Simple Summary
Although locusts can cause major agricultural damage, they also constitute a valuable food resource. At present, L. migratoria manilensis has been largely domesticated, being considered an edible insect in China. Feeding variety is one of the main characteristics of L. migratoria manilensis. There are apparent differences in the capacity of locusts to adapt to different food plants. To elucidate the effect of different food plants (i.e., goosegrass, maize leaves, soybean leaves, and pakchoi) on the growth and development of L. migratoria manilensis, the gut bacterial community composition of L. migratoria manilensis fifth instars fed on different plants was analyzed by high-throughput sequencing. Gut bacterial communities were affected by food plants and may play an essential role in host adaption. Feeding on different food plants has significant effects on the growth and development of L. migratoria manilensis. The present study establishes a theoretical foundation for studying the interplay between gut bacteria structure and L. migratoria manilensis adaptation.
Abstract
Locusts, in particular Locusta migratoria manilensis (Meyen), have been associated with major damages in agriculture, forestry, and animal husbandry in China. At present, L. migratoria manilensis has been largely domesticated, being considered an edible insect in China. Feeding variety is one of the main characteristics of L. migratoria manilensis. It has been demonstrated that microorganisms inhabiting the insect gut impact nutrition, development, defense, and reproduction of the insect host. The aim of the present study was to search for the adaptation mechanism of L. migratoria manilensis feeding on four different food plants (goosegrass, maize leaves, soybean leaves, and pakchoi) and explore changes in the gut bacterial community structure of the insect at the fifth instar nymph stage. Proteobacteria and Firmicutes were the dominant phyla, whereas Kluyvera, Enterobacter, Pseudocitrobacter, Klebsiella, Cronobacter, Citrobacter, Lactococcus, and Weissella were the dominant genera in the gut of L. migratoria manilensis. Principal component analysis and permutational multivariate analysis of variance (PERMANOVA) revealed significant differences in the gut microbiota structure of L. migratoria manilensis fed on different food plants. Moreover, functional prediction analysis revealed that metabolic and cellular processes were the most enriched categories. Within the category of metabolic processes, the most enriched pathways were carbohydrate transport and metabolism; amino acid transport and metabolism; translation, ribosomal structure, and biogenesis; cell wall/membrane/envelope biogenesis; inorganic ion transport and metabolism; and energy production and conversion. Collectively, the present results revealed that the structure of gut bacterial communities in L. migratoria manilensis fed on different food plants is impacted by food plants, which may play an essential part in the adaptation of the host.
1. Introduction
Locusts have long been known and valued in China since ancient times. Among the approximately 900 species of locusts known to date, Locusta migratoria manilensis (Meyen) is the leading species causing infestations in China [1]. Although locusts can cause major agricultural damage, they also constitute a valuable food resource. As a primary consumer in the food chain, locusts transform weeds, straws, and other unavailable carbohydrates and polysaccharides into assimilable protein resources, which can be used as feed alternatives for livestock and poultry farming, as well be considered a culinary specialty. As an important food resource, locusts can be raised in captivity. At present, L. migratoria manilensis has been largely domesticated, being considered an edible insect in China [2,3,4,5]. However, much of the costs associated with locust farming are spent on feed. Different food plants directly influence the growth and development of locusts.
Under natural conditions, plants of the two families, Gramineae and Cyperaceae, are the primary hosts of locusts. Plants of the Dicotyledon can also be temporary hosts, although are less favorable for the growth and reproduction of locusts. There are apparent differences in the capacity of locusts to adapt to different food plants [6]. Typical green forage used in the production of L. migratoria manilensis includes maize straw and weeds, occasionally combined with fruits, bean leaves, and vegetables. Similarly, differences in the type of forage can impact the development of L. migratoria manilensis [7,8,9]. It has been found that the best feed intake, weight gain, growth rate, and lowest mortality rate in L. migratoria manilensis are observed when feeding on Gramineae; in contrast, relatively low feed intake, weight gain, growth rate, and high mortality rate are observed when L. migratoria manilensis fed on lettuce leaves and carrots, which can be used as supplementary feed [7]. In addition, it has been shown that a high carrot content in the diet of farmed L. migratoria manilensis reportedly leads to an increase in the contents of lipids and vitamin A, whereas protein content decreases with an increase in the amount of wheat bran in the diet [10].
Insect biology and behavior must be studied as a complex ecological system of which microorganisms constitute a significant part [11,12]. The insect gut provides a particular habitat for a wide variety and number of microorganisms. The insect gut environment is affected by changes in the outer environment, and gut microecology diversity is closely related to insect species and feeding habits [13,14]. In long-term coevolution, insects and gut microorganisms have developed a symbiotic relationship [15], since the insect gut constitutes a stable environment and is a source of essential nutrients for gut microorganisms. In exchange, gut microorganisms are involved in various metabolic processes of the insect host, providing nutrients and digesting complex carbohydrates [16,17]. Studies have found that gut microorganisms can enhance host nutritional status and metabolism by synthesizing nutrients that are lacking in foods, but are essential to the host, as well as by secreting digestive enzymes that allow the host to digest certain food components. In addition, insect gut microorganisms protect the host against invasive pathogens, improve immunity, degrade exogenous biological toxins, facilitate interspecific and intraspecific communication, modulate reproduction, and enhance growth and development [18,19,20,21,22,23,24,25]. Significant differences have been described in gut microbiota composition among insects fed on different foods [26,27,28]. Hence, food is a central element affecting the composition of insect gut microbiota [29,30,31].
Therefore, the aim of the present study was to explore the effect of different food plants (i.e., goosegrass, maize leaves, soybean leaves, and pakchoi) on the growth and development of L. migratoria manilensis. In addition, gut bacterial community composition of L. migratoria manilensis fifth instars fed on different plants was analyzed by high-throughput sequencing. Furthermore, functional annotation of metagenomes was conducted to provide a basis for subsequent in-depth studies of the interplay between gut microbiota and food plants of L. migratoria.
