
Characterization of Extracellular Vesicles Secreted by a Clinical Isolate of Naegleria fowleri and Identification of Immunogenic Components within Their Protein Cargo

 ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Axenic Culture of Naegleria fowleri Trophozoites
2.2. Isolation of Extracellular Vesicles (EVs)
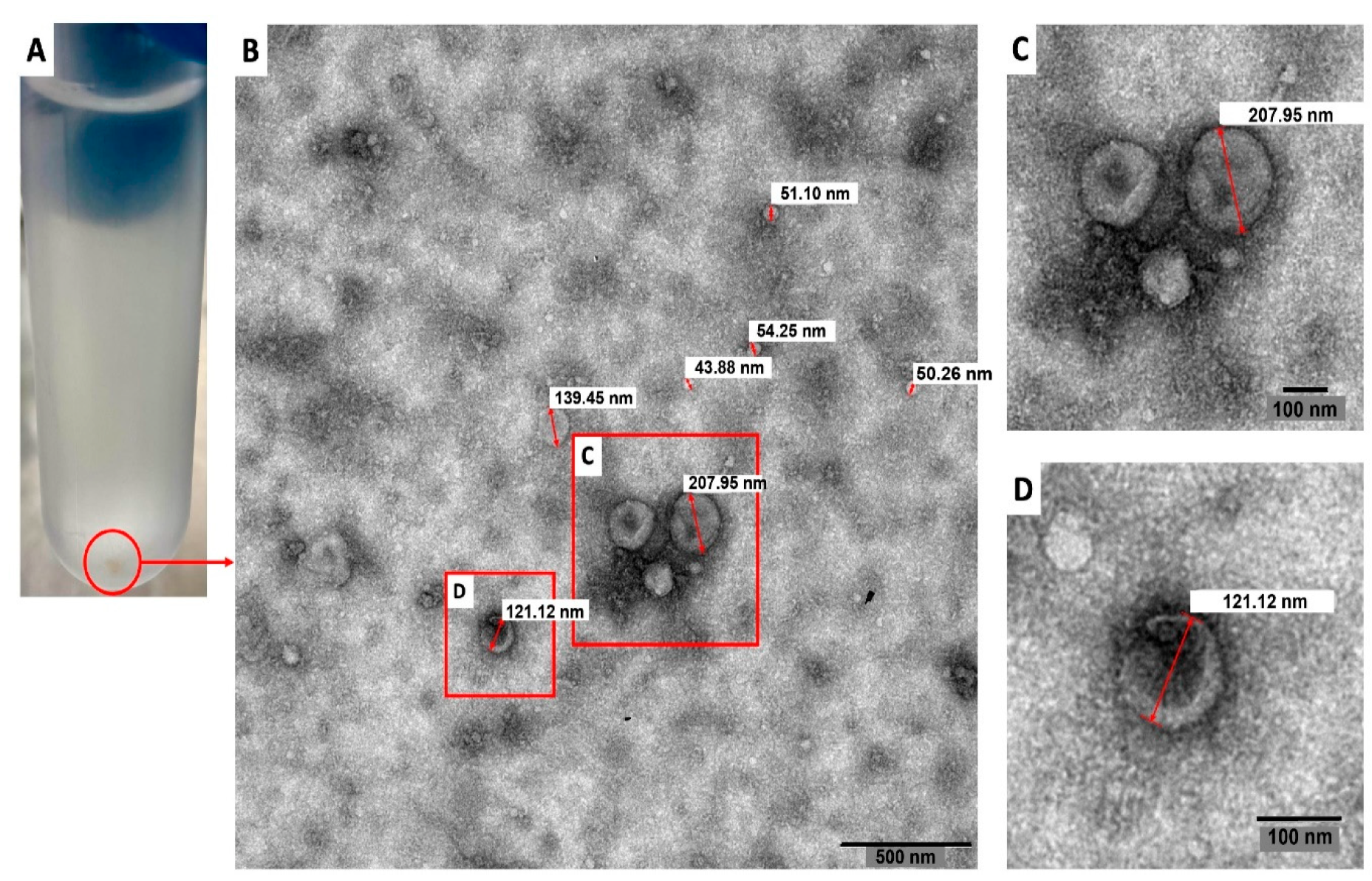
2.3. Transmission Electron Microscopy
2.4. Atomic Force Microscopy
2.5. Nanoparticle Tracking Analysis
2.6. Dynamic Light Scattering
2.7. Measurement of Zeta Potential
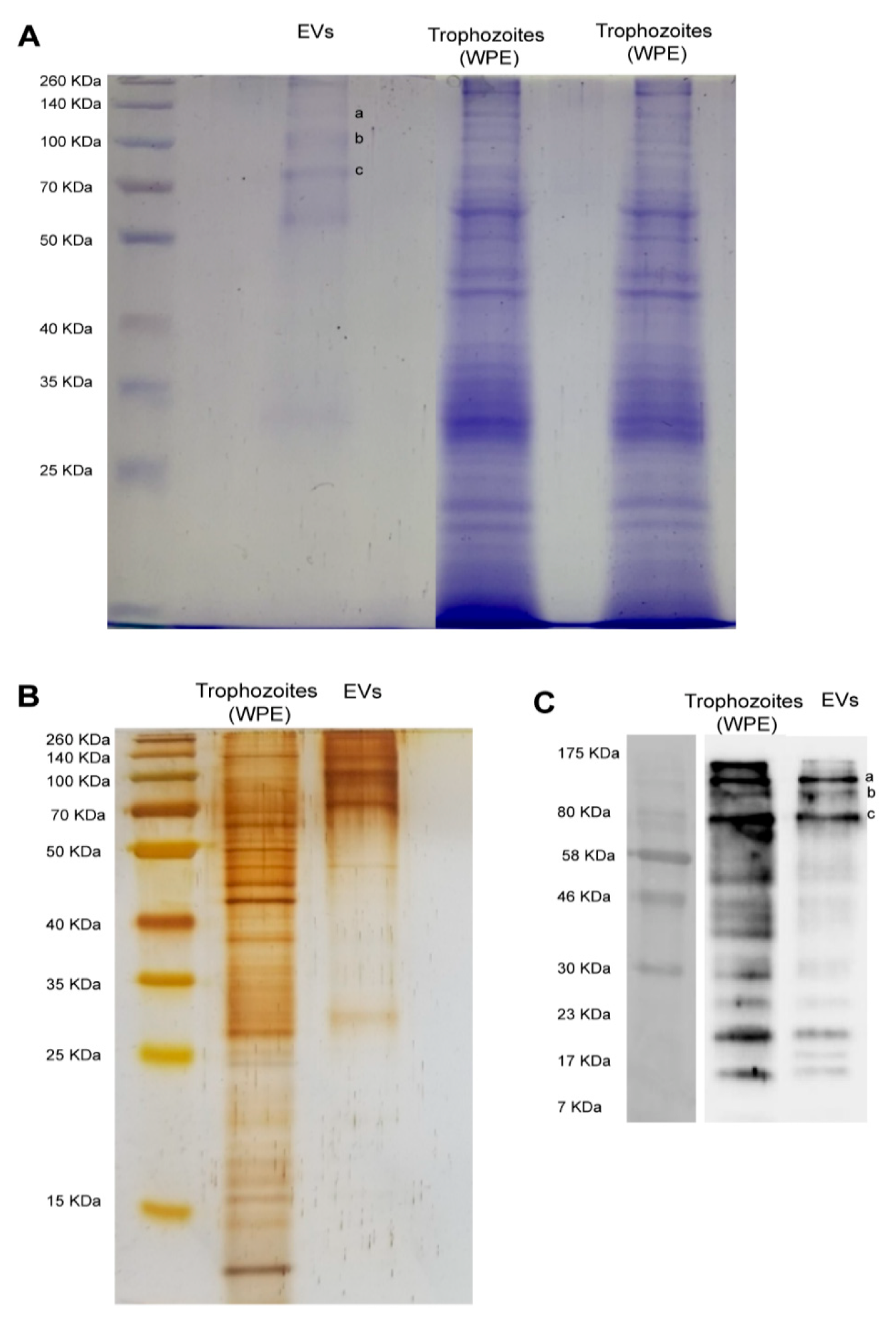
2.8. Protein Pattern and Recognition of Extracellular Vesicles by Rat Polyclonal Anti-Naegleria fowleri Antibodies
2.8.1. Preparation of Whole Protein Extracts of Naegleria fowleri Trophozoites
2.8.2. Electrophoretic Separation of Proteins Using SDS-PAGE
2.8.3. Polyclonal Antibody Production
2.8.4. Western Blot
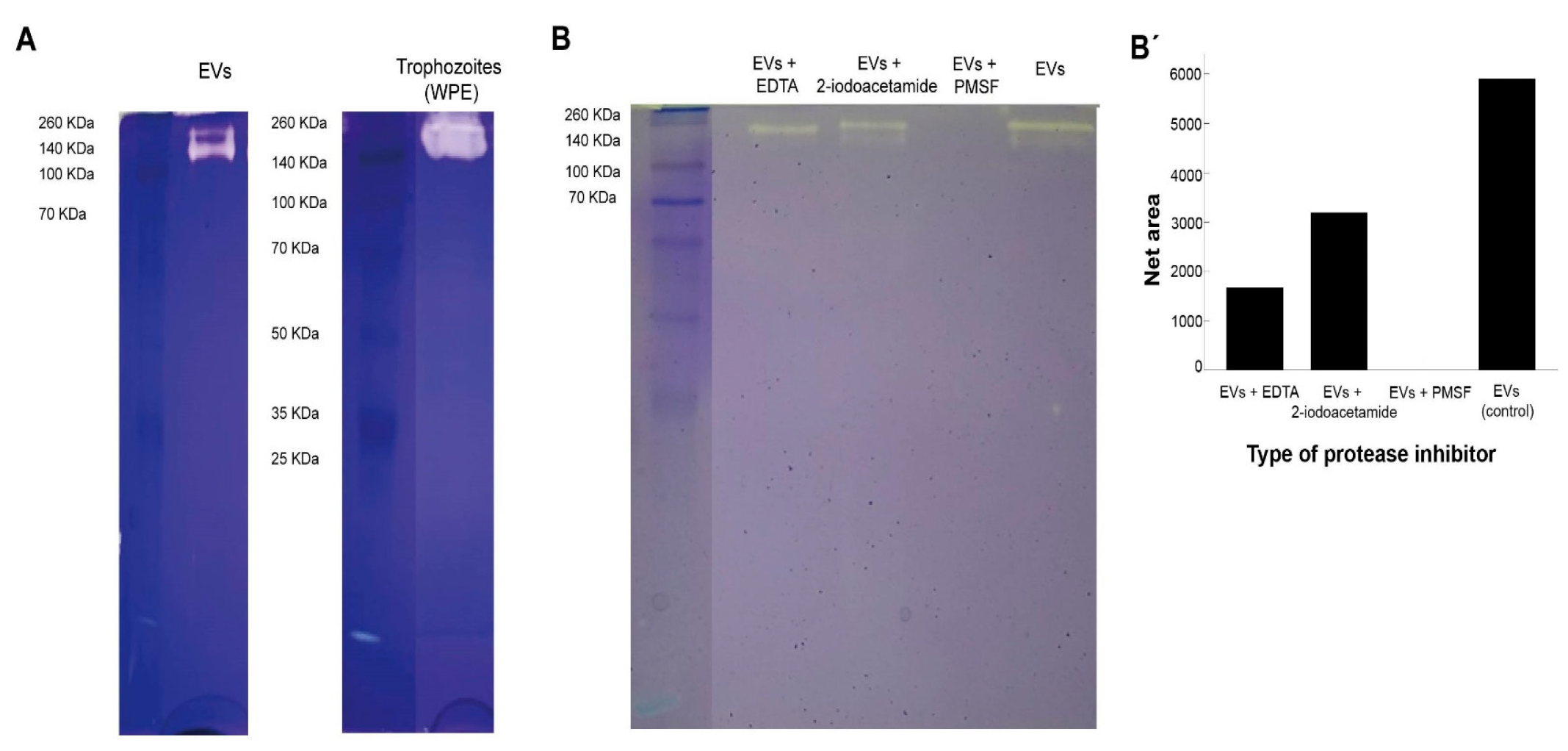
2.9. Evaluation of Protease Activity of Extracellular Vesicles
2.10. Proteomic Analyses
2.10.1. Protein Identification from SDS-PAGE/Western Blot Immunogenic Bands
2.10.2. Proteome Profiling of Whole EVs by Bottom-Up Shotgun Analysis
3. Results
3.1. Characterization of Extracellular Vesicles
3.1.1. Transmission Electron Microscopy and Atomic Force Microscopy
3.1.2. Nanoparticle Tracking Analysis and Dynamic Light Scattering
3.2. Protein Profile and Recognition of Extracellular Vesicles by Anti-Naegleria fowleri Polyclonal Antibodies
3.3. Protease Activity of Extracellular Vesicles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van der Pol, E.; Boing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, M.L.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Hargett, L.A.; Bauer, N.N. On the origin of microparticles: From “platelet dust” to mediators of intercellular communication. Pulm. Circ. 2013, 3, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular vesicle heterogeneity: Subpopulations, isolation techniques, and diverse functions in cancer progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Ciferri, M.C.; Quarto, R.; Tasso, R. Extracellular vesicles as biomarkers and therapeutic tools: From pre-clinical to clinical applications. Biology 2021, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Kodam, S.P.; Ullah, M. Diagnostic and therapeutic potential of extracellular vesicles. Technol. Cancer Res. Treat. 2021, 20, 15330338211041203. [Google Scholar] [CrossRef]
