
Perturbation by Antimicrobial Bacteria of the Epidermal Bacterial Flora of Rainbow Trout in Flow-Through Aquaculture

 and
and 
Abstract
:Simple Summary
Abstract
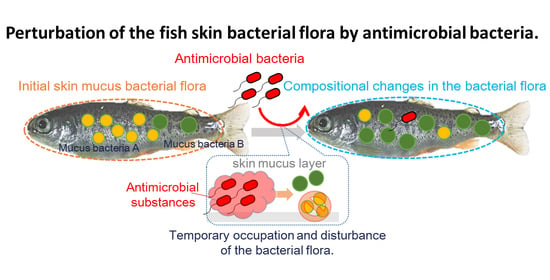
1. Introduction
2. Materials and Methods
2.1. Manipulation of Bacterial Strains, Cultivation of Fish Skin Bacteria, and Screening
2.2. Next-Generation Sequencing of 16S rDNA Amplicon Libraries
2.3. In Silico Analysis Based on 16S rDNA Amplicon Sequencing
2.4. Maintenance and Handling of Rainbow Trout in Fish Farm and Laboratory Aquarium
3. Results
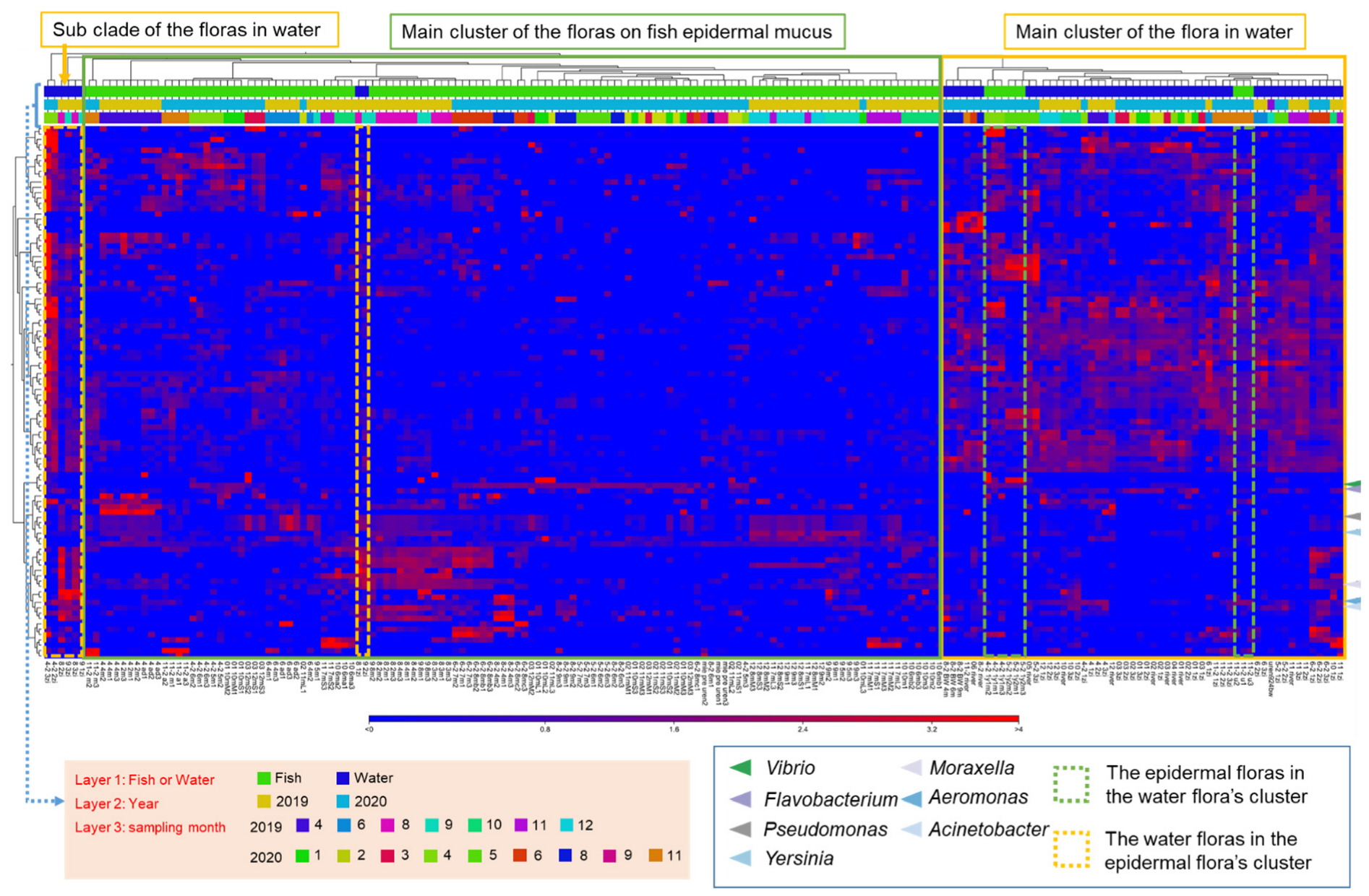
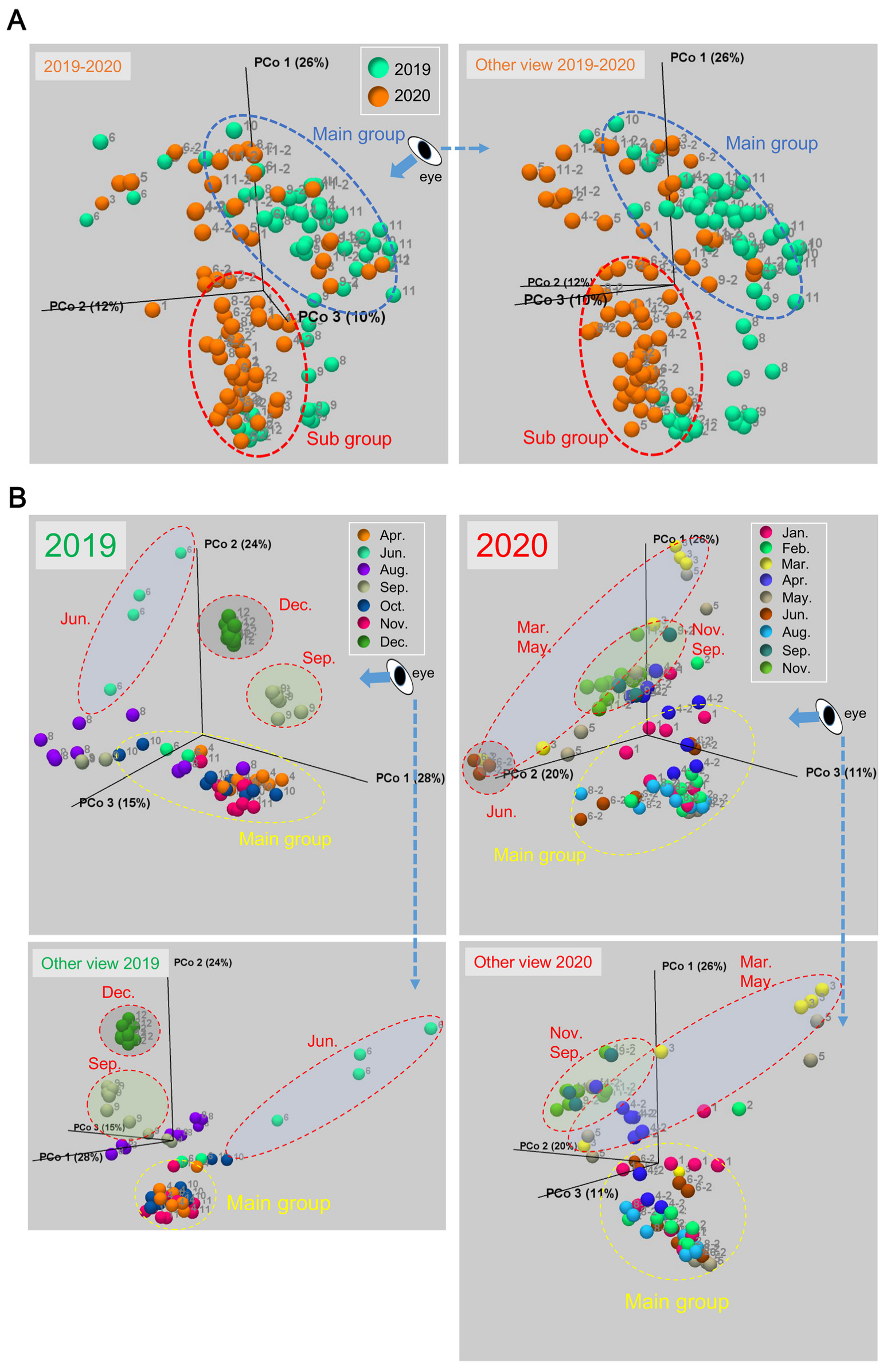
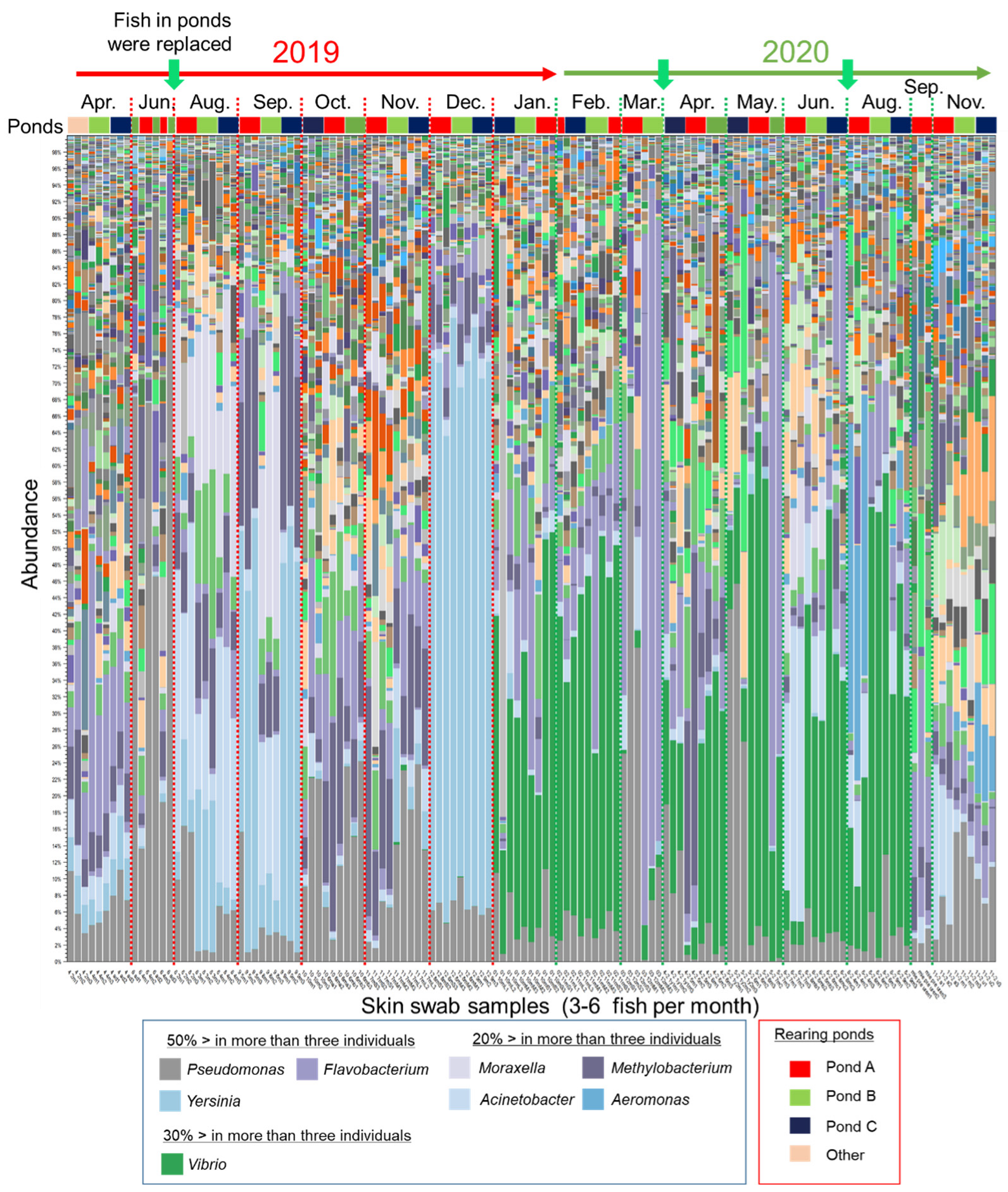
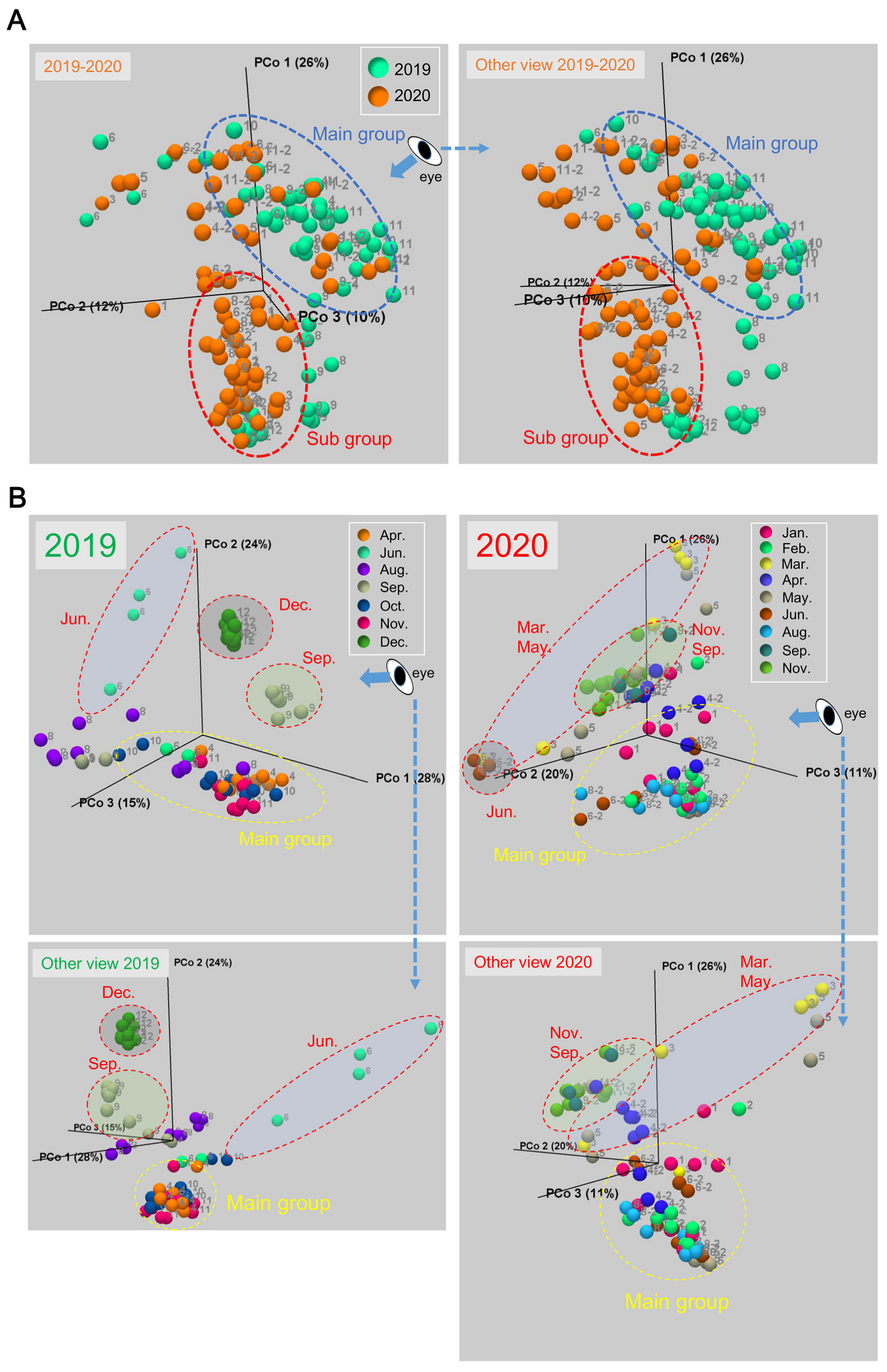
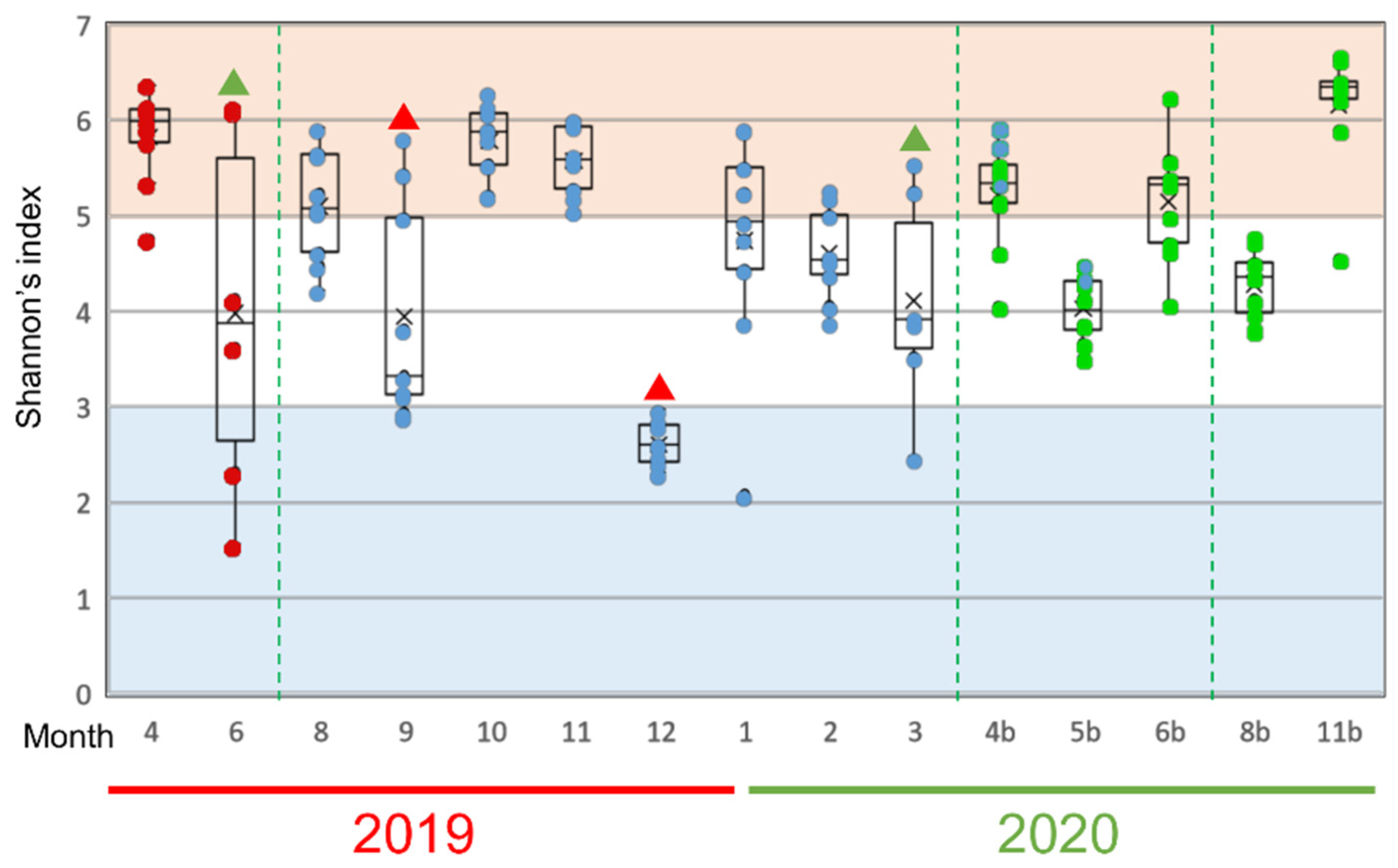
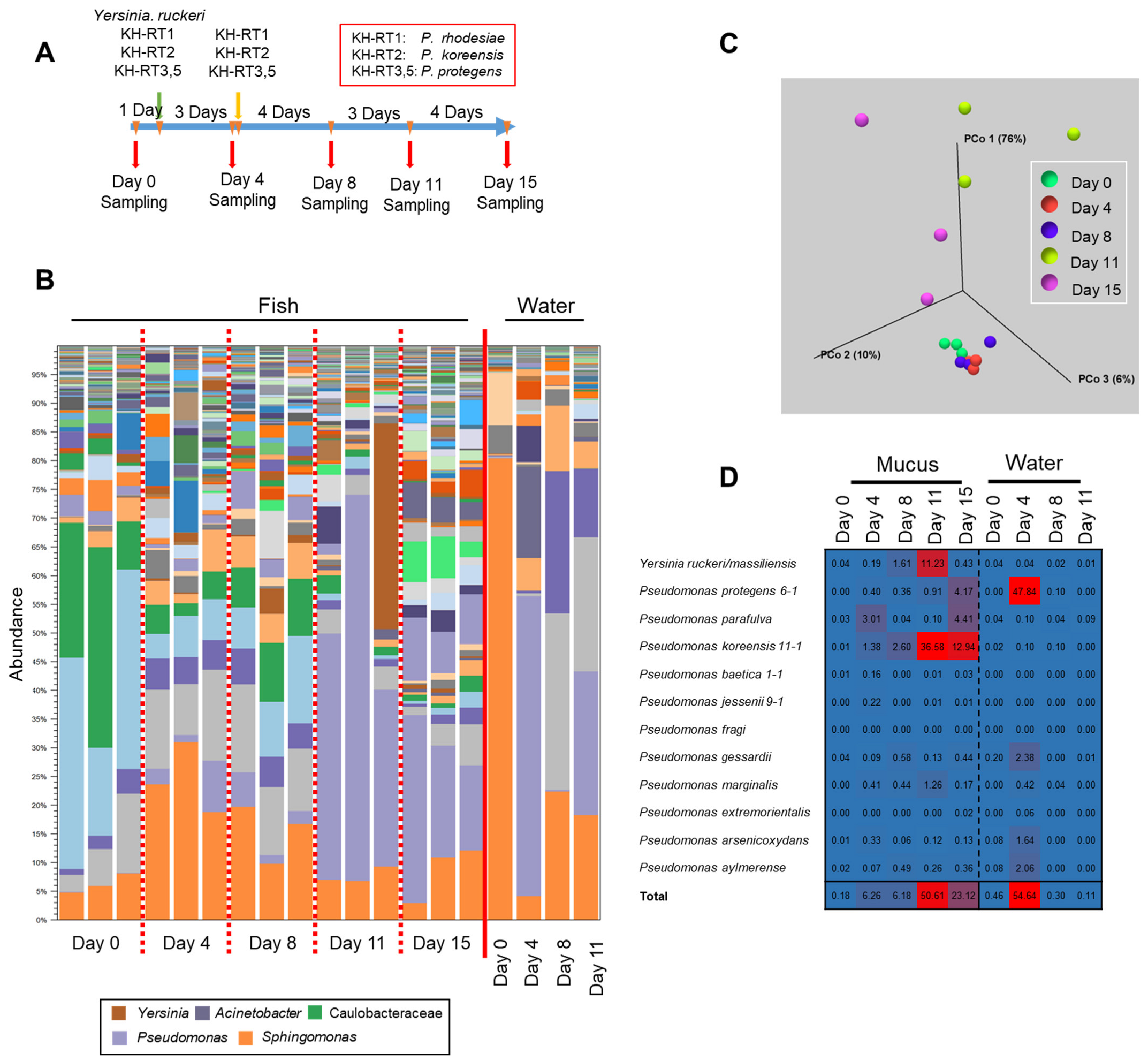
3.1. Changes of the Epidermal Mucus Microflora of Farmed Rainbow Trout in a Flow-Through System
3.2. Changes in Abundances of Epidermal Mucus Bacteria Independent of the Abundance of Bacteria in Water and Other Conditions of Sampling
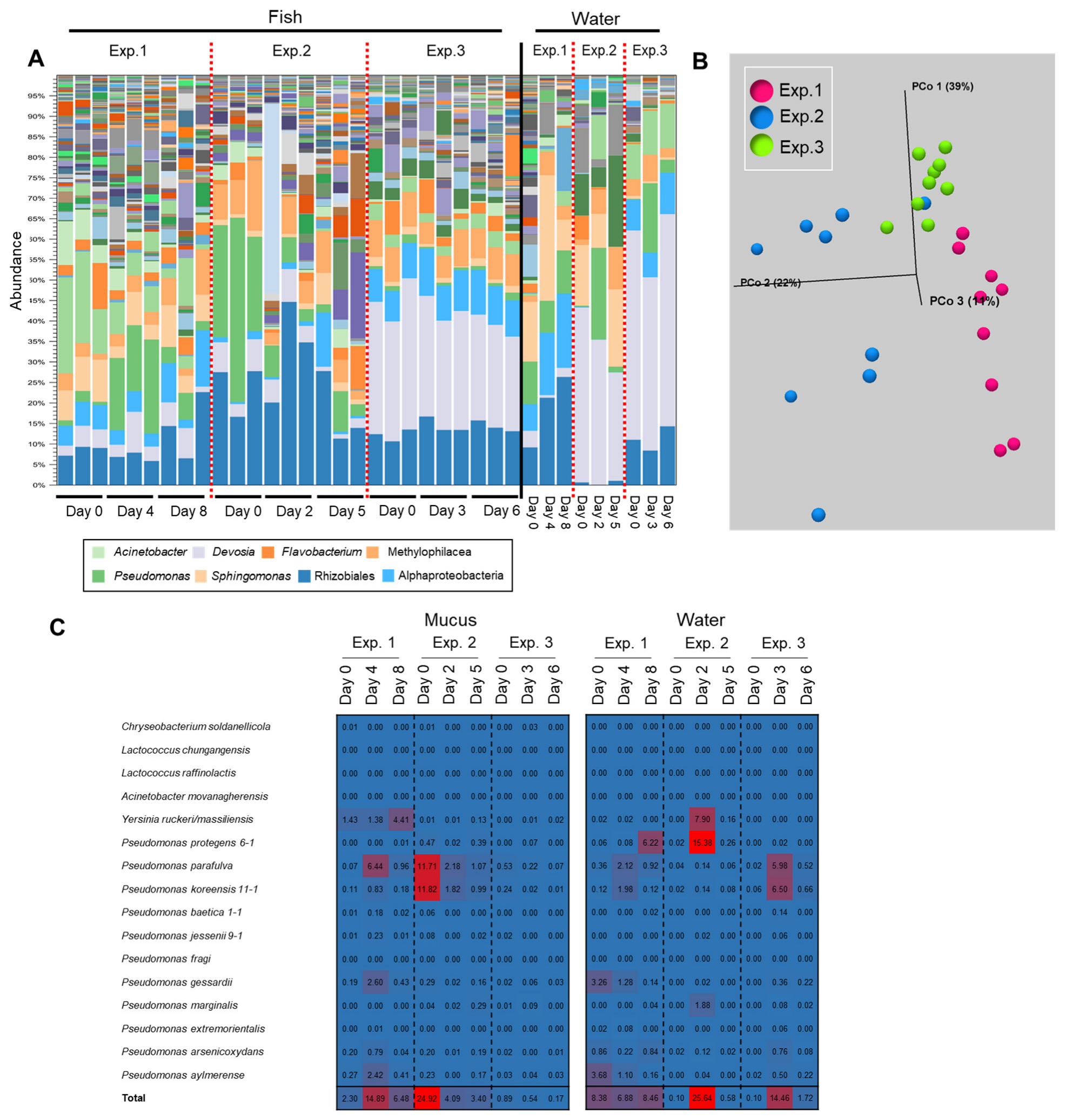
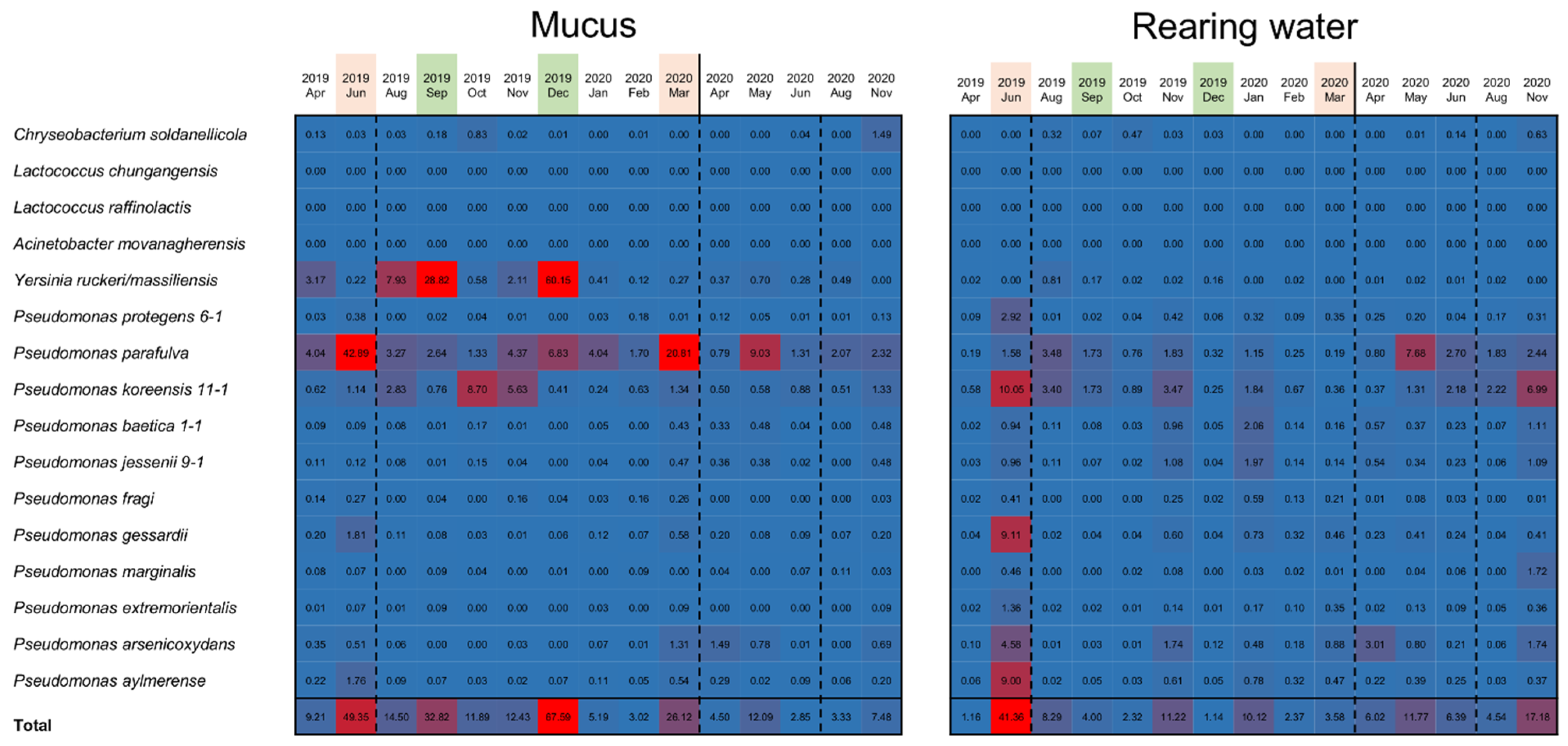
3.3. Specific Bacterial Species Involved in Fluctuations of the Composition of Skin Bacterial Flora in Farmed Rainbow Trout
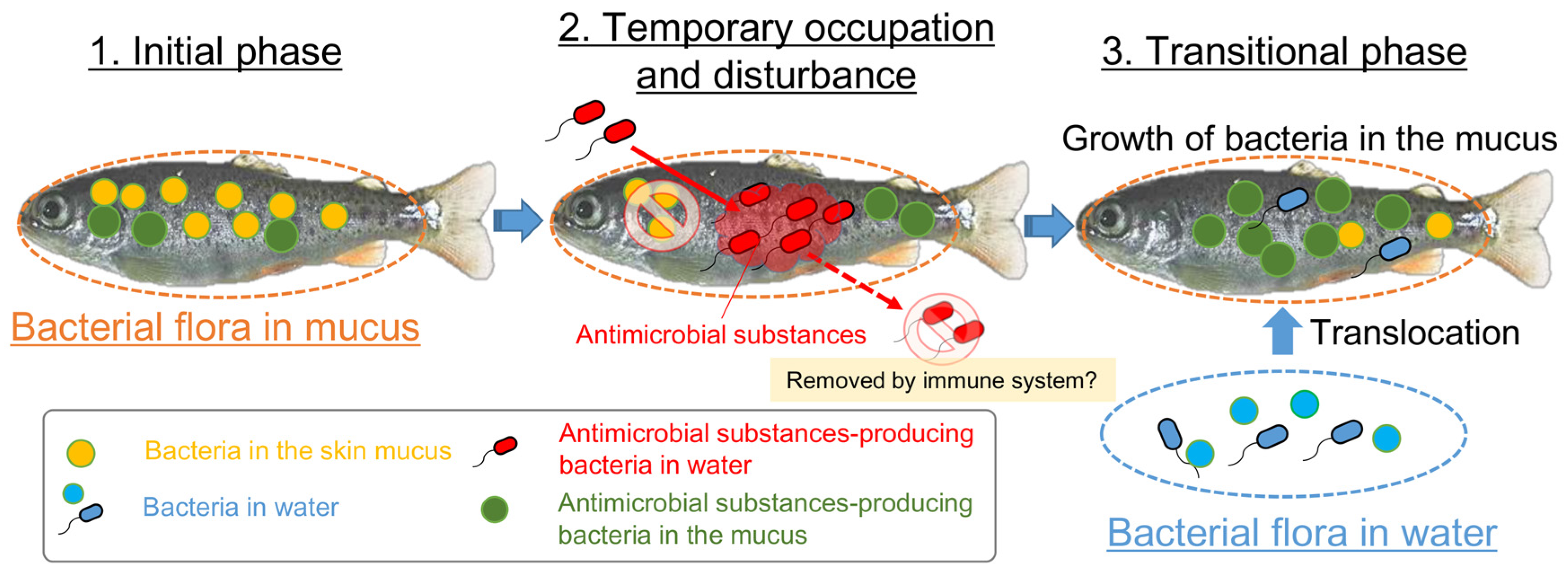
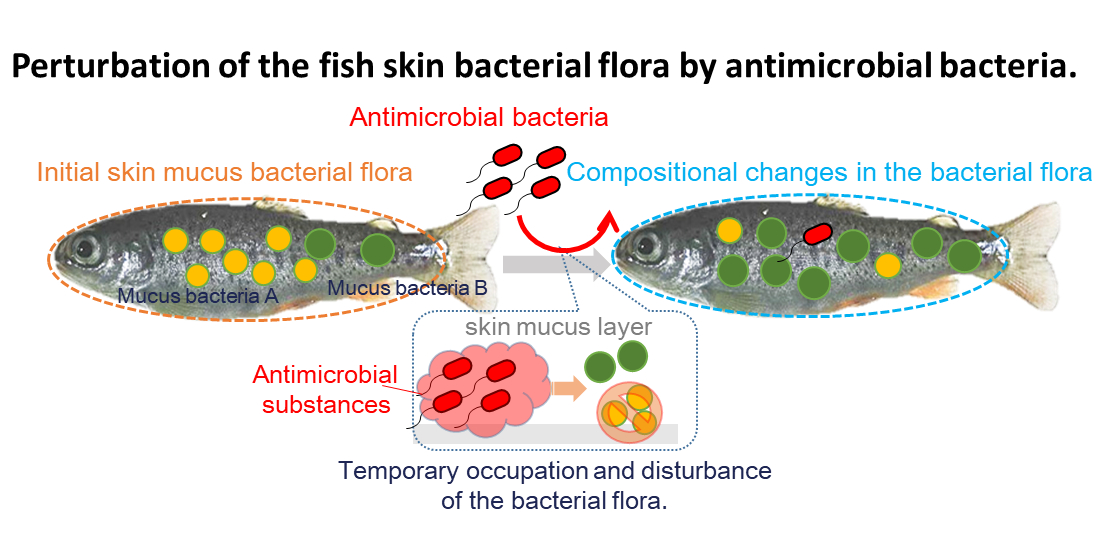
3.4. Involvement of Specific Antimicrobial Bacteria in Perturbation of the Fish Epidermal Mucus Microflora
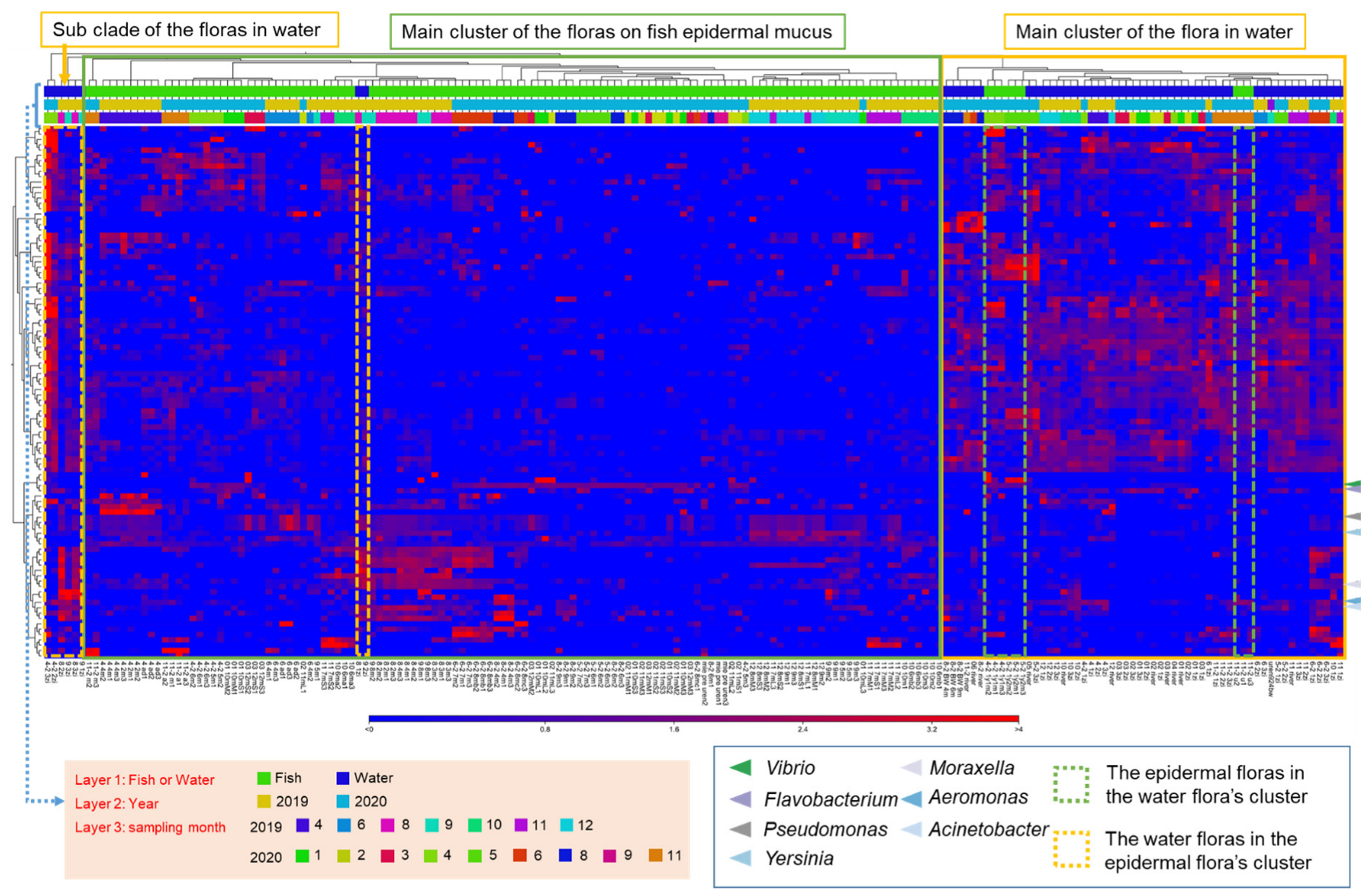
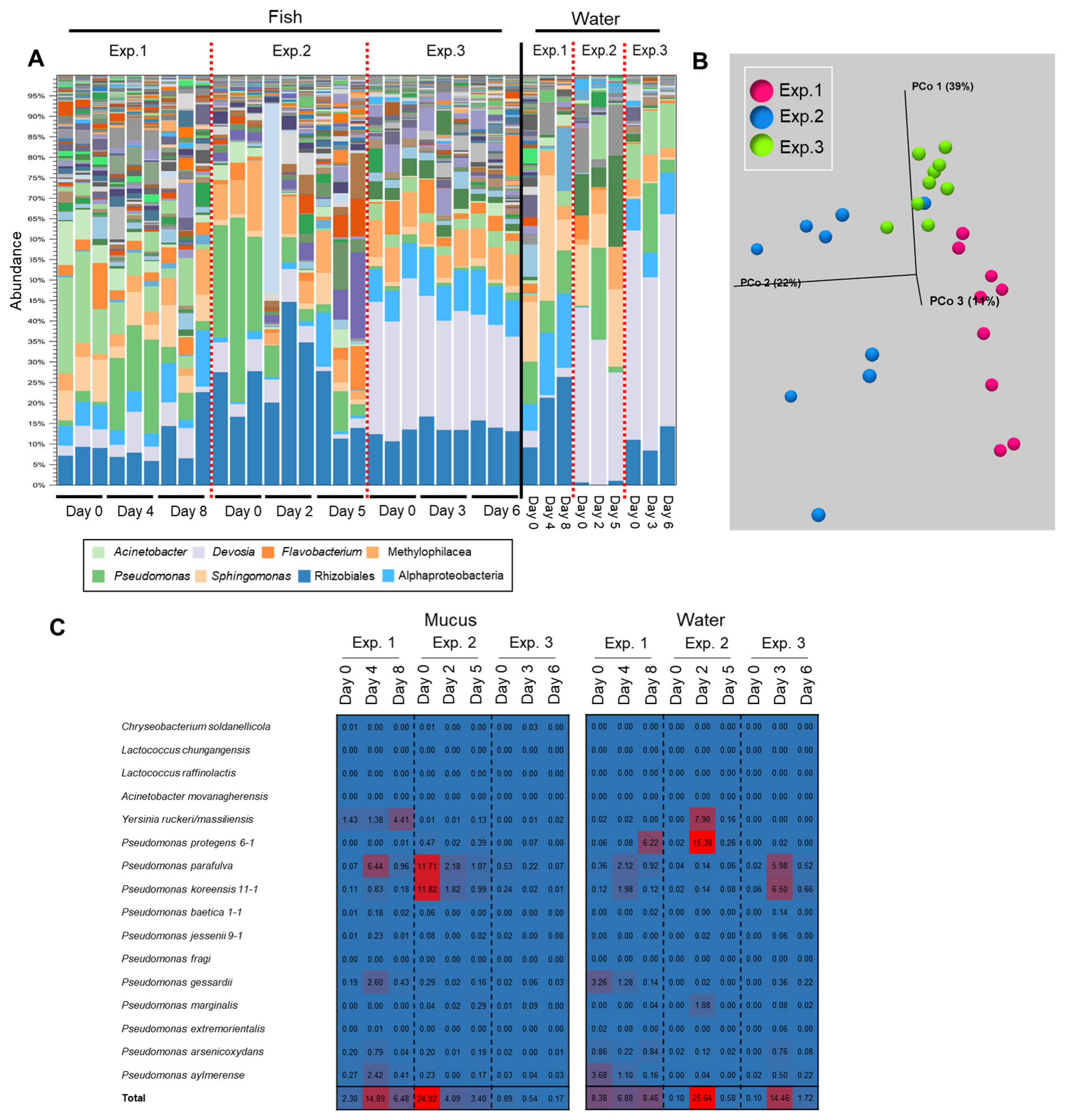
