula and South Georgia. J. Mar. Syst. 2013, 109–110, 22–42. [Google Scholar] [CrossRef]
- Darnis, G.; Fortier, L. Temperature, food and the seasonal vertical migration of key arctic copepods in the thermally stratified Amundsen Gulf (Beaufort Sea, Arctic Ocean). J. Plankton Res. 2014, 36, 1092–1108. [Google Scholar] [CrossRef]
- Bedford, J.; Ostle, C.; Johns, D.G.; Atkinson, A.; Best, M.; Bresnan, E.; Machairopoulou, M.; Graves, C.A.; Devlin, M.; Milligan, A.; et al. Lifeform indicators reveal large-scale shifts in plankton across the North-West European shelf. Glob. Change Biol. 2020, 26, 3482–3497. [Google Scholar] [CrossRef]
- Morse, R.E.; Friedland, K.D.; Tommasi, D.; Stock, C.; Nye, J. Distinct zooplankton regime shift patterns across ecoregions of the US Northeast Continental shelf Large Marine Ecosystem. J. Mar. Syst. 2017, 165, 77–91. [Google Scholar] [CrossRef]
- Ivory, J.A.; Steinberg, D.K.; Latour, R.J. Diel, seasonal, and interannual patterns in mesozooplankton abundance in the Sargasso Sea. ICES J. Mar. Sci. 2019, 76, 217–231. [Google Scholar] [CrossRef]
- Bode, A.; Álvarez, M.; García, L.M.; Louro, M.A.; Nieto-Cid, M.; Ruíz-Villarreal, M.; Varela, M.M. Climate and local hydrography underlie recent regime shifts in plankton communities off Galicia (NW Spain). Oceans 2020, 1, 181–197. [Google Scholar] [CrossRef]
- Villarino, E.; Irigoien, X.; Villate, F.; Iriarte, A.; Uriarte, I.; Zervoudaki, S.; Carstensen, J.; O’Brien, T.D.; Chust, G. Response of copepod communities to ocean warming in three time-series across the North Atlantic and Mediterranean Sea. Mar. Ecol. Prog. Ser. 2020, 636, 47–61. [Google Scholar] [CrossRef]
- Iriarte, A.; Villate, F.; Uriarte, I.; Bidegain, G.; Barroeta, Z. Shifts in neritic copepod communities off the Basque coast (Southeastern Bay of Biscay) between 1998 and 2005. ICES J. Mar. Sci. 2022, 79, 830–843. [Google Scholar] [CrossRef]
- Liu, H.; Bi, H.; Peterson, W.T. Large-scale forcing of environmental conditions on subarctic copepods in the northern California Current system. Prog. Oceanogr. 2015, 134, 404–412. [Google Scholar] [CrossRef]
- Liu, X.H.; Wang, D.P.; Su, J.; Chen, D.K.; Lian, T.; Dong, C.M.; Liu, T.Y. On the vorticity balance over steep sloops: Kuroshio intrusions Northeast of Taiwan. J. Phys. Oceanogr. 2020, 50, 2089–2104. [Google Scholar] [CrossRef]
- Hwang, J.S.; Wong, C.K. The China Coastal Current as a driving force for transporting Calanus sinicus (Copepoda: Calanoida) from its population centers to waters off Taiwan and Hong Kong during the winter northeast monsoon period. J. Plankton Res. 2005, 27, 205–210. [Google Scholar] [CrossRef]
- Tseng, L.C.; Kumar, R.; Dahms, H.U.; Chen, Q.C.; Hwang, J.S. Monsoon-driven succession of copepod assemblages in the coastal waters of the Northeastern Taiwan Strait. Zool. Stud. 2008, 47, 46–60. [Google Scholar]
- Bi, H.S.; Peterson, W.T.; Strub, P.T. Transport and coastal zooplankton communities in the northern California Current system. Geophys. Res. Lett. 2011, 38, L12607. [Google Scholar] [CrossRef]