- Barnie, P.A.; Afrifa, J.; Gyamerah, E.O.; Amoani, B. Extracellular vesicles as biomarkers and therapeutic targets in cancers. In Extracellular Vesicles—Role in Diseases Pathogenesis and Therapy; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Khosravi, M.; Mirsamadi, E.S.; Mirjalali, H.; Zali, M.R. Isolation and functions of extracellular vesicles derived from parasites: The promise of a new era in immunotherapy, vaccination, and diagnosis. Int. J. Nanomed. 2020, 15, 2957–2969. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; de’Oliveira, C.C.; Shirvani, O.; Fang, Y.; Wang, C.; Foster, L.J.; Reiner, N.E. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Cell Sci. 2010, 123, 842–852. [Google Scholar] [CrossRef] [Green Version]
- Torrecilhas, A.C.; Schumacher, R.I.; Alves, M.J.; Colli, W. Vesicles as carriers of virulence factors in parasitic protozoan diseases. Microbes Infect. 2012, 15, 1465–1474. [Google Scholar] [CrossRef]
- Cwiklinski, C.; de la Torre-Escudero, E.; Trelis, M.; Bernal, D.; Dufresne, P.J.; Brennan, G.P.; O’Neill, S.; Tort, J.F.; Paterson, S.; Marcilla, A.; et al. The extracellular vesicles of the helminth pathogen, Fasciola hepatica: Biogenesis pathways and cargo molecules involved in parasite pathogenesis. Mol. Cell. Proteom. 2015, 14, 3258–3273. [Google Scholar] [CrossRef] [Green Version]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gasan, T.A.; Kuipers, M.E.; Roberts, G.H.; Padalino, G.; Forde-Thomas, J.E.; Wilson, S.; Wawrzyniak, J.; Tukahebwa, E.M.; Hoffmann, K.F.; Chalmers, I.W. Schistosoma mansoni larval extracellular vesicle protein 1 (SmLEV1) is an immunogenic antigen found in EVs released from pre-acetabular glands of invading cercariae. PLoS Negl. Trop. Dis. 2021, 15, e0009981. [Google Scholar] [CrossRef] [PubMed]
- Marcilla, A.; Trelis, M.; Cortés, A.; Sotillo, J.; Cantalapiedra, F.; Minguez, M.T.; Valero, M.L.; del Pino, M.S.; Muñoz-Antoli, C.; Toledo, R.; et al. Extracellular vesicles from parasitic helminths contain specific excretory/secretory proteins and are internalized in intestinal host cells. PLoS ONE 2012, 7, e45974. [Google Scholar] [CrossRef]
