
Social Rank or Social Bonds: Which one Facilitates Coalition Formation in Male Tibetan Macaques?

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Subjects
2.2. Data Collection
2.3. Dominance Hierarchy of Males
2.4. Male Social Relations
2.5. Male Coalition Behavior
2.6. Statistical Analyses
2.7. Ethics Statement
3. Results
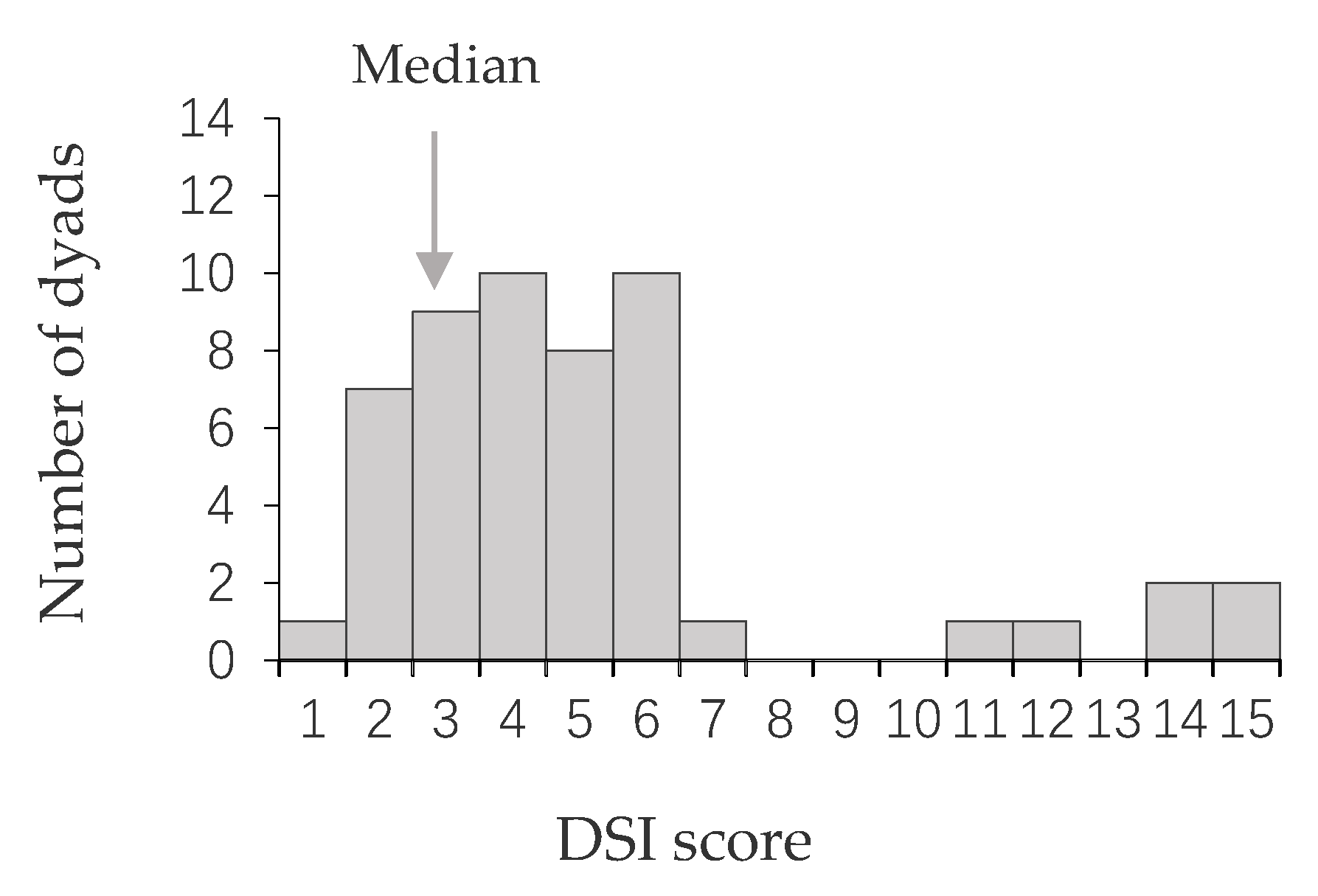
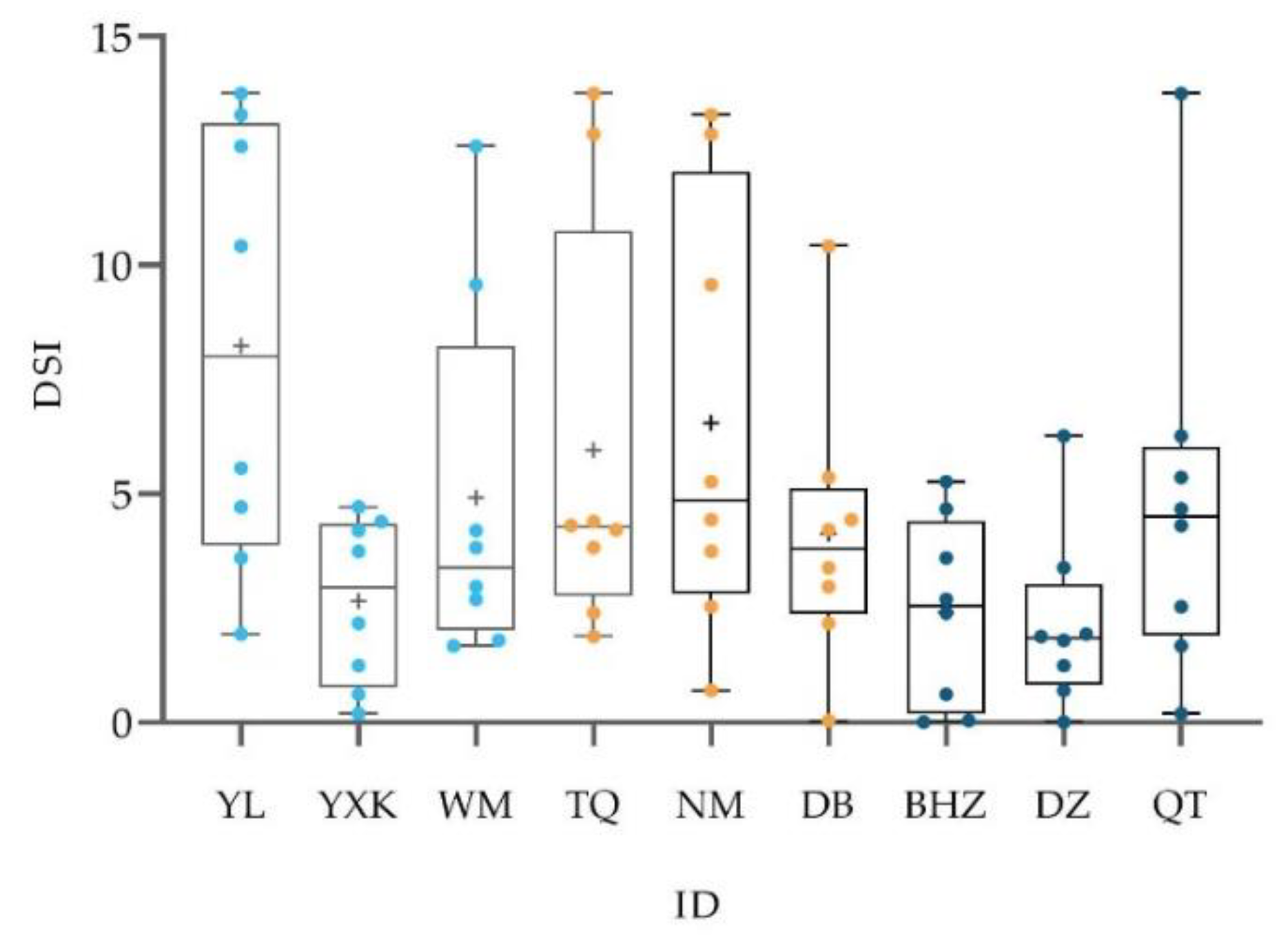
3.1. Male Social Relations: Strength and Stability
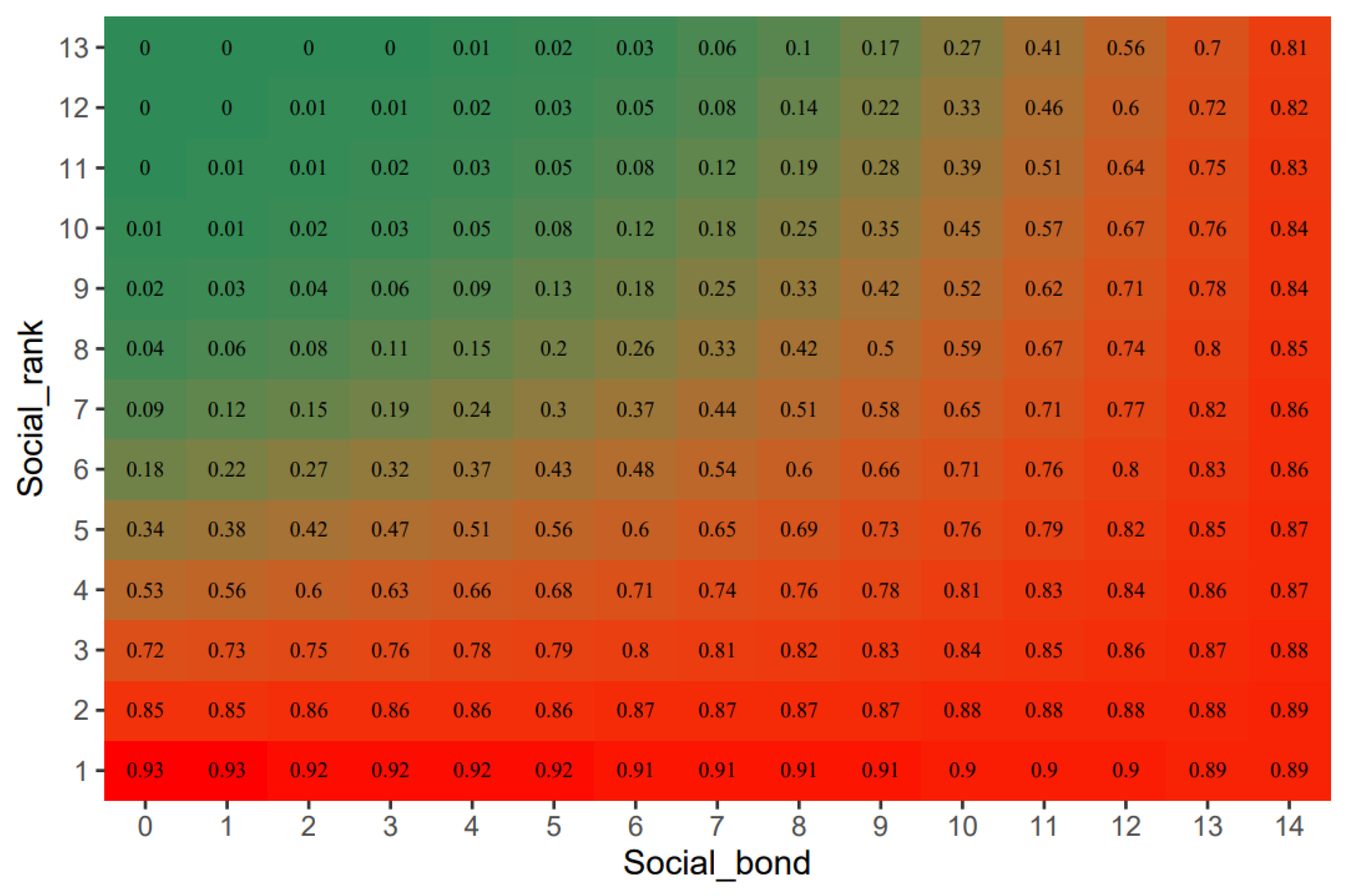
3.2. Coalitionary Behavior
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Radespiel, U.; Ehresmann, P.; Zimmermann, E. Contest versus scramble competition for mates: The composition and spatial structure of a population of gray mouse lemurs (Microcebus murinus) in North-west Madagascar. Primates 2001, 42, 207. [Google Scholar] [CrossRef]
- Cant, M.A.; Otali, E.; Mwanguhya, F. Fighting and Mating Between Groups in a Cooperatively Breeding Mammal, the Banded Mongoose. Ethology 2002, 108, 541–555. [Google Scholar] [CrossRef]
- Winkelmann, K.; Genner, M.J.; Takahashi, T.; Ruber, L. Competition-driven speciation in cichlid fish. Nat. Commun. 2014, 5, 3. [Google Scholar] [CrossRef]
- Luehrs, M.-L.; Kappeler, P.M. Polyandrous mating in treetops: How male competition and female choice interact to determine an unusual carnivore mating system. Behav. Ecol. Sociobiol. 2014, 68, 879–889. [Google Scholar] [CrossRef]
- Girard-Buttoz, C.; Heistermann, M.; Rahmi, E.; Agil, M.; Fauzan, P.A.; Engelhardt, A. Costs of and Investment in Mate-Guarding in Wild Long-Tailed Macaques (Macaca fascicularis): Influences of Female Characteristics and Male-Female Social Bonds. Int. J. Primatol. 2014, 35, 701–724. [Google Scholar] [CrossRef]
- Cassidy, K.A.; McIntyre, R.T. Do gray wolves (Canis lupus) support pack mates during aggressive inter-pack interactions? Anim. Cogn. 2016, 19, 939–947. [Google Scholar] [CrossRef]