2. Materials and Methods
2.1. Insect Rearing
L. migratoria manilensis (Meyen) were purchased from an insect breeding base in Cangzhou, Hebei, China, and reared under the following conditions: 80–100 heads per breeding cage (25 cm × 25 cm × 50 cm; L × W × H) at 28 ± 0.5 °C under 70 ± 10% relative humidity (RH) using a photoperiod (L:D) of 16:8 h and three consecutive generations. Fresh plants were inserted into water-containing sponges, and feeding and weighing were conducted daily. Insects’ developmental period, weight gain, mortality, dung production, and food intake were recorded for the calculation of each nutritional index. Newly hatched nymphs were rare in goosegrass (Gg), maize leaves (ML), soybean leaves (SL), and pakchoi (Pc) until the fifth instar stage. Experimental food plants were grown in the practical base of plant protection at Shandong Agricultural University, China. All samples for the present experiment were collected at the fifth instar nymph stage.
2.2. DNA Extraction
Randomly, 60 nymphs of comparable size and growth stage at the fifth instar stage were collected. After starving for two days, the surface of nymph was repeatedly washed with sterile water, then placed in 75% alcohol solution for 2 min, then rinsed with sterile water and irradiated with UV light for 3–5 min. Locust nymphs were dissected under aseptic conditions, and the whole intestine was removed; specifically, mid and hind intestines were intercepted, after which were rinsed with sterile phosphate-buffered saline (PBS) (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) [15,32], and intestinal contents of each sample were collected in sterile centrifuge tubes.
Total microbial DNA was extracted using the PowerSoil DNA Isolation Kit (MoBio, Qiagen Inc., Germantown, MD, USA) following the manufacturer’s instructions. The quality of extracted DNA was verified by horizontal gel electrophoresis in 0.8% agarose gels. Quantification of extracted DNA was determined by NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA) UV-vis spectrophotometer. Each sample was conducted in three replicates.
2.3. Metagenomic Analysis
The universal primer pair, 341F (5′-CCTACGGGNGGCWGCAG-3′) and 805R (5′-GACTACHVGGGTATCTAATCC-3′), with a sequencing adapter at the end, were used to amplify the V3-V4 hypervariable region of the bacterial 16s rDNA gene. The PCR reaction system was composed of: PCR Phusion High-Fidelity PCR Master Mix with HF Buffer (New England Biolabs, Ipswich, MA, USA), 25 μL; DMSO, 3 μL; 3 μL of each primer; gDNA, 10 μL; nuclease-free water, q.s. to a final volume of 50 μL. PCR cycling parameters were as follows: 98 °C for 30 s; followed by 30 amplification cycles of 98 °C for 15 s, 58 °C for 15 s, and 72 °C for 15 s; followed by 1 min of final extension at 72 °C and an indefinite hold at 4 °C. PCR products were separated on 2% agarose gels and then submitted to purification.
PCR amplification of recovery products was quantified by fluorescence determination using fluorescent reagents provided within the Quant-iT PicoGreen dsDNA Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) in a FLx800 microplate reader (BioTek Instruments, Winooski, VE, USA). Based on fluorescence quantification, each sample was mixed in an appropriate proportion according to the sequencing volume required. Sequencing libraries were prepared using the TruSeq Nano DNA LT Library Prep Kit (Illumina, San Diego, CA, USA), and 2 × 300 bp double-end sequencing was performed using a MiSeq platform with the MiSeq Reagent Kit V3 (Illumina, San Diego, CA, USA) (600 cycles). Sequencing was conducted at Shandong Kaiyuan Gene Technology Co., Ltd.
2.4. Statistical and Bioinformatic Analysis
Firstly, quality control of raw data was conducted using Trimmomatic (v0.39) software [33]. Based on the overlap (minimum: 10 bp) between PE reads after quality control, PE reads were spliced through overlap by Flash (v1.2.11) software [34]. We used UCHIME (v4.2) [35] for the identification and removal of chimeric sequences to obtain the valid data. Based on the sequence similarity, the valid sequences were classified into multiple operational taxonomic units (OTUs) by the software VSEARCH (v2.16.0) [36] at the similarity level of 97%. All representative sequences were annotated and blasted against Silva database (v138.1, http://www.arb-silva.de (accessed on 4 January 2022)) using RDP Classifier (v2.11) [37] with a confidence threshold at 80%.
Mothur (v1.46.1) software [38] was used to calculate alpha-diversity indexes, which included ACE, Chao1, the Simpson index, and the Shannon index. In addition, principal coordinate analysis (PCoA) was used to reveal differences in gut flora composition between sample groups. Permutational multivariate analysis of variance (PERMANOVA) was performed for pairwise comparison of samples. Linear discriminant analysis (LDA) was used to identify biomarkers with statistical significance between samples with LDA scores greater than 4. Gut metagenome functions were determined by annotating pathways of OTUs against the Clusters of Orthologous Genes (COG) database using PICRUSt2 (v. 2.5.0) [39]. Differences were considered significant when p values were < 0.05 and highly significant when p values were < 0.01. SPSS 26.0 (https://www.ibm.com/products/spss-statistics (accessed on 16 March 2022)) was used for statistical analysis. The calculation formula is as follows.
3. Results
3.1. Development Rate of L. migratoria manilensis Reared on Different Food Plant
L. migratoria manilensis had the highest food utilization rate, weight gain, and the lowest mortality rate when grown on maize leaves (ML), followed by goosegrass (Gg), soybean leaves (SL), and pakchoi (Pc) (Table 1, Figure 1). In contrast, the average generation period was intermediate for L. migratoria manilensis fed on SL; the nymph stage was the longest, and the adult stage was the shortest in L. migratoria manilensis fed on Pc. All four plants species completed their life cycles (Table 2). The growth rate and overall vitality of L. migratoria manilensis fed on ML were significantly higher than those fed on the other food plants, with the shortest developmental period. The growth rate and overall vitality of L. migratoria manilensis fed on Pc were substantially lower than those fed on the other food plants, with the longest developmental period (Figure 2).

Table 1.
Productive parameters of the nymph and Egg production of the female of Locusta migratoria manilensis reared on four food plants.
