
Soluble Flt-1 in AMI Patients Serum Inhibits Angiogenesis of Endothelial Progenitor Cells by Suppressing Akt and Erk’s Activity

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and EPC Isolation
2.2. Flow Cytometry Analysis of EPCs
2.3. Immunofluorescence Identification of EPCs
2.4. Functional Identification of EPCs
2.5. Real-Time Quantitative Polymerase Chain Reaction
2.6. Western Blot
2.7. Enzyme-Linked Immunosorbent Assay
2.8. Statistical Analysis
3. Results
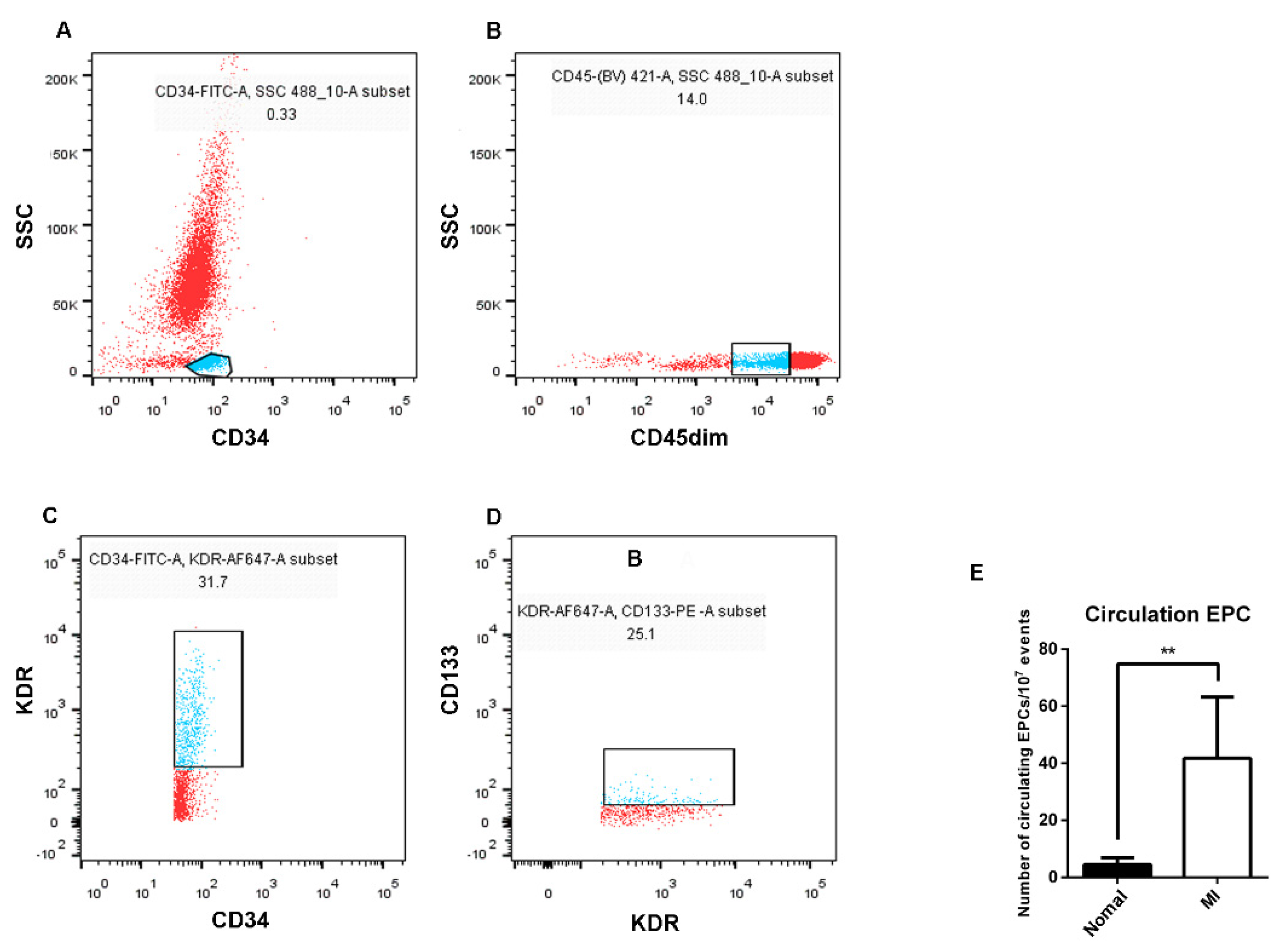
3.1. Isolation, Culture, and Identification of EPCs
3.2. The Number of EPCs in the Peripheral Blood of AMI Patients Was Higher than That of Healthy Subjects
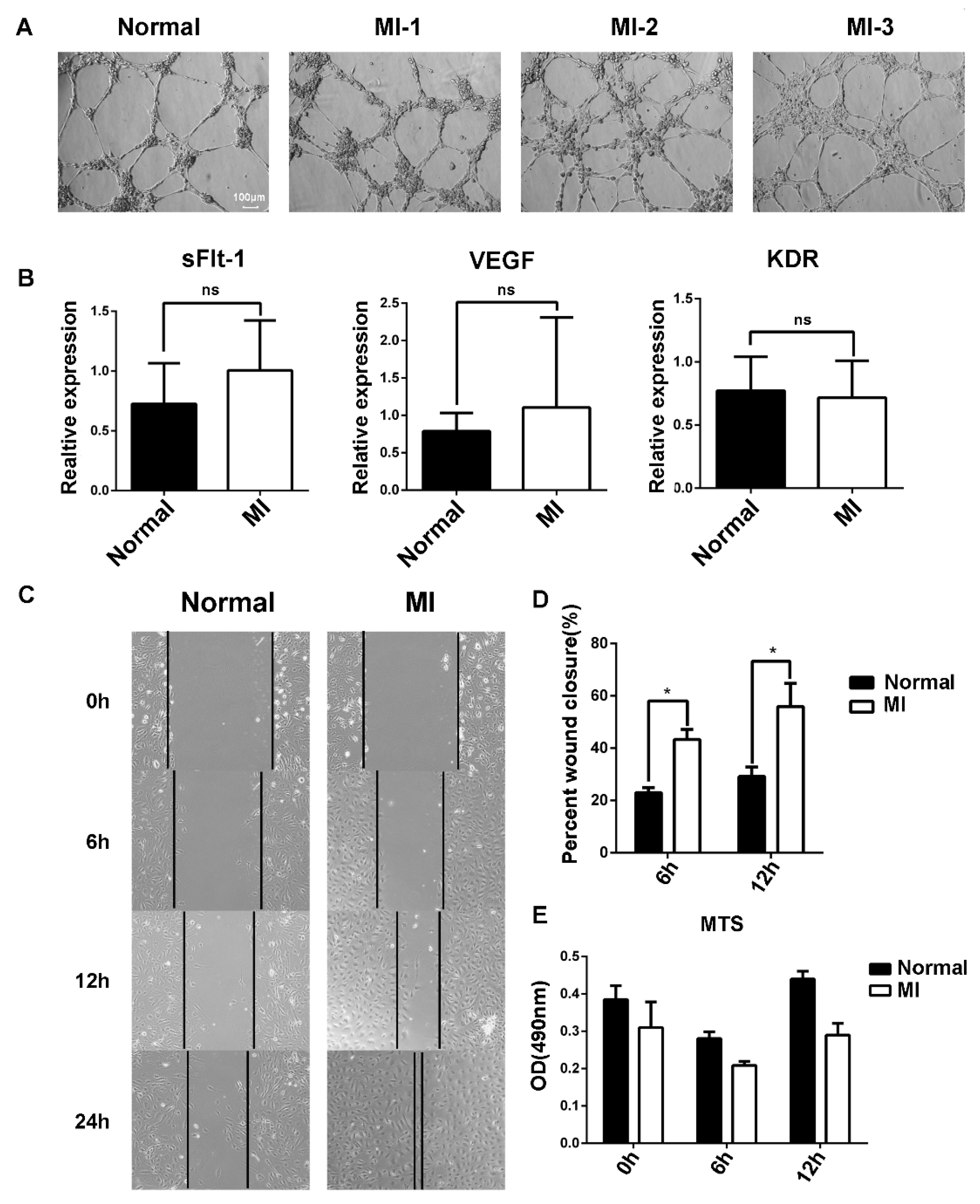
3.3. The Migration Ability of EPCs in the Peripheral Blood of AMI Patients Was Stronger That of than Healthy Subjects
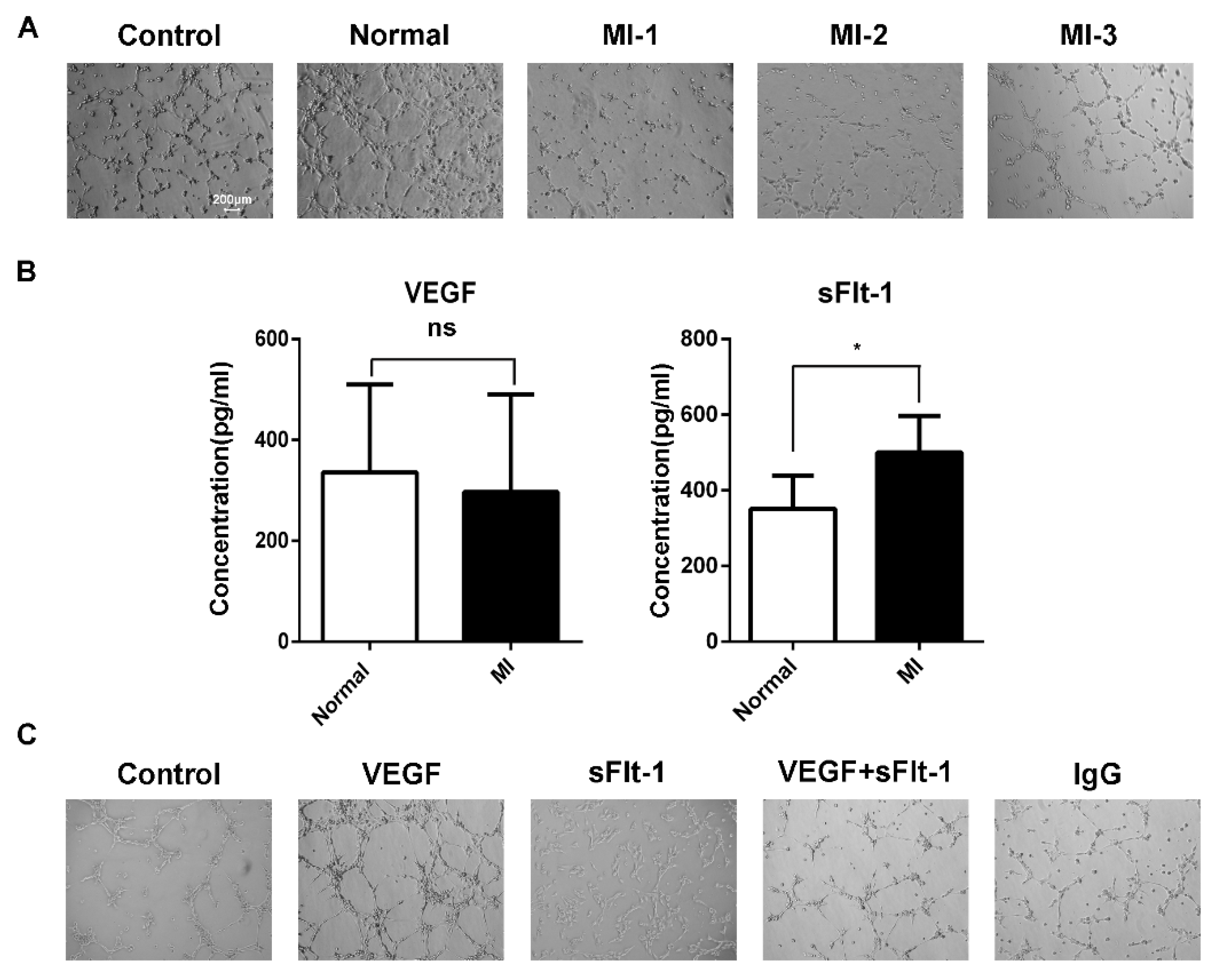
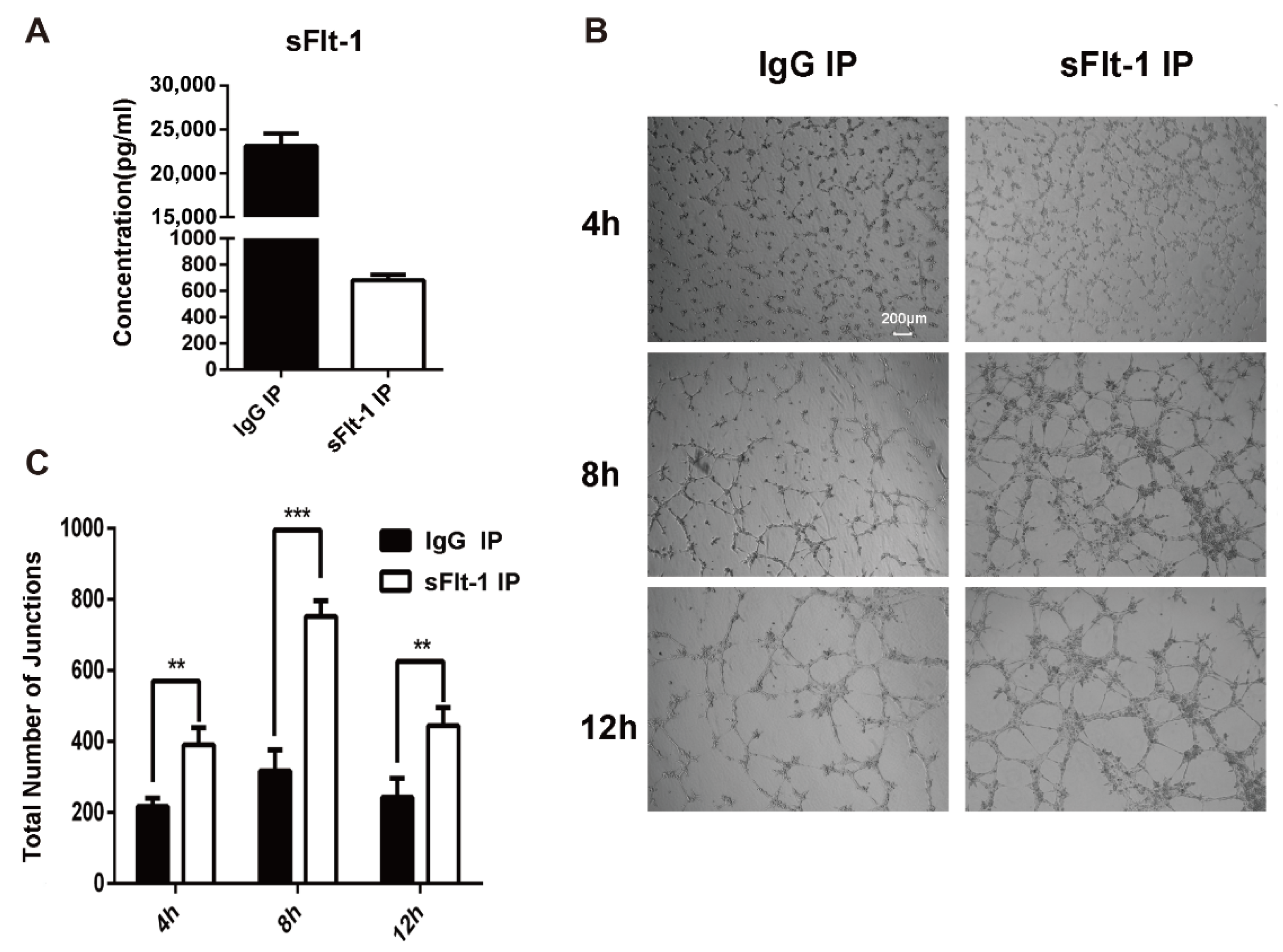
3.4. sFlt-1 in the Serum of AMI Patients Inhibited EPC Angiogenesis
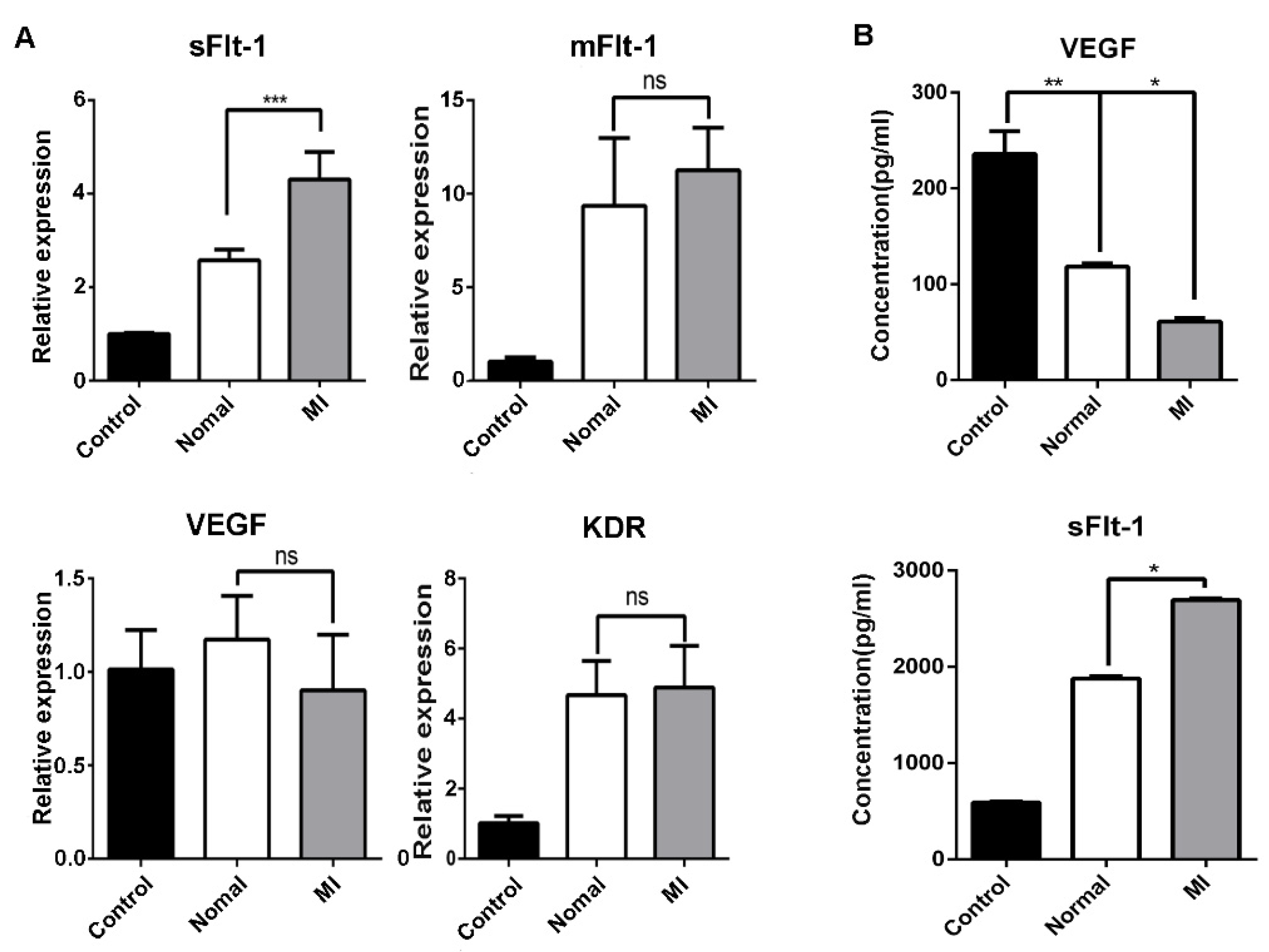
3.5. Serum from AMI Patients Promoted the Production of sFlt-1 by EPCs
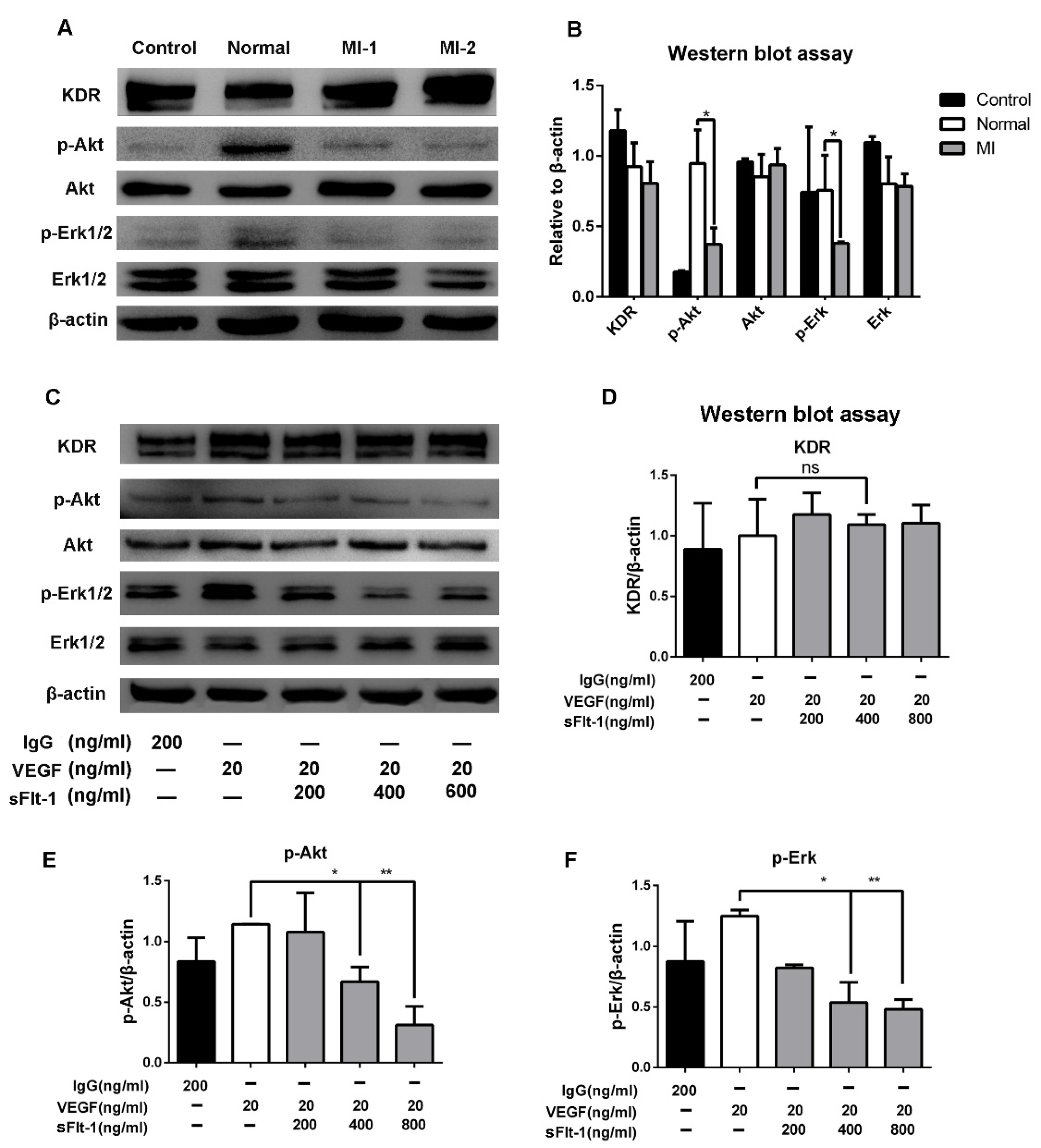
3.6. Akt and Erk 1/2 Signaling Were Hijacked by sFlt-1 to Inhibit Angiogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thiele, H.; Desch, S.; de Waha, S. Acute myocardial infarction in patients with ST-segment elevation myocardial infarction: ESC guidelines 2017. Herz 2017, 42, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Werner, N.; Kosiol, S.; Schiegl, T.; Ahlers, P.; Walenta, K.; Link, A.; Bohm, M.; Nickenig, G. Circulating endothelial progenitor cells and cardiovascular outcomes. N. Engl. J. Med. 2005, 353, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Figueras, J.; Otaegui, I.; Marti, G.; Domingo, E.; Baneras, J.; Barrabes, J.A.; Del Blanco, B.G.; Garcia-Dorado, D. Area at risk and collateral circulation in a first acute myocardial infarction with occluded culprit artery. STEMI vs non-STEMI patients. Int. J. Cardiol. 2018, 259, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Mitsos, S.; Katsanos, K.; Koletsis, E.; Kagadis, G.C.; Anastasiou, N.; Diamantopoulos, A.; Karnabatidis, D.; Dougenis, D. Therapeutic angiogenesis for myocardial ischemia revisited: Basic biological concepts and focus on latest clinical trials. Angiogenesis 2012, 15, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef]
- Lapidot, T.; Petit, I. Current understanding of stem cell mobilization: The roles of chemokines, proteolytic enzymes, adhesion molecules, cytokines, and stromal cells. Exp. Hematol. 2002, 30, 973–981. [Google Scholar] [CrossRef]
- Xia, X.; Toh, D.W.K.; Ng, S.L.; Zharkova, O.; Poh, K.K.; Foo, R.S.Y.; Wang, J.W.; Kim, J.E. Impact of following a healthy dietary pattern with co-consuming wolfberry on number and function of blood outgrowth endothelial cells from middle-aged and older adults. Food Funct. 2022, 13, 76–90. [Google Scholar] [CrossRef]
- Regueiro, A.; Cuadrado-Godia, E.; Bueno-Beti, C.; Diaz-Ricart, M.; Oliveras, A.; Novella, S.; Gene, G.G.; Jung, C.; Subirana, I.; Ortiz-Perez, J.T.; et al. Mobilization of endothelial progenitor cells in acute cardiovascular events in the PROCELL study: Time-course after acute myocardial infarction and stroke. J. Mol. Cell. Cardiol. 2015, 80, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Leal, V.; Ribeiro, C.F.; Oliveiros, B.; Antonio, N.; Silva, S. Intrinsic Vascular Repair by Endothelial Progenitor Cells in Acute Coronary Syndromes: An Update Overview. Stem. Cell. Rev. Rep. 2019, 15, 35–47. [Google Scholar] [CrossRef]