- Liu, H.; Peterson, W.T. Seasonal and inter-annual variations in the abundance and biomass of Neocalanus plumchrus in continental slope waters off Oregon. Fish. Oceanogr. 2010, 19, 354–369. [Google Scholar] [CrossRef]
- Tang, T.Y.; Tai, J.H.; Yang, Y.J. The flow pattern north of Taiwan and the migration of the Kuroshio. Cont. Shelf Res. 2000, 20, 349–371. [Google Scholar] [CrossRef]
- Isobe, A. Recent advances in ocean-circulation research on the Yellow Sea and East China Sea shelves. J. Oceanogr. 2008, 64, 569–584. [Google Scholar] [CrossRef]
- Oey, L.Y.; Hsin, Y.C.; Wu, C.R. Why does the Kuroshio northeast of Taiwan shift shelfward in winter? Ocean Dynam. 2010, 60, 413–426. [Google Scholar] [CrossRef]
- Hsin, Y.C.; Qiu, B.; Chiang, T.L.; Wu, C.R. Seasonal to interannual variations in the intensity and central position of the surface Kuroshio east of Taiwan. J. Geophys. Res. Oceans 2013, 118, 4305–4316. [Google Scholar] [CrossRef]
- Yin, Y.; Lin, X.; Li, Y.; Zeng, X. Seasonal variability of Kuroshio intrusion northeast of Taiwan Island as revealed by self-organizing map. Chin. J. Oceanol. Limnol. 2014, 32, 1435–1442. [Google Scholar] [CrossRef]
- Wu, C.R.; Wang, Y.L.; Lin, Y.F.; Chao, S.Y. Intrusion of the Kuroshio into the South and East China Seas. Sci. Rep. 2017, 7, 7895. [Google Scholar] [CrossRef]
- Yin, W.B.; Huang, D.J. Short-term variations in the surface upwelling off northeastern Taiwan observed via satellite data. J. Geophys. Res. Oceans 2019, 124, 939–954. [Google Scholar] [CrossRef]
- Yin, Y.Q.; Liu, Z.; Hu, P.; Hou, Y.J.; Lu, J.Y.; He, Y.S. Impact of mesoscale eddies on the southwestward countercurrent northeast of Taiwan revealed by ADCP mooring observations. Cont. Shelf. Res. 2020, 195, 104063. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Yu, F.; Si, G.C.; Wang, J.F. Characteristics and maintenance mechanisms of the cold eddy to the Northeast of Diaoyu Island. Reg. Stud. Mar. Sci. 2020, 36, 101259. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, H.Y.; Jan, S.; Tuo, S.H. Phytoplankton productivity enhancement and assemblage change in the upstream Kuroshio after typhoon. Mar. Ecol. Prog. Ser. 2009, 385, 111–126. [Google Scholar] [CrossRef]
- Hwang, J.S.; Souissi, S.; Tseng, L.J.; Seuront, L.; Schmitt, F.G.; Fang, L.S.; Peng, S.H.; Wu, C.H.; Hsiao, S.H.; Twan, W.H.; et al. A 5-year study for the influence of the northeast and southwest monsoons on copepod assemblages in the boundary coastal waters between the East China Sea and the Taiwan Strait. J. Plankton Res. 2006, 28, 943–958. [Google Scholar] [CrossRef]
- Hwang, J.S.; Kumar, R.; Hsieh, C.W.; Kuo, A.Y.; Souissi, S.; Hsu, M.H.; Wu, J.T.; Liu, W.; Wang, C.F.; Chen, Q.C. Patterns of zooplankton distribution along the marine, estuarine and riverine portions of the Danshuei ecosystem in northern Taiwan. Zool. Stud. 2010, 49, 335–352. [Google Scholar]
- Dur, G.; Hwang, J.S.; Souissi, S.; Tseng, L.C.; Wu, C.H.; Hsiao, S.H.; Chen, Q.C. An overview of the influence of hydrodynamics on the spatial and temporal patterns of calanoid copepod communities around Taiwan. J. Plankton Res. 2007, 29 (Suppl. S1), 97–116. [Google Scholar] [CrossRef]