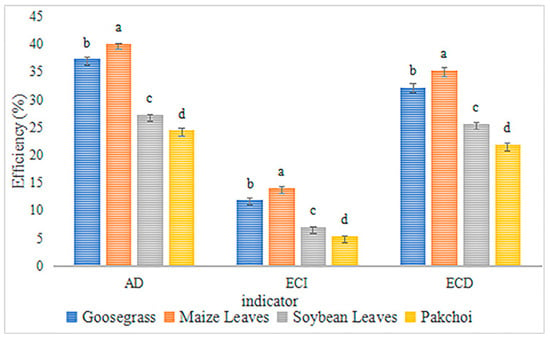
Figure 1.
Food utilization efficiency by Locusta migratoria manilensis (Meyen) reared on four food plants. AD, ECI, and ECD indicated approximate digestibility (means ± SE), efficiency of conversion of ingested food (mean ± SE) and efficiency of conversion of digested food (mean ± SE), respectively. Significant differences within AD, ECl, and ECD are indicated by lowercase letters (ANOVA, p < 0.05), respectively.
Table 2.
Developmental durations of L. migratoria manilensis reared on four food plants.
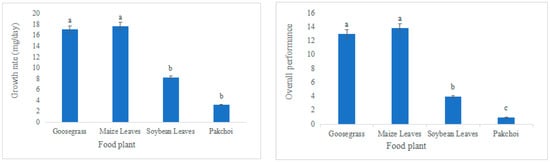
Figure 2.
Growth rate and overall performance of Locusta migratoria manilensis reared on four food plants. Bars marked by different lowercase letters are significantly different based on Turkey’s HSD analysis at p < 0.05.
3.2. Metagenomic Analysis
Four groups of 12 samples were sequenced. A total of 1,265,960 reads were obtained. After quality control, a total of 1,236,884 effective reads remained (Table S1). Cluster analysis resulted in 2838 OTUs, which corresponded to 26 phyla, 55 classes, 111 orders, 155 families, 247 genera, and 387 species. The sample scarcity curve (Figure S1A) and the Shannon diversity index scarcity curve (Figure S1B) indicate that sequencing volume was sufficient, the sequencing depth was saturated, and increasing the sample volume would not produce more OTUs. Moreover, Good’s coverage index was used to verify sequencing completeness. The results showed that sequencing coverage was greater than 99%, showing that most microbial species found in the samples were characterized.
3.3. Metagenomic Analysis
For alpha-diversity analysis, ACE and Chao1 indexes reflect the richness of the microbial community in a sample; the larger the value of these indexes, the higher the community richness. The gut microbiota in L. migratoria manilensis fed on SL had a lower abundance compared to the other treatment groups. In addition, Shannon and Simpson indexes reflect diversity in the gut microbial community; a higher diversity in a sample is indicated by higher Shannon index values and lower Simpson index values. No significant difference was found among the samples across the four feeding conditions. Thus, these results showed that the gut microbiota of L. migratoria manilensis reared on four different food plants had high species diversity and richness, although no significant difference was found among the samples (Figure 3).
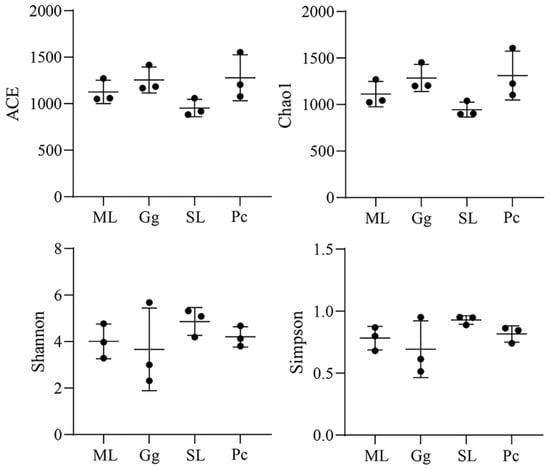
Figure 3.
Box plots of ACE, Chao 1, and Shannon and Simpson indexes of gut microbial communities in Locusta migratoria manilensis fed on four different food plants. ML: maize leaves; Gg: goosegrass; SL: soybean leaves; Pc: pakchoi.
At the phylum level, the top ten phyla in relative abundance were Proteobacteria, Firmicutes, Bacteroidota, Actinobacteriota, Acidobacteriota, Verrucomicrobiota, Planctomycetota, Chloroflexi, Gemmatimonadota, and Myxococcota (Figure 4A). Among them, Proteobacteria was the dominant phylum, showing the highest relative abundance in L. migratoria manilensis fed on Pc (94.66 ± 4.511%); the relative abundance of Proteobacteria in the other feeding groups was as follows: 77.18 ± 9.762% in L. migratoria manilensis fed on ML; 87.86 ± 6.509% on Gg; and 45.19 ± 9.619% on SL. In contrast, Firmicutes was the dominant phylum in the gut microbiota of L. migratoria manilensis fed on SL, whose relative abundance was 54.80 ± 9.63% (Figure S2).
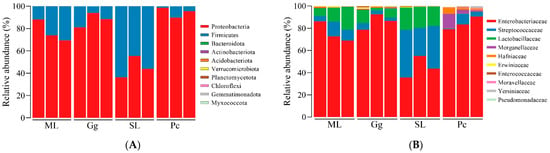
Figure 4.
Bacterial composition of the top 10 relative abundances of bacteria at the phylum (A) and (B) family levels in the gut of Locusta migratoria manilensis fed on four different food plants. ML: maize leaves; Gg: goosegrass; SL: soybean leaves; Pc: pakchoi. Each color represents a phylum in (A) and a family in (B), and the height of the color block represents the proportion of relative abundance of that species.
At the family level, the top ten families in relative abundance were Enterobacteriaceae, Streptococcaceae, Lactobacillaceae, Morganellaceae, Hafniaceae, Erwiniaceae, Enterococcaceae, Moraxellaceae, Yersiniaceae, and Pseudomonadaceae (Figure 4B). The highest relative abundances of Enterobacteriaceae were observed in the gut microbiota of L. migratoria manilensis fed on ML, Gg, and Pc (75.88 ± 9.106%, 85.92 ± 7.005%, and 84.37 ± 5.735%, respectively). In contrast, the relative abundance of Enterobacteriaceae in L. migratoria manilensis fed on SL was 44.79 ± 9.69%. The relative abundance of Streptococcaceae and Lactobacillaceae was significantly higher in the gut microbiota of L. migratoria manilensis fed on SL (35.56 ± 9.11% and 18.85 ± 1.28%, respectively) (Table S2).