- Port, M.; Schulke, O.; Ostner, J. Reproductive tolerance in male primates: Old paradigms and new evidence. Evol. Anthropol. 2018, 27, 107–120. [Google Scholar] [CrossRef]
- Young, C.; Schülke, O.; Ostner, J. How males form coalitions against group rivals and the Pandit/van Schaik coalition model. Behaviour 2014, 151, 907–934. [Google Scholar] [CrossRef]
- Ostner, J.; Schuelke, O. The evolution of social bonds in primate males. Behaviour 2014, 151, 871–906. [Google Scholar] [CrossRef]
- Hinde, R.A. Primate Social Relationships: An Integrated Approach; Blackwell: Oxford, UK, 1983. [Google Scholar]
- Berghaenel, A.; Ostner, J.; Schroeder, U.; Schuelke, O. Social bonds predict future cooperation in male Barbary macaques, Macaca sylvanus. Anim. Behav. 2011, 81, 1109–1116. [Google Scholar] [CrossRef]
- Sandel, A.A.; Reddy, R.B.; Mitani, J.C. Adolescent male chimpanzees do not form a dominance hierarchy with their peers. Primates 2017, 58, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Kawazoe, T.; Sosa, S. Social networks predict immigration success in wild Japanese macaques. Primates 2019, 60, 213–222. [Google Scholar] [CrossRef]
- Duffy, K.G.; Wrangham, R.W.; Silk, J.B. Male chimpanzees exchange political support for mating opportunities. Curr. Biol. 2007, 17, R586–R587. [Google Scholar] [CrossRef]
- Schuelke, O.; Bhagavatula, J.; Vigilant, L.; Ostner, J. Social Bonds Enhance Reproductive Success in Male Macaques. Curr. Biol. 2010, 20, 2207–2210. [Google Scholar] [CrossRef]
- De Moor, D.; Roos, C.; Ostner, J.; Schulke, O. Bonds of bros and brothers: Kinship and social bonding in postdispersal male macaques. Mol. Ecol. 2020, 29, 3346–3360. [Google Scholar] [CrossRef]
- Koykka, C.; Wild, G. Concessions, lifetime fitness consequences, and the evolution of coalitionary behavior. Mol. Ecol. 2017, 28, 20–30. [Google Scholar] [CrossRef]
- Broom, M.; Koenig, A.; Borries, C. Variation in dominance hierarchies among group-living animals: Modeling stability and the likelihood of coalitions. Mol. Ecol. 2009, 20, 844–855. [Google Scholar] [CrossRef]