3.5. Transition of the Epidermal Mucus Bacterial Flora in a Stable Environment Was Linked to Antimicrobial Bacteria
3.6. Transition of the Epidermal Mucus Bacterial Flora of Rainbow Trout Owing to Antimicrobial Bacteria in the Water
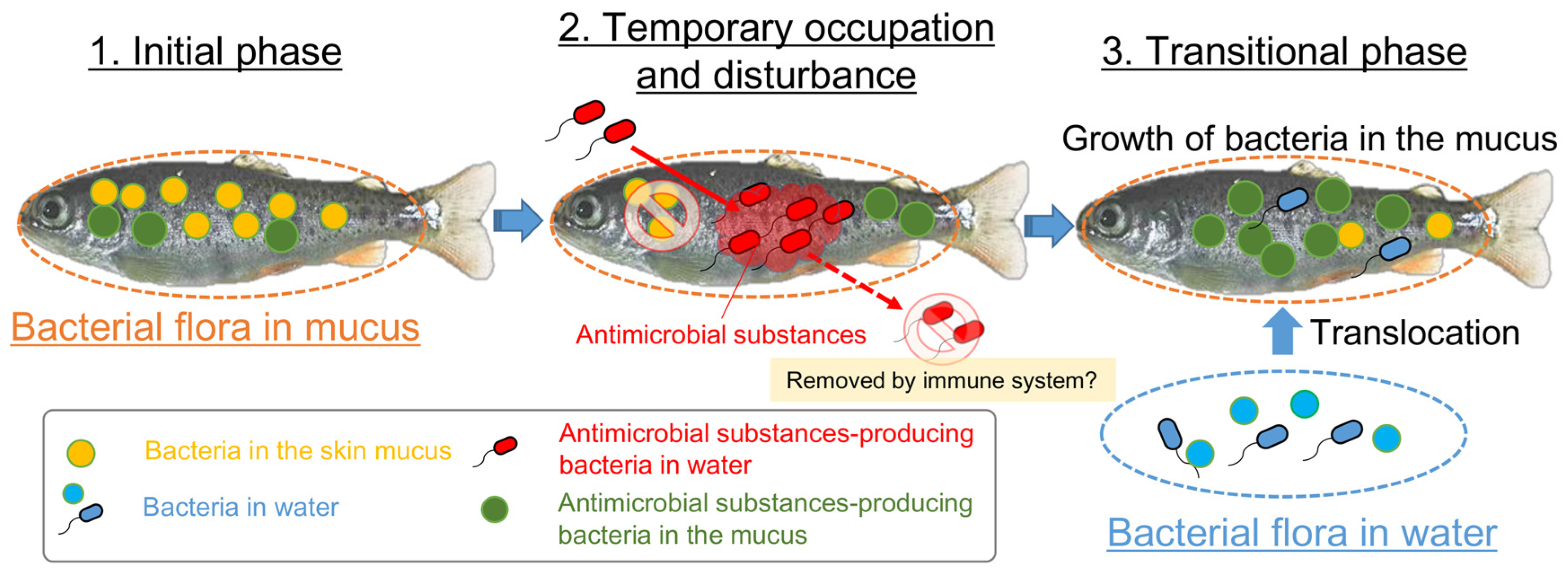
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brinchmann, M.F. Immune relevant molecules identified in the skin mucus of fish using -omics technologies. Mol. Biosyst. 2016, 12, 2056–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wu, H.; Liu, Q.; Wang, Q.; Xiao, J.; Zhang, Y. Skin-injured zebrafish, Danio rerio, are more susceptible to Vibrio anguillarum infection. J. World Aquac. Soc. 2015, 46, 301–310. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, S.; Knapp, R.A.; Sparagon, W.; Swei, A.; Vredenburg, V.T. Reduced skin bacterial diversity correlates with increased pathogen infection intensity in an endangered amphibian host. Mol. Ecol. 2019, 28, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ding, L.; Yu, Y.; Kong, W.; Yin, Y.; Huang, Z.; Zhang, X.; Xu, Z. The change of teleost skin commensal microbiota is associated with skin mucosal transcriptomic responses during parasitic infection by Ichthyophthirius multifillis. Front. Immunol. 2018, 9, 2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutin, S.; Bernatchez, L.; Audet, C.; Derome, N. Network analysis highlights complex interactions between pathogen, host and commensal microbiota. PLoS ONE 2013, 8, e84772. [Google Scholar] [CrossRef]