For beta-diversity analysis, a clustering heat map was created to reveal the dynamics of L. migratoria manilensis gut microbiota fed on different food plants, based on the top 20 relative abundances of bacteria at the genus level. Collectively, the gut microbiota of L. migratoria manilensis fed on ML, Gg, and Pc was similar in composition at the genus level, in which Kluyvera, Enterobacter, and Pseudocitrobacter were the dominant genera. In contrast, Lactococcus and Weissella were the dominant genera in the gut microbiota of L. migratoria manilensis fed on SL, with Lactococcus present in significantly higher relative abundance compared to the other groups (Figure 5).
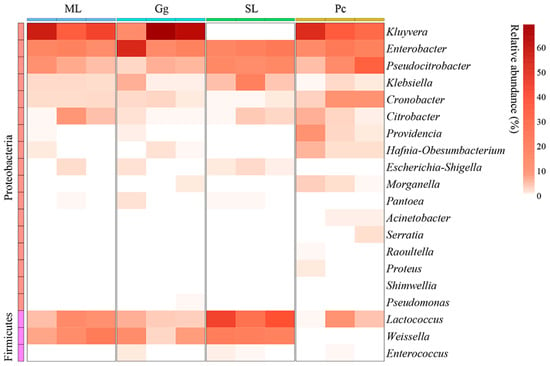
Figure 5.
Clustering heat map of the 20 most abundant genera in the gut bacterial community of Locusta migratoria manilensis fed on four different food plants. ML: maize leaves; Gg: goosegrass; SL: soybean leaves; Pc: pakchoi.
In order to identify biomarkers of bacteria and different levels of taxa change that could enable distinguishing the different sample groups, LDA effect size (LEfSe) was used on OTUs at various taxonomic ranks (kingdom, phylum, class, order, family, genus, and species) with standard LDA values > 4 (Figure S3). In addition, a cladogram at taxonomic ranks phylum, class, order, family, genus, and species was used to elucidate the distribution of changes at various taxonomic ranks (Figure 6).
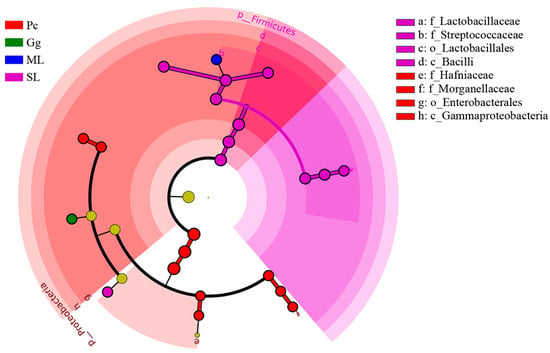
Figure 6.
Cladogram of bacterial biomarkers from the phylum to genus (innermost ring to outermost ring) level, with LDA score > 4. ML: maize leaves; Gg: goosegrass; SL: soybean leaves; Pc: pakchoi. Differential bacterial taxa are marked by lowercase letters. The circles at different taxonomic levels represent a taxon at that level, and the diameter of the circle corresponds to the relative abundance. Different colors represent different groups species, with yellow representing species with insignificant differences and the other different species as the group with the highest abundance of the species.
The gut bacterial community of L. migratoria manilensis fed on different food plants was composed of different taxa (LDA > 4), mainly Proteobacteria and Firmicutes. There were 11 different taxa in SL, primarily Firmicutes; ten groups were found in Pc, mainly Proteobacteria; one different group was found in ML, which corresponded to Firmicutes; one different group was found in Gg, which corresponded to Proteobacteria. Collectively, these results indicated that host plant had a significant effect on shaping the gut microbiota structure of L. migratoria manilensis.
3.4. Functional Annotation of Gut Microbial Community in L. migratoria manilensis
To gain insights into the role of gut microbiota in L. migratoria manilensis, the PICRUSt2 software was used to predict the function of obtained metagenomes by annotating against the COG database.
Overall, the results showed that most functional prediction categories are related to metabolic and cellular processes. The main metabolic functions include carbohydrate transport and metabolism; amino acid transport and metabolism; translation, ribosomal structure and biogenesis; cell wall/membrane/envelope biogenesis; inorganic ion transport and metabolism; energy production and conversion; transcription, thus, representing the most active functions in the gut microbiota of L. migratoria manilensis fed on four different food plants (Figure 7). The functional category translation, ribosomal structure, and biogenesis were significantly more enriched in SL compared to the other treatment groups, whereas differences between the remainder of the samples were not significant (Figure 8D). Moreover, carbohydrate transport and metabolism were found to be enriched the lowest in Pc, and the highest in Gg, whereas differences in the proportion of OTUs mapped to this category were not significant between the other two sample groups (Figure 8B). Considering amino acid transport and metabolism, the lowest enrichment was found in SL, whereas no difference was found among the other three sample groups (Figure 8A). Finally, the proportion of OTUs mapped to nucleotide transport and metabolism category was comparable to those mapped to translation, ribosomal structure, and biogenesis category (Figure 8C).
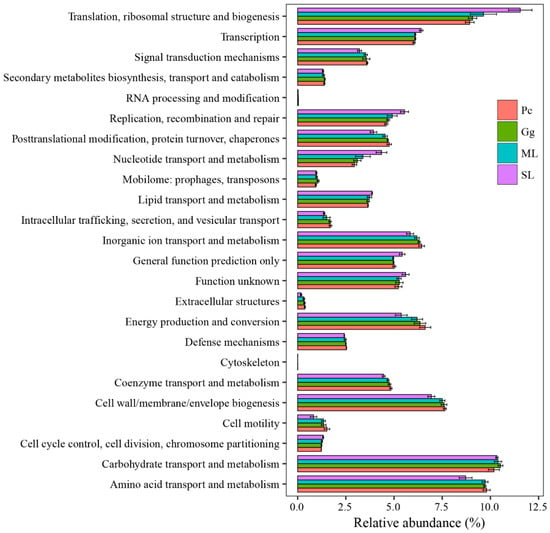
Figure 7.