- Reddy, R.B.; Mitani, J.C. Adolescent and young adult male chimpanzees form affiliative, yet aggressive, relationships with females. J. Hum. Evol. 2020, 144, 102813. [Google Scholar] [CrossRef]
- Schoof, V.A.M.; Jack, K.M. Male social bonds: Strength and quality among co-resident white-faced capuchin monkeys (Cebus capucinus). Behaviour 2014, 151, 963–992. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- Berman, C.M.; Ionica, C.S.; Li, J.H. Dominance style among Macaca thibetana on Mt. Huangshan, China. Int. J. Primatol. 2004, 25, 1283–1312. [Google Scholar] [CrossRef]
- Li, J.H. The Tibetan Macaque Society: A Field Study; Anhui University Press: Hefei, China, 1999. [Google Scholar]
- Gammell, M.P.; De Vries, H.; Jennings, D.J.; Carlin, C.M.; Hayden, T.J. David’s score: A more appropriate dominance ranking method than Clutton-Brock et al.’s index. Anim. Behav. 2003, 66, 601–605. [Google Scholar] [CrossRef]
- Miss, F.M.; Sadoughi, B.; Meunier, H.; Burkart, J.M. Individual differences in co-representation in three monkey species (Callithrix jacchus, Sapajus apella and Macaca tonkeana) in the joint Simon task: The role of social factors and inhibitory control. Anim. Cogn. 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Silk, J.; Cheney, D.; Seyfarth, R. A Practical Guide to the Study of Social Relationships. Evol. Anthropol. 2013, 22, 213–225. [Google Scholar] [CrossRef]
- Bissonnette, A.; de Vries, H.; van Schaik, C.P. Coalitions in male Barbary macaques, Macaca sylvanus: Strength, success and rules of thumb. Anim. Behav. 2009, 78, 329–335. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bray, J.; Feldblum, J.T.; Gilby, I.C. Social bonds predict dominance trajectories in adult male chimpanzees. Anim. Behav. 2021, 179, 339–354. [Google Scholar] [CrossRef]