- Tseng, L.C.; Kumar, R.; Chen, Q.C.; Hwang, J.S. Faunal shift between two copepod congeners (Temora discaudata and T. turbinata) in the vicinity of two nuclear power plants in southern East China Sea: Spatiotemporal patterns of population trajectories over a decade. Hydrobiologia 2011, 666, 301–315. [Google Scholar] [CrossRef]
- Hsieh, H.; Lo, W.Z. Mesoscale distribution and assemblage structure of fish larvae in the Kuroshio waters off eastern Taiwan. Mar. Biodiv. 2019, 49, 1971–1986. [Google Scholar] [CrossRef]
- Shih, C.T.; Chiu, T.S. Copepod diversity in the water masses of the southern East China Sea north of Taiwan. J. Mar. Syst. 1998, 15, 533–542. [Google Scholar] [CrossRef]
- Shih, C.T.; Hwang, J.S.; Huang, W.B. Planktonic copepods from an upwelling station north of Taiwan, western North Pacific. J. Taiwan Mus. 2000, 10, 19–35. [Google Scholar]
- Lee, C.Y.; Liu, D.C.; Su, W.C. Seasonal and spatial variations in the planktonic copepod community of I-Lan Bay and adjacent Kuroshio waters off Northeast Taiwan. Zool. Stud. 2009, 48, 151–161. [Google Scholar]
- Wang, Y.G.; Tseng, L.C.; Xing, B.P.; Sun, R.X.; Chen, X.Y.; Wang, C.G.; Hwang, J.S. Seasonal population structure of the copepod Temora turbinata (Dana, 1849) in the Kuroshio Current edge, Southeastern East China Sea. Appl. Sci. 2021, 11, 7545. [Google Scholar] [CrossRef]
- Chen, Q.C.; Zhang, S.Z. The planktonic copepods of the Yellow Sea and the East China Sea. I. Calanoida. Studia Mar. Sin. 1965, 7, 20–131, (In Chinese with English Summary). [Google Scholar]
- Chen, Q.C. Zooplankton of China Seas; Science Press: Beijing, China, 1992; Volume 1, pp. 1–87. ISBN 7-03-002599-7. [Google Scholar]
- Chihara, M.; Murano, M. An Illustrated Guide to Marine Plankton in Japan; Tokai University Press: Tokyo, Japan, 1997; pp. 1–1000. [Google Scholar]
- Lian, G.S.; Wang, Y.G.; Sun, R.X.; Hwang, J.S. Species Diversity of Marine Planktonic Copepods in China’s Seas; China Ocean Press: Beijing, China, 2019; p. 868. [Google Scholar]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Peter, R.; Minchin, R.B.; O’Hara, G.; Simpson, L.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 25 August 2022).
- Simberloff, D.A. Flagships, umbrellas, and keystones: Is single-species management pass in the landscape era. Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of forest sustainability biodiversity: The selection of forest indicator species. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Guan, B.X.; Fang, G.H. Winter counter-wind currents off the Southeastern China coast: A review. J. Oceanogr. 2006, 62, 1–24. [Google Scholar] [CrossRef]
- Liu, Z.; Gan, J.; Wu, H.; Deng, Y. Progress on circulation dynamics in the East China Sea and southern Yellow Sea: Origination, pathways, and destinations of shelf currents. Prog. Oceanogr. 2021, 193, 102553. [Google Scholar] [CrossRef]
- Chao, S.Y. Circulation of the East China Sea, a numerical study. J. Oceanogr. 1990, 46, 273–295. [Google Scholar] [CrossRef]