- Huang, H.; Huang, W. Regulation of Endothelial Progenitor Cell Functions in Ischemic Heart Disease: New Therapeutic Targets for Cardiac Remodeling and Repair. Front. Cardiovasc. Med. 2022, 9, 896782. [Google Scholar] [CrossRef]
- Matkar, P.N.; Ariyagunarajah, R.; Leong-Poi, H.; Singh, K.K. Friends Turned Foes: Angiogenic Growth Factors beyond Angiogenesis. Biomolecules 2017, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Sanyal, S.; Mukhopadhyay, D. Tyrosine residues 951 and 1059 of vascular endothelial growth factor receptor-2 (KDR) are essential for vascular permeability factor/vascular endothelial growth factor-induced endothelium migration and proliferation, respectively. J. Biol. Chem. 2001, 276, 32714–32719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Liebetrau, C.; Nef, H.M.; Dorr, O.; Gaede, L.; Hoffmann, J.; Hahnel, A.; Rolf, A.; Troidl, C.; Lackner, K.J.; Keller, T.; et al. Release kinetics of early ischaemic biomarkers in a clinical model of acute myocardial infarction. Heart 2014, 100, 652–657. [Google Scholar] [CrossRef]
- Tjwa, M.; Luttun, A.; Autiero, M.; Carmeliet, P. VEGF and PlGF: Two pleiotropic growth factors with distinct roles in development and homeostasis. Cell Tissue Res. 2003, 314, 5–14. [Google Scholar] [CrossRef]
- Takahashi, H.; Shibuya, M. The vascular endothelial growth factor (VEGF)/VEGF receptor system and its role under physiological and pathological conditions. Clin. Sci. 2005, 109, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling-in control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef]