Comparison of predicted Clusters of Orthologous Genes (COG) functional categories of the gut bacterial community in Locusta migratoria manilensis fed on different food plants. ML: maize leaves; Gg: goosegrass; SL: soybean leaves; Pc: pakchoi.
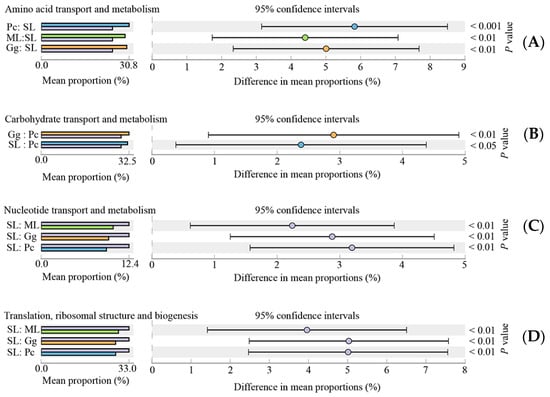
Figure 8.
Significant differences (p < 0.05) in the abundance of operational taxonomic units (OTUs) mapped to predicted Clusters of Orthologous Genes (COG) functional categories amino acid transport and metabolism (A), carbohydrate transport and metabolism (B), nucleotide transport and metabolism (C), and translation, ribosomal structure, and biogenesis (D). ML: maize leaves; Gg: goosegrass; SL: soybean leaves; Pc: pakchoi.
4. Discussion
As the most widely distributed locust species in China, L. migratoria manilensis (Meyen) has a vigorous life cycle, with two to four generations yearly, and can survive under various environmental conditions. In addition, its feeding, migration, and aggregation habits are versatile. It is a typical polyphagous insect that feeds on plants of the families Gramineae and Cyperaceae, as well as occasionally on certain plants of Dicotyledon [40,41,42]. The present study aimed to explore the differences in the gut bacterial community of L. migratoria manilensis fed on four different food plants. L. migratoria manilensis fed on Gramineae showed better growth performance than those fed on Dicotyledon. Significant differences were found in the adaptability of L. migratoria manilensis to different food plant, mainly in terms of growth parameters and differences in food usage. Taken together, the results described herein provide a more integrative understanding of the connections between L. migratoria manilensis and its symbiotic gut microorganisms. Based on the data, it is tentatively concluded that food plants affected the diversity and abundance of the gut bacterial community of L. migratoria manilensis, which revealed a sophisticated relationship between the gut bacteria of L. migratoria manilensis and food plants, thus providing a theoretical basis for comprehending the adaptation mechanisms of L. migratoria manilensis to food plants.
Previous studies have shown that insect gut microbial composition is significantly influenced by host species and diet [32,43]. However, the influence of the host dietary is greater compared to the influence of host species [44,45,46]. The growth rate, fecundity, and survival rate of locusts fed on different food plants differ widely [6,40,47], which will affect gut microorganisms development. Moreover, it has been shown that food plants induce differences in the gut microenvironment, and the diversity in the abundance of gut microbes can change [48]. Therefore, the abundance of gut bacteria depends on the composition and content of food plant upon which locusts feed. Thus, changes in food plant will necessarily lead to changes in the contents of nutrients available to gut bacteria, hence resulting in variations in the gut microbiota composition [49,50,51]. The diversity of gut bacteria in L. migratoria manilensis changed to varying degrees after feeding on four different food plants, and the gut bacterial community changed in response to changes in the dietary. Similar changes were observed in the diversity and structure of gut bacterial communities in Grapholita molesta and Plutella xylostella fed on different food plants [15,52].
Differences in the relative abundance of phyla enable a comprehensive assessment of differences in the composition of the gut bacterial community, as each phylum usually contributes to different functions. Proteobacteria and Firmicutes were the dominant bacterial phyla in the four treatment groups. Many studies have shown that Proteobacteria and Firmicutes predominate in many insects’ gut microbiota [18,53,54,55,56], playing an essential role in carbohydrate metabolism, amino acid metabolism, energy production, and membrane transport [57,58,59]. Moreover, it has been described that colonization of insect gut to form a stable microbiota may be the result of long-term interaction and adaptation between host insects and food plants, which is essential for insects to adapt to feeding on specific food plants [52]. In the present study, the gut bacterial community of L. migratoria manilensis was closely associated with the host plant, which may play an essential part in food ingestion and nutrient utilization. Future research combining metabolomics would elucidate how gut bacterial communities affect insects’ metabolic processes.
Herein, Enterobacteriaceae, Streptococcaceae, and Lactobacillaceae were found to be the predominant families in the gut bacterial community of L. migratoria manilensis in the four treatment groups; in particular, Enterobacteriaceae was the most predominant in all samples. It has been reported that Enterobacteriaceae is commonly found in the gut microbiota of many insects [18,58]. Previous studies have shown that Enterobacteriaceae plays an essential role in the host’s glucose metabolism, digestion, as well as courtship and reproductive behaviors [32,60,61]. In addition, the high abundance of Enterobacteriaceae may be related to host adaptability [15,32]. In the present study, functional prediction analysis revealed that Enterobacteriaceae is related to carbohydrate transport and metabolism. Within the family Enterobacteriaceae, the most abundant genus was Kluyvera, a Gram-negative bacterium with cellulose-degrading and nitrogen fixation abilities that can improve the host’s nutrition by providing nitrogenous compounds necessary for amino acid synthesis [62,63]. Plants of the family Gramineae contain high amounts of cellulose, which may account for the higher abundance of Kluyvera in the samples. In the family Streptococcaceae, the most abundant genus was Lactococcus in SL. It has been reported that Lactococcus can be found in the viscera of different animals (including insects), which may be responsible for lignocellulose digestion and fermentation [20,64]. Combined with functional prediction analysis, Lactococcus could be related to the category translation, ribosomal structure, and biogenesis. The high protein content in SL may account for the high abundance of Lactococcus in this sample.