- Tang, T.Y.; Hsueh, Y.; Yang, Y.J.; Ma, J.C. Continental slope flow northeast of Taiwan. J. Oceanogr. 1999, 29, 1353–1362. [Google Scholar] [CrossRef]
- Gawarkiewicz, G.; Jan, S.; Lermusiaux, P.F.J.; Mcclean, J.; Centurioni, L.; Taylor, K.; Cornuelle, B.; Duda, T.F.; Wang, J.; Yang, Y.J.; et al. Circulation and intrusions northeast of Taiwan: Chasing and predicting uncertainty in the cold dome. Oceanography 2011, 24, 110–121. [Google Scholar] [CrossRef]
- Lee, H.J.; Chao, S.Y. A climatological description of circulation in and around the East China Sea. Deep-Sea Res. II 2003, 50, 1065–1084. [Google Scholar] [CrossRef]
- Liang, W.D.; Tang, T.Y.; Yang, T.J.; Ko, M.T.; Chuang, W.S. Upper-ocean currents around Taiwan. Deep-Sea Res. II 2003, 50, 1085–1106. [Google Scholar] [CrossRef]
- Liu, K.K.; Peng, T.H.; Shaw, P.T.; Shiah, F.K. Circulation and biogeochemical processes in the East China Sea and the vicinity of Taiwan: An overview and brief synthesis. Deep-Sea Res. II 2003, 50, 1055–1064. [Google Scholar] [CrossRef]
- Tseng, R.S.; Shen, Y.T. Lagrangian observations of surface flow patterns in the vicinity of Taiwan. Deep-Sea Res.II 2003, 50, 1107–1116. [Google Scholar] [CrossRef]
- Jan, S.; Chen, C.C.; Tsai, Y.L.; Yang, Y.J.; Wang, J.; Chern, C.S.; Gawarkiewicz, G.; Lien, R.C.; Centurioni, L.; Kuo, J.Y. Mean structure and variability of the cold dome Northeast of Taiwan. Oceanography 2011, 24, 100–109. [Google Scholar] [CrossRef]
- Wang, J.; Oey, Y. Seasonal exchanges of the Kuroshio and shelf waters and their impacts on the shelf currents of the East China Sea. J. Phys. Oceanogr. 2016, 46, 1615–1632. [Google Scholar] [CrossRef]
- Hsu, P.C.; Zheng, Q.; Lu, C.Y.; Cheng, K.H.; Lee, H.J.; Ho, C.R. Interaction of coastal countercurrent in I-Lan Bay with the Kuroshio Northeast of Taiwan. Cont. Shelf Res. 2018, 171, 30–41. [Google Scholar] [CrossRef]
- Zhou, P.; Song, X.X.; Yuan, Y.Q.; Wang, W.T.; Cao, X.H.; Yu, Z.M. Intrusion pattern of the Kuroshio subsurface water onto the East China Sea continental shelf traced by dissolved inorganic iodine species during the spring and autumn of 2014. Mar. Chem. 2017, 196, 24–34. [Google Scholar] [CrossRef]
- Webber, M.; Edwards-Myers, E.; Campbell, C.; Webber, D. Phytoplankton and zooplankton as indicators of water quality in Discovery Bay, Jamaica. Hydrobiologia 2005, 545, 177–193. [Google Scholar] [CrossRef]
- McKinnon, A.D.; Duggan, S. Community Ecology of Pelagic Copepods in Tropical Waters. In Copepods: Diversity, Habitat and Behavior; Nova Science Publisher: New York, NY, USA, 2014; pp. 25–49. [Google Scholar]
- Polunia, J.J.; Lange, E.K.; Krechik, V.A. Structure and distribution of autumn zooplankton in the Southeastern Baltic Sea in 2015. Oceanology 2019, 59, 66–74. [Google Scholar] [CrossRef]
- Anandavelu, I.; Robin, R.S.; Purvaja, R.; Ganguly, D.; Ramesh, R.R. Spatial heterogeneity of mesozooplankton along the tropical coastal waters. Cont. Shelf Res. 2020, 206, 104193. [Google Scholar] [CrossRef]