- Boutin, S.; Bernatchez, L.; Audet, C.; Derome, N. Antagonistic effect of indigenous skin bacteria of brook charr (Salvelinus fontinalis) against Flavobacterium columnare and F. psychrophilum. Vet. Microbiol. 2012, 155, 355–361. [Google Scholar] [CrossRef]
- Legrand, T.; Catalano, S.R.; Wos-Oxley, M.L.; Stephens, F.; Landos, M.; Bansemer, M.S.; Stone, D.A.J.; Qin, J.G.; Oxley, A.P.A. The inner workings of the outer surface: Skin and gill microbiota as indicators of changing gut health in yellowtail kingfish. Front. Microbiol. 2018, 8, 2664. [Google Scholar] [CrossRef]
- Lowrey, L.; Woodhams, D.C.; Tacchi, L.; Salinas, I. Topographical mapping of the rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community with antifungal properties in the skin. Appl. Environ. Microbiol. 2015, 81, 6915–6925. [Google Scholar] [CrossRef] [Green Version]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome composition and function in aquatic vertebrates: Small organisms making big impacts on aquatic animal health. Front. Microbiol. 2021, 12, 567408. [Google Scholar] [CrossRef]
- Chiarello, M.; Auguet, J.C.; Bettarel, Y.; Bouvier, C.; Claverie, T.; Graham, N.A.J.; Rieuvilleneuve, F.; Sucre, E.; Bouvier, T.; Villeger, S. Skin microbiome of coral reef fish is highly variable and driven by host phylogeny and diet. Microbiome 2018, 6, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarello, M.; Villeger, S.; Bouvier, C.; Bettarel, Y.; Bouvier, T. High diversity of skin-associated bacterial communities