- Aline, F.; Bout, D.; Amigorena, S.; Roingeard, P.; Dimier Poisson, I. Toxoplasma gondii antigen-pulsed dendritic cell-derived exosomes induce a protective immune response against T. gondii infection. Infect. Immun. 2004, 72, 4127–4137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrecilhas, A.C.; Tonelli, R.R.; Pavanelli, W.R.; da Silva, J.S.; Schumacher, R.I.; de Souza, W.; de Silva, N.C.; Abrahamsohn, I.A.; Colli, W.; Manso Alves, M.J. Trypanosoma cruzi: Parasite shed vesicles increase heart parasitism and generate an intense inflammatory response. Microbes. Infect. 2009, 11, 29–39. [Google Scholar] [CrossRef]
- Cestari, I.; Ansa Addo, E.; Deolindo, P.; Inal, J.M.; Ramirez, M.I. Trypanosoma cruzi immune evasion mediated by host cell derived microvesicles. J. Immunol. 2012, 188, 1942–1952. [Google Scholar] [CrossRef] [Green Version]
- Díaz Lozano, I.M.; de Pablos, L.M.; Longhi, S.A.; Zago, M.P.; Schijman, A.G.; Osuna, A. Immune complexes in chronic Chagas disease patients are formed by exovesicles from Trypanosoma cruzi carrying the conserved MASPs N-terminal region. Sci. Rep. 2017, 7, 44451. [Google Scholar] [CrossRef] [Green Version]
- Retana Moreira, L.; Rodríguez Serrano, F.; Osuna, A. Extracellular vesicles of Trypanosoma cruzi tissue-culture cell-derived trypomastigotes: Induction of physiological changes in non-parasitized culture cells. PLoS Negl. Trop. Dis. 2019, 13, e0007163. [Google Scholar] [CrossRef] [Green Version]
- Retana Moreira, L.; Prescilla-Ledezma, A.; Cornet-Gomez, A.; Linares, F.; Jódar-Reyes, A.B.; Fernandez, J.; Ibarrola Vannucci, A.K.; De Pablos, L.M.; Osuna, A. Biophysical and biochemical comparison of extracellular vesicles produced by infective and non-infective stages of Trypanosoma cruzi. Int. J. Mol. Sci. 2021, 22, 5183. [Google Scholar] [CrossRef]
- Zauli, R.C.; Vidal, A.S.; Dupin, T.V.; de Morais, A.C.C.; Batista, W.L.; Xander, P. Extracellular vesicles released by Leishmania: Impact on disease development and immune system cells. In Leishmaniasis—General Aspects of a Stigmatized Disease; de Azevedo Calderonon, L., Ed.; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Martin