In addition, the five genera with the highest relative abundance in the gut bacterial community of L. migratoria manilensis fed on Gg and ML were highly comparable, which included Kluyvera, Enterobacter, Pseudocitrobacter, Lactococcus, and Weissella. However, the abundance of these genera varied. The top five genera in terms of relative abundance in the gut bacterial community in L. migratoria manilensis fed on Pc were Kluyvera, Enterobacter, Pseudocitrobacter, Cronobacter, and Lactococcus. In contrast, the top five genera in relative abundance in the gut bacterial community in L. migratoria manilensis fed on SL were Lactococcus, Weissella, Enterobacter, Pseudocitrobacter, and Klebsiella. PERMANOVA analyses revealed substantial differences between treatment groups (R2 = 0.601; p = 0.002). Thus, these results indicate that the abundance of the gut microbiota varied according to the host plant source, even those belonging to the family Gramineae. It has been previously stated that the gut microbiota structure changes considerably according to the host requirements for growth and development [52,65].
Plants of the families Gramineae and Cyperaceae are the primary hosts of locusts in nature, whereas plants of Dicotyledon can also serve as temporary host. Feeding on different food plant had a significant impact on the growth and reproduction of L. migratoria manilensis [6], as well as on species diversity and abundance in the insect gut bacterial community [66]. Thus, it can be stated that there were obvious differences in adaptation to different food plants in L. migratoria manilensis, and plants of the family Gramineae are the most suitable food plant for locusts. However, previous studies have been conducted to identify some gut bacteria by isolation and culture, and have only involved gut microbiota that feeds on plants of the family Gramineae. In the present study, it was shown that the structure of the gut bacterial community of L. migratoria manilensis fed on SL and Pc differed significantly from that of locusts fed on Gg and ML. Moreover, the abundance and diversity of the gut microbiota in SL and Pc treatment groups were not lower than those in Gg and ML treatment groups. Combined with COG functional prediction analysis, it can be stated that the gut microbiota structure of L. migratoria manilensis changed qualitatively and quantitatively in response to the characteristics of food plants in order to adapt to the type of feed source as well as to meet the nutritional requirements of the host.
Collectively, the results discussed in the present study indicated that host plant could affect the gut bacterial structure of L. migratoria manilensis. Gut bacterial communities were affected by the host’s diet and may play an essential role in host adaption. However, gut bacterial structures were measured after three rearing generations in the present study, whereas it would be ideally to explore differences in gut bacterial structure in each generation. Such information would contribute to a higher comprehension of the dynamic changes in the gut bacteria of L. migratoria manilensis during adaptation to various food plants over different generations. A study combining multi-omics techniques to elucidate the vital role of gut bacteria in host adaptation would enable finding new targets for controlling locusts. Finally, the present study establishes a theoretical foundation for studying the interplay between gut bacteria structure and L. migratoria manilensis adaptation.
5. Conclusions
In the present work, we explored the effect of different food plants (i.e., goosegrass, maize leaves, soybean leaves, and pakchoi) on the growth and development of L. migratoria manilensis. In addition, changes in the gut bacterial community structure of the insect at the fifth instar nymph stage were explored by 16S rDNA sequencing. L. migratoria manilensis that fed on Gramineae showed better growth performance than those that fed on Dicotyledon. Collectively, the host plant affected the diversity and abundance of the gut bacterial community of L. migratoria manilensis. The gut microbiota structure of L. migratoria manilensis changed qualitatively and quantitatively in response to the characteristics of food plants in order to adapt to the type of feed, as well as to meet the nutritional requirements of the host. The current study provides a theoretical basis for a better understanding of the adaptation mechanisms of L. migratoria manilensis to its food plant, as well as sheds new light on the role of gut bacteria in host adaptation and nutrition.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biology11091347/s1, Figure S1: (A) Sample rarefaction curves and (B) Shannon index rarefaction curves of individual samples for metagenomic analysis of gut microbiota of Locusta migratoria manilensis fed on four different food plants; Figure S2: The top 5 relative abundances of bacteria at the phylum level in the gut of Locusta migratoria manilensis fed on four different food plants; Figure S3: Histogram of LDA value distribution of the gut bacterial community of Locusta migratoria manilensis fed on different food plants; Table S1: Summary of high-throughput sequencing statistics obtained for each sample evaluated in the present study; Table S2: The top 10 relative abundances at the family level in Locusta migratoria manilensis fed on four different food plants.
Author Contributions
Q.W., Y.L. and X.Y. conceived the study and designed the experiments. Q.W. conducted the experiments, analyzed the data, and wrote the manuscript. Y.L. and X.Y. commented on and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the fund for the China Postdoctoral Science Foundation (No. 2017M612312) and the Shandong Province Agricultural Major Application Technology Innovation Project (No. SD2019ZZ015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original 16S rRNA sequence data are available in the NCBI Sequence Read Archive under accession number PRJNA858491.