- Tseng, L.C.; Dahms, H.U.; Chen, Q.C.; Hwang, J.S. Mesozooplankton and copepod community structures in the southern East China Sea: The status during the monsoonal transition period in September. Helgol. Mar. Res. 2012, 66, 621–634. [Google Scholar] [CrossRef]
- Shi, C.T.; Young, S.S. A checklist of free-living copepods, including those associated with invertebrates, reported from the adjacent seas of Taiwan. Acta Zool. Taiwanica 1995, 6, 65–81. [Google Scholar]
- Gomez-Gutierrez, J.; Funes-Rodriguez, R.; Arroyo-Ramirez, K.; Sanchez-Ortiz, C.A.; Beltran-Castro, J.R.; Hernandez-Trujillo, S.; Palomares-Garcia, J.R.; Aburto-Oropeza, O.; Ezcurra, E. Oceanographic mechanisms that possibly explain dominance of neritic-tropical zooplankton species assemblages around the Islas Marias Archipelago, Mexico. Lat. Am. J. Aquat. Res. 2014, 42, 1009–1034. [Google Scholar] [CrossRef]
- Beltran-Castro, J.R.; Hernandez_Trujillo, S.; Gomez-Gutierrez, J.; Trasvia-Castro, A.; Aburto-Oropeza, O. Copepod species assemblage and carbon biomass during two anomalous warm periods of distinct origin during 2014–2015 in the southern Gulf of California. Cont. Shelf Res. 2020, 207, 104215. [Google Scholar] [CrossRef]
- Kazmi, Q.B. Copepods from shore and offshore waters of Pakistan. J. Mar. Sci. Technol. 2004, 12, 223–238. [Google Scholar] [CrossRef]
- Rakhesh, M.; Raman, A.V.; Sudarsan, D. Discriminating zooplankton assemblages in neritic and oceanic waters: A case for the northeast coast of India, Bay of Bengal. Mar. Environ. Res. 2006, 61, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Jessica, G.; Thomas, H.; Ismar, S.M.H.; Sommer, U. Combined effects of ocean warming and acidification on copepod abundance, body size and fatty acid content. PLoS ONE 2016, 11, e0155952. [Google Scholar]
- Fitzer, S.C.; Caldwell, G.S.; Close, A.J.; Clare, A.S.; Upstill-Goddard, R.C.; Bentley, M.G. Ocean acidification induces multi-generational decline in copepod naupliar production with possible conflict for reproductive resource allocation. J. Exp. Mar. Biol. Ecol. 2012, 418, 30–36. [Google Scholar] [CrossRef]
- Aguilera, V.M. pH and other upwelling hydrographic drivers in regulating copepod reproduction during the 2015 El Niño event: A follow-up study. Estuar. Coast. Shelf Sci. 2020, 234, 106640. [Google Scholar] [CrossRef]
- Lee, E.H.; Choi, S.Y.; Seo, M.H.; Lee, S.J.; Soh, H.Y. Effects of temperature and pH on the egg production and hatching success of a common Korean copepod. Diversity 2020, 12, 372. [Google Scholar] [CrossRef]
- Halsband, C.; Dix, M.F.; Sperrea, K.H.; Reinardy, H.C. Reduced pH increases mortality and genotoxicity in an Arctic coastal copepod, Acartia longiremis. Aquat. Toxicol. 2021, 239, 105961. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | MA | RA (%) | OR (%) |
|---|---|---|---|
| Calanoid copepodites | 217.24 | 28.06 | 100.0 |
| Paracalanus aculeatus | 142.79 | 18.44 | 88.0 |
| Clausocalanus furcatus | 37.14 | 4.80 | 96.0 |
| Canthocalanus pauper | 27.79 | 3.59 | 96.0 |
| Oncaea venusta | 19.30 | 2.49 | 100.0 |