- Estes, M.L.; Mund, J.A.; Mead, L.E.; Prater, D.N.; Cai, S.; Wang, H.; Pollok, K.E.; Murphy, M.P.; An, C.S.; Srour, E.F.; et al. Application of polychromatic flow cytometry to identify novel subsets of circulating cells with angiogenic potential. Cytom. A 2010, 77, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Fadini, G.P.; Losordo, D.; Dimmeler, S. Critical reevaluation of endothelial progenitor cell phenotypes for therapeutic and diagnostic use. Circ. Res. 2012, 110, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Groeneveld, D.J.; van Bekkum, T.; Dirven, R.J.; Wang, J.W.; Voorberg, J.; Reitsma, P.H.; Eikenboom, J. Angiogenic characteristics of blood outgrowth endothelial cells from patients with von Willebrand disease. J. Thromb. Haemost. 2015, 13, 1854–1866. [Google Scholar] [CrossRef]
- Wang, J.W.; Bouwens, E.A.; Pintao, M.C.; Voorberg, J.; Safdar, H.; Valentijn, K.M.; de Boer, H.C.; Mertens, K.; Reitsma, P.H.; Eikenboom, J. Analysis of the storage and secretion of von Willebrand factor in blood outgrowth endothelial cells derived from patients with von Willebrand disease. Blood 2013, 121, 2762–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornig, C.; Barleon, B.; Ahmad, S.; Vuorela, P.; Ahmed, A.; Weich, H.A. Release and complex formation of soluble VEGFR-1 from endothelial cells and biological fluids. Lab. Investig. 2000, 80, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zheng, Z.; Hu, H.; Zhou, Q.; Liu, W.; Li, X.; Liu, Z.; Wang, Y.; Ma, Y. A point-of-care selenium nanoparticle-based test for the combined detection of anti-SARS-CoV-2 IgM and IgG in human serum and blood. Lab Chip 2020, 20, 4255–4261. [Google Scholar] [CrossRef]