Acknowledgments
We would like to thank Ningxin Wang and Jiaxing Fang for their suggestions on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhu, E.L. Prospects of integrated Locust management for twenty-first century in China. In Proceedings of the Symposium on Integrated Pest Management in China, Kunming, China, 2 December 1996; pp. 1031–1036. [Google Scholar]
- Cao, C.Q.; Huang, M.Y.; Chen, S.Z.; Liu, Y.S. Problems and Counter measures for the Resource Utilization of Locust. J. Anhui Agric. Sci. 2008, 36, 15491–15492, 15499. [Google Scholar] [CrossRef]
- Liu, S.P.; He, F.; Feng, W.J.; Li, Q.X. Research progress on edible insects of Orthoptera. Light Ind. Sci. Technol. 2016, 32, 9–11. [Google Scholar]
- Hou, S.K.; Hu, H.Y.; Liu, J.; Zhang, J. Development and Utilization of a New Type of Renewable Animal Protein Feed Resource-Insect Powder. Feed. Rev. 2017, 12, 35–38. [Google Scholar]
- Egonyu, J.P.; Subramanian, S.; Tanga, C.M.; Dubois, T.; Ekesi, S.; Kelemu, S. Global overview of locusts as food, feed and other uses. Glob. Food Secur. 2021, 31, 100574. [Google Scholar] [CrossRef]
- Qin, J.D.; Guo, F.; Zheng, Z.Y. Food specialization and food utilization of the Oriental migratory locust and the influence of different food plants on its growth and fecundity. Acta Entomol. Sin. 1957, 7, 143–166. [Google Scholar] [CrossRef]
- Chen, Y.L. Major achievements in the study of Locusta migratoria and their management in China. Chin. J. Appl. Entomol. 2000, 37, 50–59. [Google Scholar]
- Liu, Y.S. Captive breeding of Locusta migratoria manilensis (Meyen). Sci. Farming 2006, 02, 36–37. [Google Scholar] [CrossRef]
- Zhao, C. Study on the Feeding Ability on Maize Straw of Locusta migratoria manilensis (Meyen). Master’s Thesis, Shandong Agricultural University, Shandong, China, 2015. [Google Scholar]
- Salama, S.M. Nutrient Composition and Bioactive Components of the Migratory Locust (Locusta migratoria). In African Edible Insects as Alternative Source of Food, Oil, Protein and Bioactive Components; Adam Mariod, A., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 231–239. [Google Scholar]
- Steinhaus, E.A., II. The importance of environmental factors in the insect-microbe ecosystem. Microbiol. Mol. Biol. Rev. 1960, 24, 365–373. [Google Scholar] [CrossRef]
- Fang, J.; Liu, M.; Zhang, S.; Liu, F.; Zhang, Z.; Zhang, Q.; Kong, X. Chemical signal interactions of the bark beetle with fungal symbionts, and host/non-host trees. J. Exp. Bot. 2020, 71, 6084–6091. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B. Advances in the study of insect gut microbes. Technol. Innov. Appl. 2017, 05, 50. [Google Scholar]
- Mei, C.; Fan, S.; Yang, H. The strategies of isolation of insect gut microorganisms. Acta Microbiol. Sin. 2018, 58, 985–994. [Google Scholar] [CrossRef]
- Yuan, X.Q.; Zhang, X.; Liu, X.Y.; Dong, Y.L.; Yan, Z.Z.; Lv, D.B.; Wang, P.; Li, Y.P. Comparison of Gut Bacterial Communities of Grapholita molesta (Lepidoptera: Tortricidae) Reared on Different Host Plants. Int. J. Mol. Sci. 2021, 22, 6843. [Google Scholar] [CrossRef] [PubMed]
- Cazemier, A.E.; Hackstein, J.H.P.; Op Den Camp, H.J.M.; Rosenberg, J.; van der Drift, C. Bacteria in the Intestinal Tract of Different Species of Arthropods. Microb. Ecol. 1997, 33, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.J.; Dillon, V.M. THE GUT BACTERIA OF INSECTS: Nonpathogenic Interactions. Annu. Rev. Entomol. 2004, 49, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.; Charnley, K. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef]
- Bashir, Z.; Kondapalli, V.K.; Adlakha, N.; Sharma, A.; Bhatnagar, R.K.; Chandel, G.; Yazdani, S.S. Diversity and functional significance of cellulolytic microbes living in termite, pill-bug and stem-borer guts. Sci. Rep. 2013, 3, 2558. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects-diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Lee, K.-A.; Kim, S.-H.; Kim, E.-K.; Ha, E.-M.; You, H.; Kim, B.; Kim, M.-J.; Kwon, Y.; Ryu, J.-H.; Lee, W.-J. Bacterial-Derived Uracil as a Modulator of Mucosal Immunity and Gut-Microbe Homeostasis in Drosophila. Cell 2013, 153, 797–811. [Google Scholar] [CrossRef]
- Calcagnile, M.; Tredici, S.M.; Talà, A.; Alifano, P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects 2019, 10, 441. [Google Scholar] [CrossRef]
- Suenami, S.; Nobu, M.K.; Miyazaki, R. Community analysis of gut microbiota in hornets, the largest eusocial wasps, Vespa mandarinia and V. simillima. Sci. Rep. 2019, 9, 9830. [Google Scholar] [CrossRef]
- Li, F.; Huang, S.S. Isolation and Identification of Lignin-degrading Bacteria from the Gut of Termite and Their Degradation Characteristics. Biotechnol. Bull. 2020, 36, 61–68. [Google Scholar]
- Wang, S.C.; Wang, L.Y.; Fan, X.; Yu, C.; Feng, L.; Yi, L. An Insight into Diversity and Functionalities of Gut Microbiota in Insects. Curr. Microbiol. 2020, 77, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.Q.; Wang, X.Q.; Feng, W.; Zhou, W.; Xie, H.X.; Wan, Y.J. Comparative analysis of the composition of dominant intestinal microflora in silkworm reared with different forages. Acta Ecol. Sin. 2010, 30, 3875–3882. [Google Scholar]
- Pérez-Cobas, A.E.; Maiques, E.; Angelova, A.; Carrasco, P.; Moya, A.; Latorre, A. Diet shapes the gut microbiota of the omnivorous cockroach Blattella germanica. FEMS Microbiol. Ecol. 2015, 91, 22. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Choi, M.-Y.; Kim, J.-W.; Lee, S.A.; Ahn, J.-H.; Song, J.; Kim, S.-H.; Weon, H.-Y. Effects of diet type, developmental stage, and gut compartment in the gut bacterial communities of two Cerambycidae species (Coleoptera). J. Microbiol. 2017, 55, 21–30. [Google Scholar] [CrossRef]
- Mikaelyan, A.; Dietrich, C.; Köhler, T.; Poulsen, M.; Sillam-Dussès, D.; Brune, A. Diet is the primary determinant of bacterial community structure in the guts of higher termites. Mol. Ecol. 2015, 24, 5284–5295. [Google Scholar] [CrossRef]