of marine fishes is promoted by their high variability among body parts, individuals and species. FEMS Microbiol. Ecol. 2015, 91, fiv061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarello, M.; Paz-Vinas, I.; Veyssière, C.; Santoul, F.; Loot, G.; Ferriol, J.; Boulêtreau, S. Environmental conditions and neutral processes shape the skin microbiome of european catfish (Silurus glanis) populations of southwestern france. Environ. Microbiol. Rep. 2019, 11, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.; Danilowicz, B.S.; Meijer, W.G. The diversity of bacterial communities associated with atlantic cod Gadus morhua. Microb. Ecol. 2008, 55, 425–434. [Google Scholar] [CrossRef]
- Sylvain, F.E.; Cheaib, B.; Llewellyn, M.; Gabriel Correia, T.; Barros Fagundes, D.; Luis Val, A.; Derome, N. Ph drop impacts differentially skin and gut microbiota of the amazonian fish tambaqui (Colossoma macropomum). Sci. Rep. 2016, 6, 32032. [Google Scholar] [CrossRef] [Green Version]
- Carda-Diéguez, M.; Ghai, R.; Rodríguez-Valera, F.; Amaro, C. Wild eel microbiome reveals that skin mucus of fish could be a natural niche for aquatic mucosal pathogen evolution. Microbiome 2017, 5, 162. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, V.T.; Smith, K.F.; Melvin, D.W.; Amaral-Zettler, L.A. Community assembly of a euryhaline fish microbiome during salinity acclimation. Mol. Ecol. 2015, 24, 2537–2550. [Google Scholar] [CrossRef]