Jaular, L.; Nakayasu, E.S.; Ferrer, M.; Almeida, I.C.; del Portillo, H.A. Exosomes from Plasmodium yoelii-infected reticulocytes protect mice from lethal infections. PLoS ONE 2011, 6, e26588. [Google Scholar] [CrossRef] [Green Version]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and other extracellular vesicles in host-pathogen interactions. EMBO Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nievas, Y.R.; Lizarraga, A.; Salas, N.; Cóceres, V.M.; de Miguel, N. Extracellular vesicles released by anaerobic protozoan parasites: Current situation. Cell Microbiol. 2020, 22, e13257. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, D.; da Silva Ferreira, M.; Guimarães, A.J. Extracellular vesicles from the protozoa Acanthamoeba castellanii: Their role in pathogenesis, environmental adaptation and potential applications. Bioengineering 2019, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.C.; Tsai, C.Y.; Huang, J.M.; Wu, S.R.; Chu, L.J.; Huang, K.Y. Quantitative proteomic analysis and functional characterization of Acanthamoeba castellanii exosome-like vesicles. Parasit. Vectors 2019, 12, 467. [Google Scholar] [CrossRef] [PubMed]
- Retana Moreira, L.; Vargas Ramírez, D.; Linares, F.; Prescilla Ledezma, A.; Vaglio Garro, A.; Osuna, A.; Lorenzo Morales, J.; Abrahams Sandí, E. Isolation of Acanthamoeba T5 from water: Characterization of its pathogenic potential, including the production of extracellular vesicles. Pathogens 2020, 9, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.O.; Chagas, I.A.R.; de Menezes-Neto, A.; Rêgo, F.D.; Nogueira, P.M.; Torrecilhas, A.C.; Furst, C.; Fux, B.; Soares, R.P. Distinct immunomodulatory properties of extracellular vesicles released by different strains of Acanthamoeba. Cell Biol. Int. 2021, 45, 1060–1071. [Google Scholar] [CrossRef]
- Jahangeer, M.; Mahmood, Z.; Munir, N.; Waraich, U.; Mahmood Tahir, I.; Muhammad, A.; Muhammad Ali Shah, S.; Zulfqar, A.; Zainab, R. Naegleria fowleri: Sources of infection, pathophysiology, diagnosis and management; a review. Clin. Exp. Pharmacol. Physiol. 