- Priya, N.G.; Ojha, A.; Kajla, M.K.; Raj, A.; Rajagopal, R. Host plant induced variation in gut bacteria of Helicoverpa armigera. PLoS ONE 2017, 7, e30768. [Google Scholar] [CrossRef]
- Strano, C.P.; Malacrinò, A.; Campolo, O.; Palmeri, V. Influence of Host Plant on Thaumetopoea pityocampa Gut Bacterial Community. Microb. Ecol. 2018, 75, 487–494. [Google Scholar] [CrossRef]
- Liu, Y.J.; Shen, Z.J.; Yu, J.M.; Li, Z.; Liu, X.X.; Xu, H.L. Comparison of gut bacterial communities and their associations with host diets in four fruit borers. Pest Manag. Sci. 2020, 76, 1353–1362. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Ma, S.J. Occurrence dynamics of Locusta migratoria manilensis (Meyen) in China. Acta Entomol. Sin. 1958, 8, 1–40, 98–99. [Google Scholar] [CrossRef]
- Chen, Y.L. Characteristics, Causes and Ecological Management of Locust Disaster. Bull. Biol. 2000, 35, 1–5. [Google Scholar]
- Wu, W.W. Occurrence characteristics and control strategies of Oriental migratory locust. Hortic. Seed 2003, 23, 366. [Google Scholar]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.L. Diets Affects Hemolymphatic Metabolomics Hemolymph and Gut Microbial Diversity in the Silkworm, Bombyx mori. Master’s Thesis, Soochow University, Suzhou, China, 2017. [Google Scholar]
- Leite-Mondin, M.; DiLegge, M.J.; Manter, D.K.; Weir, T.L.; Silva-Filho, M.C.; Vivanco, J.M. The gut microbiota composition of Trichoplusia ni is altered by diet and may influence its polyphagous behavior. Sci. Rep. 2021, 11, 5786. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Z. Effect to Growth and Development of Locusta migratoria manilensis (Meyen) Reared on Different Kinds of Diets and Preliminary Identification of Intestinal Bacterium of Locusta migratoria manilensis (Meyen). Master’s Thesis, Shandong Agricultural University, Shandong, China, 2008. [Google Scholar]
- Lee, J.-H.; Lee, K.-A.; Lee, W.-J. Microbiota, Gut Physiology, and Insect Immunity; Academic Press: Cambridge, MA, USA, 2017; Volume 52, pp. 111–138. [Google Scholar] [CrossRef]
- Long, W.M. Molecular Ecological Study on the Intestinal Flora Structure of Grasshoppers. Master’s Thesis, Shanxi University, Shanxi, China, 2009. [Google Scholar]
- Shi, W.B.; Xie, S.X.; Chen, X.Y.; Sun, S.; Zhou, X.; Liu, L.T.; Gao, P.; Kyrpides, N.C.; No, E.-G.; Yuan, J.S. Comparative genomic analysis of the microbiome [corrected] of herbivorous insects reveals eco-environmental adaptations: Biotechnology applications. PLoS Genet. 2013, 9, e1003131. [Google Scholar] [CrossRef]
- Tinker, K.A.; Ottesen, E.A. The Core Gut Microbiome of the American Cockroach, Periplaneta americana, Is Stable and Resilient to Dietary Shifts. Appl. Environ. Microbiol. 2016, 82, 6603–6610. [Google Scholar] [CrossRef]
- Wu, X.L.; Xia, X.F.; Chen, J.H.; GURR, G.M.; You, M.S. Effects of different diets on the diversity of larval gut bacteria of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Acta Entomol. Sin. 2019, 62, 1172–1185. [Google Scholar] [CrossRef]
- Lv, F.; Liu, Y.S. The identification of dominant intestinal bacteriae of Clanis bilineata tsingtauica Mell larva. Chin. J. Microecol. 2009, 21, 435–439. [Google Scholar] [CrossRef]
- Jin, L.; Wang, H.C.; Wang, H.X.; Zhang, J.G.; Gang, Y.; Jin, H.Y. Bacterial Community in Midguts of Ectropic oblique Larvae by PCR-DGGE and 16SrRNA Gene Library Analysis. Jiangxi Sci. 2013, 31, 759–763, 829. [Google Scholar] [CrossRef]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef]
- Mao, Y.X.; Tan, R.R.; Wang, Y.P.; Chen, X.; Wang, H.J.; Huang, D.J.; Gong, Z.M. Analysis of the bacterial diversity in adults of Empoasca (Matsumurasca) onukii based on 16SrDNA sequences. Plant Prot. 2018, 44, 17–23, 48. [Google Scholar] [CrossRef]
- Genta, F.A.; Dillon, R.J.; Terra, W.R.; Ferreira, C. Potential role for gut microbiota in cell wall digestion and glucoside detoxification in Tenebrio molitor larvae. J. Insect Physiol. 2006, 52, 593–601. [Google Scholar] [CrossRef]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Sun, S.J.; Yang, X.L.; Cheng, J.; Wei, H.S.; Li, Z.; Michaud, J.P.; Liu, X.X. Variability of Gut Microbiota Across the Life Cycle of Grapholita molesta (Lepidoptera: Tortricidae). Front. Microbiol. 2020, 11, 1366. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.-S.; Song, S.-L.; Chua, K.-O.; Lim, P.-E. High Diversity of Bacterial Communities in Developmental Stages of Bactrocera carambolae (Insecta: Tephritidae) Revealed by Illumina MiSeq Sequencing of 16S rRNA Gene. Curr. Microbiol. 2017, 74, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.F.; Zhang, X.Y.; Chen, Z.S.; Wang, Z.; Lu, Y.Y.; Cheng, D.F. The Divergence in Bacterial Components Associated with Bactrocera dorsalis across Developmental Stages. Front. Microbiol. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Joynson, R.; Swamy, A.; Bou, P.A.; Chapuis, A.; Ferry, N. Characterization of cellulolytic activity in the gut of the terrestrial land slug Arion ater: Biochemical identification of targets for intensive study. Comp. Biochem. Physiol. Part. B 2014, 177, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X. Biological Nitrogen Fixation and Mechanism of Bite-Degrading Polystyrene Insects. Master’s Thesis, Wuhan University of Technology, Wuhan, China, 2019. [Google Scholar]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Pasquini, M.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Loreto, N.; et al. The bacterial biota of laboratory-reared edible mealworms (Tenebrio molitor L.): From feed to frass. Int. J. Food Microbiol. 2018, 272, 49–60. [Google Scholar] [CrossRef]
- Wei, D.F.; Wang, X.J.; Yang, J.; Geng, Y.X.; Chen, M. Analysis of the diversity and difference of intestinal bacteria in larvae Hyphantria cunea Drury (Lepidoptera: Arctiidae) on different diets. J. Environ. Entomol. 2017, 39, 515–524. [Google Scholar] [CrossRef]
- He, X.H.; Liu, Y.S.; Zheng, J.F. Effects of different feeding plants on the intestinal bacteria of Locusta migratoria manilensis. Chin. J. Microecol. 2010, 22, 492–497. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).