- Spadaccio, C.; Nenna, A.; Rose, D.; Piccirillo, F.; Nusca, A.; Grigioni, F.; Chello, M.; Vlahakes, G.J. The Role of Angiogenesis and Arteriogenesis in Myocardial Infarction and Coronary Revascularization. J. Cardiovasc. Transl. Res. 2022. [Google Scholar] [CrossRef]
- Shintani, S.; Murohara, T.; Ikeda, H.; Ueno, T.; Honma, T.; Katoh, A.; Sasaki, K.; Shimada, T.; Oike, Y.; Imaizumi, T. Mobilization of endothelial progenitor cells in patients with acute myocardial infarction. Circulation 2001, 103, 2776–2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massa, M.; Rosti, V.; Ferrario, M.; Campanelli, R.; Ramajoli, I.; Rosso, R.; De Ferrari, G.M.; Ferlini, M.; Goffredo, L.; Bertoletti, A.; et al. Increased circulating hematopoietic and endothelial progenitor cells in the early phase of acute myocardial infarction. Blood 2005, 105, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.M.; Zalos, G.; Halcox, J.P.; Schenke, W.H.; Waclawiw, M.A.; Quyyumi, A.A.; Finkel, T. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N. Engl. J. Med. 2003, 348, 593–600. [Google Scholar] [CrossRef]
- Pirro, M.; Schillaci, G.; Menecali, C.; Bagaglia, F.; Paltriccia, R.; Vaudo, G.; Mannarino, M.R.; Mannarino, E. Reduced number of circulating endothelial progenitors and HOXA9 expression in CD34+ cells of hypertensive patients. J. Hypertens. 2007, 25, 2093–2099. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, G.X.; Zhang, X.Y.; Xia, W.H.; Yang, Z.; Su, C.; Qiu, Y.X.; Xu, S.Y.; Zhan, H.; Tao, J. Lacidipine improves endothelial repair capacity of endothelial progenitor cells from patients with essential hypertension. Int. J. Cardiol. 2013, 168, 3317–3326. [Google Scholar] [CrossRef]