- Krotman, Y.; Yergaliyev, T.M.; Alexander Shani, R.; Avrahami, Y.; Szitenberg, A. Dissecting the factors shaping fish skin microbiomes in a heterogeneous inland water system. Microbiome 2020, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Minniti, G.; Hagen, L.H.; Porcellato, D.; Jørgensen, S.M.; Pope, P.B.; Vaaje-Kolstad, G. The skin-mucus microbial community of farmed atlantic salmon (Salmo salar). Front. Microbiol. 2017, 8, 2043. [Google Scholar] [CrossRef] [Green Version]
- Uren Webster, T.M.; Consuegra, S.; Hitchings, M.; Garcia de Leaniz, C. Interpopulation variation in the atlantic salmon microbiome reflects environmental and genetic diversity. Appl. Environ. Microbiol. 2018, 84, e00691-18. [Google Scholar] [CrossRef] [Green Version]
- Nakatani, H.; Hori, K. Establishing a percutaneous infection model using zebrafish and a salmon pathogen. Biology 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Menanteau-Ledouble, S.; Saleh, M.; El-Matbouli, M. Yersinia ruckeri, the causative agent of enteric redmouth disease in fish. Vet. Res. 2015, 46, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, E.; Borsodi, A.; Felföldi, T.; Vajna, B.; Sipos, R.; Márialigeti, K. Practical Microbiology: Based on the Hungarian Practical Notes Entitled "Mikrobiológiai Laboratóriumi Gyakorlatok"; Eötvös Loránd University: Budapest, Hungary, 2013. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- López, J.R.; Lorenzo, L.; Marcelino-Pozuelo, C.; Marin-Arjona, M.C.; Navas, J.I. Pseudomonas baetica: Pathogenicity for marine fish and development of protocols for rapid diagnosis. FEMS Microbiol. Lett. 2016, 364, fnw286. [Google Scholar] [CrossRef] [Green Version]