| Acrocalanus gracilis | 20.10 | 2.60 | 92.0 |
| Paracalanus parvus | 25.02 | 3.23 | 68.0 |
| Temora turbinata | 25.89 | 3.34 | 64.0 |
| Farranula gibbula | 18.08 | 2.34 | 88.0 |
| Oncaea media | 18.36 | 2.37 | 72.0 |
| Species | Group A | Group B | Group C | |||
|---|---|---|---|---|---|---|
| Abundance | IndVal | Abundance | IndVal | Abundance | IndVal | |
| Acartia pacifica | 18.83 | 58.83 | 7.8 | 23.92 | 0.04 | 0.04 |
| Acrocalanus monachus | 1.47 | 3.86 | 0.63 | 1.8 | 4.25 | 50.22 |
| Calanopia elliptica | 8.43 | 67.25 | 1.71 | 11.91 | 0.31 | 0.73 |
| Candacia aethiopica | 0.17 | 1.29 | 0.35 | 5.88 | 1.66 | 66.60 |
| Candacia (copepodites) | / | / | / | / | 5.15 | 75.00 |
| Canthocalanus pauper | 60.86 | 65.25 | 23.44 | 22.85 | 8.96 | 9.61 |
| Centropages calaninus | / | / | 0.25 | 4.89 | 1.13 | 61.56 |
| Centropages furcatus | 0.27 | 14.72 | 1.28 | 52.4 | / | / |
| Copilia mirabilis | 0.1 | 1.49 | 0.17 | 2.74 | 3.09 | 92.01 |
| Corycaeus agilis | 4.03 | 2.1 | 10.26 | 23.38 | 17.63 | 55.22 |
| Corycaeus speciosus | 7.62 | 55.44 | 1.52 | 12.07 | 2.31 | 15.15 |
| Euchaeta concinna | 12.29 | 61.12 | 4.36 | 23.68 | 0.1 | 0.15 |
| Euchaeta (copepodites) | 31.1 | 20.96 | 62.2 | 51.45 | 5.61 | 1.42 |
| Farranula concinna | 1.31 | 0.62 | 0.69 | 0.71 | 33.32 | 94.36 |
| Farranula gibbula | 8.34 | 10.67 | 17.4 | 30.38 | 26.33 | 50.58 |
| Macrosetella gracilis | 15.41 | 57.31 | 7.08 | 26.35 | 4.39 | 16.34 |
| Oncaea clevei | 8.74 | 18.86 | 21.31 | 55.19 | 8.56 | 11.09 |
| Paracalanus aculeatus | 184.75 | 44.96 | 217.13 | 52.83 | 9.09 | 1.38 |
| Paracalanus parvus | 42.82 | 53.58 | 23.94 | 19.06 | 13.16 | 8.23 |
| Paracandacia truncata | 0.17 | 8.03 | 1.18 | 69.47 | 0.04 | 1.53 |
| Temora discaudata | 10.66 | 61.82 | 4.5 | 26.08 | 2.09 | 7.56 |
| Temora turbinata | 78.28 | 69.06 | 16.07 | 15.47 | 0.1 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-G.; Tseng, L.-C.; Sun, R.-X.; Chen, X.-Y.; Xiang, P.; Wang, C.-G.; Xing, B.-P.; Hwang, J.-S. Copepods as Indicators of Different Water Masses during the Northeast Monsoon Prevailing Period in the Northeast Taiwan. Biology 2022, 11, 1357. https://doi.org/10.3390/biology11091357
Wang Y-G, Tseng L-C, Sun R-X, Chen X-Y, Xiang P, Wang C-G, Xing B-P, Hwang J-S. Copepods as Indicators of Different Water Masses during the Northeast Monsoon Prevailing Period in the Northeast Taiwan. Biology. 2022; 11(9):1357. https://doi.org/10.3390/biology11091357
Chicago/Turabian StyleWang, Yan-Guo, Li-Chun Tseng, Rou-Xin Sun, Xiao-Yin Chen, Peng Xiang, Chun-Guang Wang, Bing-Peng Xing, and Jiang-Shiou Hwang. 2022. "Copepods as Indicators of Different Water Masses during the Northeast Monsoon Prevailing Period in the Northeast Taiwan" Biology 11, no. 9: 1357. https://doi.org/10.3390/biology11091357
APA StyleWang, Y.-G., Tseng, L.-C., Sun, R.-X., Chen, X.-Y., Xiang, P., Wang, C.-G., Xing, B.-P., & Hwang, J.-S. (2022). Copepods as Indicators of Different Water Masses during the Northeast Monsoon Prevailing Period in the Northeast Taiwan. Biology, 11(9), 1357. https://doi.org/10.3390/biology11091357