- Gilby, I.C.; Brent, L.J.N.; Wroblewski, E.E.; Rudicell, R.S.; Hahn, B.H.; Goodall, J.; Pusey, A.E. Fitness benefits of coalitionary aggression in male chimpanzees. Mol. Ecol. Sociobiol. 2013, 67, 373–381. [Google Scholar] [CrossRef]
- Bray, J.; Gilby, I.C. Social relationships among adult male chimpanzees (Pan troglodytes schweinfurthii): Variation in the strength and quality of social bonds. Mol. Ecol. Sociobiol. 2020, 74, 112. [Google Scholar] [CrossRef]
- St-Pierre, A.; Larose, K.; Dubois, F. Long-term social bonds promote cooperation in the iterated Prisoner’s Dilemma. Proc. R. Soc. B-Biol. Sci. 2009, 276, 4223–4228. [Google Scholar] [CrossRef] [Green Version]
- Chapais, B. Alliances as means of competition in primates: Evolutionary, developmental and cognitive aspects. Yearb. Phys. Anthropol. 1995, 38, 115–136. [Google Scholar] [CrossRef]
- Perry, S.; Barrett, H.C.; Manson, J.H. White-faced capuchin monkeys show triadic awareness in their choice of allies. Anim. Behav. 2004, 67, 165–170. [Google Scholar] [CrossRef]
- Neumann, C.; Kulik, L.; Agil, M.; Engelhardt, A.; Widdig, A. Temporal dynamics and fitness consequences of coalition formation in male primates. Proc. R. Soc. B Biol. Sci. 2022, 289, 20212626. [Google Scholar] [CrossRef]
- Carne, C.; Wiper, S.; Semple, S. Reciprocation and interchange of grooming, agonistic support, feeding tolerance, and aggression in semi-free-ranging Barbary macaques. Am. J. Primatol. 2011, 73, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Gerber, L.; Connor, R.C.; King, S.L.; Allen, S.J.; Wittwer, S.; Bizzozzero, M.R.; Friedman, W.R.; Kalberer, S.; Sherwin, W.B.; Wild, S.; et al. Affiliation history and age similarity predict alliance formation in adult male bottlenose dolphins. Mol. Ecol. 2020, 31, 361–370. [Google Scholar] [CrossRef]
- Gerber, L.; Wittwer, S.; Allen, S.J.; Holmes, K.G.; King, S.L.; Sherwin, W.B.; Wild, S.; Willems, E.P.; Connor, R.C.; Krutzen, M. Cooperative partner choice in multi-level male dolphin alliances. Sci. Rep. 2021, 11, 6901. [Google Scholar] [CrossRef]