- Kozińska, A.; Paździor, E.; Pȩkala, A.; Niemczuk, W. Acinetobacter johnsonii and Acinetobacter lwoffii—The emerging fish pathogens. J. Vet. Res. 2014, 58, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35. [Google Scholar] [CrossRef] [Green Version]
- Nematollahi, A.; Decostere, A.; Pasmans, F.; Haesebrouck, F. Flavobacterium psychrophilum infections in salmonid fish. J. Fish Dis. 2003, 26, 563–574. [Google Scholar] [CrossRef]
- Jun, L.; Woo, N.Y.S. Pathogenicity of Vibrios in fish: An overview. J. Ocean Univ. 2003, 2, 117–128. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. Unifrac: An effective distance metric for microbial community comparison. Isme. J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, M.; Fujiwara-Nagata, E.; Katayama, T.; Suetake, H. Skin bacteria of rainbow trout antagonistic to the fish pathogen Flavobacterium psychrophilum. Sci. Rep. 2021, 11, 7518. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Takizawa, F.; Casadei, E.; Shibasaki, Y.; Ding, Y.; Sauters, T.J.C.; Yu, Y.; Salinas, I.; Sunyer, J.O. Specialization of mucosal immunoglobulins in pathogen control and microbiota homeostasis occurred early in vertebrate evolution. Sci. Immunol. 2020, 5, eaay3254. [Google Scholar] [CrossRef] [PubMed]