2019, 47, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Matanock, A.; Mehal, J.M.; Liu, L.; Blau, D.M.; Cope, J.R. Estimation of undiagnosed Naegleria fowleri primary amebic meningoencephalitis, United States. Emerg. Inf. Dis. 2018, 24, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Marciano-Cabral, F.; Cabral, G.A. The immune response to Naegleria fowleri amebae and pathogenesis of infection. FEMS Immunol. Med. Microbiol. 2007, 51, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.A.; Yazdani, N.; Ahmad, R.; Zehra, F.; Ahmad, N. Epidemiology of primary amoebic meningoencephalitis-related deaths due to Naegleria fowleri infections from freshwater in Pakistan: An analysis of 8-year dataset. Arch. Pharm. Pract. 2016, 7, 11–19. [Google Scholar] [CrossRef]
- Grace, E.; Asbill, S.; Virga, K. Naegleria fowleri: Pathogenesis, diagnosis, and treatment options. Antimicrob. Agents Chemother. 2015, 59, 6677–6681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retana Moreira, L.; Zamora Rojas, L.; Grijalba Murillo, M.; Molina Castro, S.E.; Abrahams Sandí, E. Primary amebic meningoencephalitis related to groundwater in Costa Rica: Diagnostic confirmation of three cases and environmental investigation. Pathogens 2020, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.; Aikawa, E.; Alacaraz, M.J.; Anderson, J.D.; Andriantsitohaina, A.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV): A position statement of the International Society for Extracellular Vesicles and update of the MISEV guidelines. J. Extracel. Ves. 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Heukeshoven, J.; Dernick, R. Improved silver staining procedure for fast staining in PhastSystem Development Unit. I. Staining of sodium dodecyl sulfate gels. Electrophoresis 1988, 9, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [Green Version]
- Herron, G.S.; Banda, M.J.; Clark, E.J.; Gavrilovic, J.; Werb, Z. Secretion of metalloproteinases by stimulated capillary endothelial cells. II. Expression of collagenase and stromelysin activities is regulated by endogenous inhibitors. J. Biol. Chem. 1986, 261, 2814–2818. [Google Scholar] [CrossRef]
- Castro Artavia, E.; Retana Moreira, L.; Lorenzo-Morales, J.; Abrahams Sandí, E. Potentially pathogenic Acanthamoeba genotype T4 isolated from dental units and emergency combination showers. Mem. Inst. Oswaldo Cruz 2017, 112, 817–821. [Google Scholar] [CrossRef] [Green Version]