- Widdig, A.; Streich, W.J.; Tembrock, G. Coalition formation among male Barbary macaques (Macaca sylvanus). Am. J. Primatol. 2000, 50, 37–51. [Google Scholar] [CrossRef]
- Engh, A.L.; Siebert, E.R.; Greenberg, D.A.; Holekamp, K.E. Patterns of alliance formation and postconflict aggression indicate spotted hyaenas recognize third-party relationships. Anim. Behav. 2005, 69, 209–217. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Behavior | Description |
|---|---|
| Proximity | Two or more individuals are sitting or lying within 5 m of one another. |
| Stare | An individual looks directly at another individual with its eyes wide open and with its shoulders raised for about 3–5 s. |
| Ground slap | One individual support with one hand on the ground, the other flaps the ground, and stares at another individual, then it looks down. |
| Chase | An individual stares at the recipient and rushes at him/her at great speed. The recipient typically flees. |
| Seizing | One individual grabs the hair of the body, face, or neck of another one, or it may simply grab its ears. Shake it back and forth a few times before letting go. |
| Bite | The performer grabs the receiver tightly, preventing him/her from fleeing, and bites the recipient vigorously. |
| Avoid | The performer turns its body away from the attacker as if preparing to flee while displaying a ‘horrified’ facial expression towards the attacker. |
| Flee | The target of an attack will run in the opposite direction from the attacker. |
| Agonistic support | The behavior of a social animal in which an individual participates in an ongoing struggle to support one party and attack the other. |
| ID | Social Rank (DS Value) | Age Class * | Residence Time (Year) |
|---|---|---|---|
| YL | 1 (22.63) | 1 | 0.4 |
| YXK | 2 (22.52) | 1 | 9 |
| WM | 3 (14.08) | 3 | 2 |
| TQ | 4 (4.90) | 2 | 2 |
| NM | 5 (2.90) | 2 | 0.4 |
| DB | 6 (−1.20) | 1 | 0.4 |
| BHZ | 7 (−17.17) | 2 | 0.3 |