- Hasan, K.N.; Banerjee, G. Recent studies on probiotics as beneficial mediator in aquaculture: A review. J. Basic Appl. Zool. 2020, 81, 53. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorriehzahra, M.J.; Delshad, S.T.; Adel, M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: An update on their multiple modes of action: A review. Vet. Q. 2016, 36, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Dymock, D.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.T.; Giovannoni, S.J. Bias caused by template annealing in the amplification of mixtures of 16S rRNA genes by PCR. Appl. Environ. Microbiol. 1996, 62, 625–630. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identified Bacteria (16s rDNA V1–V3) | Percent Identity against Nucleotide Database (%) | 2019 (Month) | 2020 (Month) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (16S_Ribosomal_ RNA) | (nt) | 4 | 6 | 8 | 9 | 10 | 11 | 12 | 1 | 2 | 3 | Total | |
| Acinetobacter sp./movanagherensis | 95.62 | 96.12 (Acinetobacter sp) | 1 | 1 | |||||||||
| Chryseobacterium soldanellicola | 97.51 | 97.67 | 1 | 1 | |||||||||
| Lactococcus chungangensis | 99.74 | 99.74 | 1 | 1 | |||||||||
| L. raffinolactis | 99.48 | 100.00 | 2 | 2 | |||||||||
| Pseudomonas arsenicoxydans | 99.74 | 100.00 | 2 | 2 | |||||||||
| P. aylmerense/sp. | 99.48 | 100.00 (Pseudomonas. sp.) | 1 | 1 | |||||||||
| P. baetica | 99.23 | 100.00 | 1 | 1 | 4 | 6 | |||||||
| P. extremorientalis | 99.74 | 100.00 | 3 | 1 | 1 | 5 | |||||||
| P. fragi | 99.74 | 100.00 | 1 | 1 | |||||||||
| P. gessardii | 99.74 | 100.00 | 1 | 1 | |||||||||
| P. jessenii | 99.74 | 100.00 | 1 | 1 | |||||||||