- Lomonte, B.; Fernández, J. Solving the microheterogeneity of Bothrops asper myotoxin-II by high-resolution mass spectrometry: Insights into C-terminal region variability in Lys49-phospholipase A2 homologs. Toxicon 2022, 210, 123–131. [Google Scholar] [CrossRef]
- De Pablos Torró, L.M.; Retana Moreira, L.; Osuna, A. Extracellular vesicles in Chagas disease: A new passenger for an old disease. Front. Microbiol. 2018, 9, 1190. [Google Scholar] [CrossRef] [Green Version]
- Nederveen, J.P.; Warnier, G.; Di Carlo, A.; Nilsson, M.I.; Tarnopolsky, M.A. Extracellular vesicles and exosomes: Insights from exercise science. Front. Physiol. 2021, 11, 604274. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.; Cwiklinski, K.; Lalor, R.; O’Connell, B.; Robinson, M.W.; Gerlach, J.; Joshi, L.; Kilcoyne, M.; Dalton, J.P.; O’Neill, S.M. Fasciola hepatica extracellular vesicles isolated from excretory-secretory products using a gravity flow method modulate dendritic cell phenotype and activity. PLoS Negl. Trop. Dis. 2020, 14, e0008626. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, L.; Li, J.; Wang, L.; Wu, Z.; Sun, X. Extracellular vesicle-mediated communication within host-parasite interactions. Front. Immunol. 2019, 15, 3066. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, D.S.; Ferreira, M.D.S.; Liedke, S.C.; Gomes, K.X.; de Oliveira, G.A.; Leão, P.E.L.; Cesar, G.V.; Seabra, S.H.; Cortines, J.R.; Casadevall, A.; et al. Extracellular vesicles and vesicle-free secretome of the protozoa Acanthamoeba castellanii under homeostasis and nutritional stress and their damaging potential to host cells. Virulence 2018, 9, 818–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudbergsson, J.M.; Johnsen, K.B.; Skov, M.N.; Duroux, M. Systematic review of factors influencing extracellular vesicle yield from cell cultures. Cytotechnology 2016, 68, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Beit Yannai, E.; Tabak, S.; Stamer, W.D. Physical exosome:exosome interactions. J. Cell. Mol. Med. 2018, 22, 2001–2006. [Google Scholar] [CrossRef] [Green Version]