| DZ | 8 (−20.08) | 1 | 0.2 |
| QT | 9 (−29.02) | 1 | 0.4 |
| Model | Predictive Variables | Response Variables |
|---|---|---|
| Model 1 | DSI score | A male from individuals in conflict was selected as a coalition partner (Yes/No) |
| High rank (Yes/No) | ||
| The rank difference of individuals in conflict | ||
| Model 2 | DSI score of dyad I-J | Number of coalitions of dyad I-J |
| The residence time of J | ||
| Age class of J | ||
| Model 3 | The interaction between social rank and social bond | The recruiter is selected as a coalition partner (Yes/No) |
| Predictors | Estimate | SE | Z Value | p Values |
|---|---|---|---|---|
| DSI score | 0.19 | 0.09 | 2.02 | p = 0.044 |
| High rank (Yes) | 4.62 | 0.76 | 6.06 | p < 0.001 |
| Predictors | Estimate | SE | Z Values | p Values |
|---|---|---|---|---|
| DSI | 0.145 | 0.027 | 5.223 | p < 0.001 |
| Residence time | 0.119 | 0.044 | 2.710 | p = 0.007 |
| Age class | 0.051 | 0.181 | 0.287 | p = 0.774 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tai, Y.-M.; Chen, M.-M.; Zhang, Y.-H.; Ma, A.-X.; Wang, H.; Wang, X. Social Rank or Social Bonds: Which one Facilitates Coalition Formation in Male Tibetan Macaques? Biology 2022, 11, 1269. https://doi.org/10.3390/biology11091269
Tai Y-M, Chen M-M, Zhang Y-H, Ma A-X, Wang H, Wang X. Social Rank or Social Bonds: Which one Facilitates Coalition Formation in Male Tibetan Macaques? Biology. 2022; 11(9):1269. https://doi.org/10.3390/biology11091269
Chicago/Turabian StyleTai, Yi-Mei, Meng-Meng Chen, Yu-Heng Zhang, An-Xiang Ma, Huan Wang, and Xi Wang. 2022. "Social Rank or Social Bonds: Which one Facilitates Coalition Formation in Male Tibetan Macaques?" Biology 11, no. 9: 1269. https://doi.org/10.3390/biology11091269
APA StyleTai, Y.-M., Chen, M.-M., Zhang, Y.-H., Ma, A.-X., Wang, H., & Wang, X. (2022). Social Rank or Social Bonds: Which one Facilitates Coalition Formation in Male Tibetan Macaques? Biology, 11(9), 1269. https://doi.org/10.3390/biology11091269