- Chong, S.Y.; Zharkova, O.; Yatim, S.; Wang, X.; Lim, X.C.; Huang, C.; Tan, C.Y.; Jiang, J.; Ye, L.; Tan, M.S.; et al. Tissue factor cytoplasmic domain exacerbates post-infarct left ventricular remodeling via orchestrating cardiac inflammation and angiogenesis. Theranostics 2021, 11, 9243–9261. [Google Scholar] [CrossRef]
- Huang, Z.; Bao, S.D. Roles of main pro- and anti-angiogenic factors in tumor angiogenesis. World J. Gastroenterol. 2004, 10, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Mac Gabhann, F.; Qutub, A.A.; Annex, B.H.; Popel, A.S. Systems biology of pro-angiogenic therapies targeting the VEGF system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 694–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buda, V.; Andor, M.; Petrescu, L.; Cristescu, C.; Baibata, D.E.; Voicu, M.; Munteanu, M.; Citu, I.; Muntean, C.; Cretu, O.; et al. Perindopril Induces TSP-1 Expression in Hypertensive Patients with Endothelial Dysfunction in Chronic Treatment. Int. J. Mol. Sci. 2017, 18, 348. [Google Scholar] [CrossRef] [PubMed]
- Hueso, L.; Rios-Navarro, C.; Ruiz-Sauri, A.; Chorro, F.J.; Nunez, J.; Sanz, M.J.; Bodi, V.; Piqueras, L. Dynamics and implications of circulating anti-angiogenic VEGF-A165b isoform in patients with ST-elevation myocardial infarction. Sci. Rep. 2017, 7, 9962. [Google Scholar] [CrossRef]