- Midekessa, G.; Godakumara, K.; Ord, J.; Viil, J.; Lättekivi, F.; Dissanayake, K.; Kopanchuk, S.; Rinken, A.; Andronowska, A.; Bhattacharjee, S.; et al. Zeta potential of extracellular vesicles: Toward understanding the attributes that determine colloidal stability. ACS Omega 2020, 5, 16701–16710. [Google Scholar] [CrossRef]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [Green Version]
- Soares Martins, T.; Catita, J.; Martins Rosa, I.; da Cruz, A.B.E.; Silva, O.; Henriques, A.G. Exosome isolation from distinct biofluids using precipitation and column-based approaches. PLoS ONE 2018, 13, e0198820. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, N.G.; Cheng, L.; Eriksson, E.M. The role of extracellular vesicles in malaria biology and pathogenesis. Malar J. 2017, 16, 245. [Google Scholar] [CrossRef] [Green Version]
- Couper, K.N.; Barnes, T.; Hafalla, J.C.; Combes, V.; Ryffel, B.; Secher, T.; Grau, G.; Riley, E.M.; De Souza, J.B. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010, 6, e1000744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigione, I.; Facchetti, P.; Lecordier, L.; Deslée, D.; Chiesa, S.; Cesbron-Delauw, M.F.; Pistoia, V. T cell clones raised from chronically infected healthy humans by stimulation with Toxoplasma gondii excretory-secretory antigens cross-react with live tachyzoites: Characterization of the fine antigenic specificity of the clones and implications for vaccine development. J. Immunol. 2000, 164, 3741–3748. [Google Scholar] [CrossRef] [Green Version]
- Costa-Silva, T.A.; Borges, M.M.; Galhardo, C.S.; Pereira-Chioccola, V.L. Immunization with excreted/secreted proteins in AS/n mice activating cellular and humoral response against Toxoplasma gondii infection. Acta Trop. 2012, 124, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.O.; Maia, M.M.; Torrecilhas, A.C.; Taniwaki, N.N.; Namiyama, G.M.; Oliveira, K.C.; Ribeiro, K.S.; Toledo, M.D.S.; Xander, P.; Pereira-Chioccola, V.L. Extracellular vesicles isolated from Toxoplasma gondii induce host immune response. Parasite Immunol. 2018, 40, e12571. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.; et al. Leishmania exosomes modulate innate and adaptive immune responses through effects on monocytes and dendritic cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef] [Green Version]