| P. koreensis | 99.49 (P. baetica) | 100.00 | 3 | 1 | 3 | 4 | 1 | 12 | |||||
| P. marginalis/rhodesiae | 99.48 | 100.00 | 2 | 2 | |||||||||
| P. parafulva | 99.48 | 99.74 | 5 | 5 | |||||||||
| P. protegens | 99.74 | 100.00 | 1 | 1 | |||||||||
| Yersinia ruckeri/massiliensis | 99.49 | 100.00 | 3 | 1 | 4 | ||||||||
| unidentified | - | - | 2 | 2 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakatani, H.; Yamada, N.; Hashimoto, N.; Okazaki, F.; Arakawa, T.; Tamaru, Y.; Hori, K. Perturbation by Antimicrobial Bacteria of the Epidermal Bacterial Flora of Rainbow Trout in Flow-Through Aquaculture. Biology 2022, 11, 1249. https://doi.org/10.3390/biology11081249
Nakatani H, Yamada N, Hashimoto N, Okazaki F, Arakawa T, Tamaru Y, Hori K. Perturbation by Antimicrobial Bacteria of the Epidermal Bacterial Flora of Rainbow Trout in Flow-Through Aquaculture. Biology. 2022; 11(8):1249. https://doi.org/10.3390/biology11081249
Chicago/Turabian StyleNakatani, Hajime, Naoki Yamada, Naoki Hashimoto, Fumiyoshi Okazaki, Tomoko Arakawa, Yutaka Tamaru, and Katsutoshi Hori. 2022. "Perturbation by Antimicrobial Bacteria of the Epidermal Bacterial Flora of Rainbow Trout in Flow-Through Aquaculture" Biology 11, no. 8: 1249. https://doi.org/10.3390/biology11081249
APA StyleNakatani, H., Yamada, N., Hashimoto, N., Okazaki, F., Arakawa, T., Tamaru, Y., & Hori, K. (2022). Perturbation by Antimicrobial Bacteria of the Epidermal Bacterial Flora of Rainbow Trout in Flow-Through Aquaculture. Biology, 11(8), 1249. https://doi.org/10.3390/biology11081249