- Hochholzer, W.; Reichlin, T.; Stelzig, C.; Hochholzer, K.; Meissner, J.; Breidthardt, T.; Reiter, M.; Duehsler, B.; Freidank, H.; Winkler, K.; et al. Impact of soluble fms-like tyrosine kinase-1 and placental growth factor serum levels for risk stratification and early diagnosis in patients with suspected acute myocardial infarction. Eur. Heart J. 2011, 32, 326–335. [Google Scholar] [CrossRef]
- Gruson, D.; Hermans, M.P.; Ferracin, B.; Ahn, S.A.; Rousseau, M.F. Sflt-1 in heart failure: Relation with disease severity and biomarkers. Scand. J. Clin. Lab. Investig. 2016, 76, 411–416. [Google Scholar] [CrossRef]
- Kendall, R.L.; Thomas, K.A. Inhibition of vascular endothelial cell growth factor activity by an endogenously encoded soluble receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 10705–10709. [Google Scholar] [CrossRef] [Green Version]
- Kendall, R.L.; Wang, G.; Thomas, K.A. Identification of a natural soluble form of the vascular endothelial growth factor receptor, FLT-1, and its heterodimerization with KDR. Biochem. Biophys. Res. Commun. 1996, 226, 324–328. [Google Scholar] [CrossRef]
- Rajakumar, A.; Michael, H.M.; Rajakumar, P.A.; Shibata, E.; Hubel, C.A.; Karumanchi, S.A.; Thadhani, R.; Wolf, M.; Harger, G.; Markovic, N. Extra-placental expression of vascular endothelial growth factor receptor-1, (Flt-1) and soluble Flt-1 (sFlt-1), by peripheral blood mononuclear cells (PBMCs) in normotensive and preeclamptic pregnant women. Placenta 2005, 26, 563–573. [Google Scholar] [CrossRef]
- Ye, L.; Gratton, A.; Hannan, N.J.; Cannon, P.; Deo, M.; Palmer, K.R.; Tong, S.; Kaitu’u-Lino, T.J.; Brownfoot, F.C. Nuclear factor of activated T-cells (NFAT) regulates soluble fms-like tyrosine kinase-1 secretion (sFlt-1) from human placenta. Placenta 2016, 48, 110–118. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence | |
|---|---|---|
| Forward | Reverse | |
| β-actin | CCACCATGTACCCTGGCATT | CGGACTCGTCATACTCCTGC |
| VEGF | GGGCCTCCGAAACCATGAACTT | AGGGGCACACAGGATGGCTTGA |
| sFlt-1 | GCGCATGGCAATAATAGAAGGAAA | CCTTTTTGTTGCAGTGCTCACCTC |
| mFlt-1 | GCCCGGGAGAGACTTAAACTGG | GGCCTGGCTCCATTTTTTCTTTC |
| KDR | CAACAAAGTCGGGAGAGGAG | ATGACGATGGACAAGTAGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, X.; Zhong, X.; Fan, M.; Wang, G.; Shi, W.; Xie, R.; Wei, Y.; Zhang, H.; Meng, X.; et al. Soluble Flt-1 in AMI Patients Serum Inhibits Angiogenesis of Endothelial Progenitor Cells by Suppressing Akt and Erk’s Activity. Biology 2022, 11, 1194. https://doi.org/10.3390/biology11081194
Zhang L, Zhang X, Zhong X, Fan M, Wang G, Shi W, Xie R, Wei Y, Zhang H, Meng X, et al. Soluble Flt-1 in AMI Patients Serum Inhibits Angiogenesis of Endothelial Progenitor Cells by Suppressing Akt and Erk’s Activity. Biology. 2022; 11(8):1194. https://doi.org/10.3390/biology11081194
Chicago/Turabian StyleZhang, Lijie, Xingkun Zhang, Xiaoming Zhong, Mengya Fan, Guoliang Wang, Wei Shi, Ran Xie, Yinxiang Wei, Hailong Zhang, Xiangxu Meng, and et al. 2022. "Soluble Flt-1 in AMI Patients Serum Inhibits Angiogenesis of Endothelial Progenitor Cells by Suppressing Akt and Erk’s Activity" Biology 11, no. 8: 1194. https://doi.org/10.3390/biology11081194
APA StyleZhang, L., Zhang, X., Zhong, X., Fan, M., Wang, G., Shi, W., Xie, R., Wei, Y., Zhang, H., Meng, X., Wang, Y., & Ma, Y. (2022). Soluble Flt-1 in AMI Patients Serum Inhibits Angiogenesis of Endothelial Progenitor Cells by Suppressing Akt and Erk’s Activity. Biology, 11(8), 1194. https://doi.org/10.3390/biology11081194