- Hassani, K.; Antoniak, E.; Jardim, A.; Olivier, M. Temperature-induced protein secretion by Leishmania mexicana modulates macrophage signalling and function. PLoS ONE 2011, 6, e18724. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.; Kelly, P.H.; Singh, B.K.; Pope, R.M.; Kim, P.; Zhanbolat, B.; Wilson, M.E.; Yao, C. Extracellular release of virulence factor major surface protease via exosomes in Leishmania infantum promastigotes. Parasit. Vectors 2018, 11, 355. [Google Scholar] [CrossRef]
- Gutiérrez-Sánchez, M.; Carrasco-Yepez, M.M.; Herrera-Díaz, J.; Rojas-Hernández, S. Identification of differential protein recognition pattern between Naegleria fowleri and Naegleria lovaniensis. Parasite Immunol. 2020, 42, e12715. [Google Scholar] [CrossRef]
- Tillery, L.; Barrett, K.; Goldstein, J.; Lassner, J.W.; Osterhout, B.; Tran, N.L.; Xu, L.; Young, R.M.; Craig, J.; Chun, I.; et al. Naegleria fowleri: Protein structures to facilitate drug discovery for the deadly, pathogenic free-living amoeba. PLoS ONE 2021, 16, e0241738. [Google Scholar] [CrossRef]
- Martínez-Castillo, M.; Ramírez-Rico, G.; Serrano-Luna, J.; Shibayama, M. Iron-binding protein degradation by cysteine proteases of Naegleria fowleri. BioMed Res. Int. 2015, 2015, 416712. [Google Scholar] [CrossRef] [Green Version]
- Vyas, I.K.; Jamerson, M.; Cabral, G.A.; Marciano-Cabral, F. Identification of peptidases in highly pathogenic vs. weakly pathogenic Naegleria fowleri amebae. J. Eukaryot. Microbiol. 2015, 62, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.M.; Reiner, N.E. Leishmania exosomes deliver preemptive strikes to create an environment permissive for early infection. Front. Cell. Infect. Microbiol. 2012, 1, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timm, T.; Annoscia, G.; Klein, J.; Lochnit, G. The eukaryotic elongation factor 1 alpha (eEF1α) from the parasite Leishmania infantum is modified with the immunomodulatory substituent phosphorylcholine (PC). Molecules 2017, 22, 2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Luna, J.; Cervantes-Sandoval, I.; Tsutsumi, V.; Shibayama, M. A biochemical comparison of proteases from pathogenic Naegleria fowleri and non-pathogenic Naegleria gruberi. J. Eukaryot. Microbiol. 2007, 54, 411–417. [Google Scholar] [CrossRef]
- Khan, N.A.; Jarroll, E.L.; Panjwani, N.; Cao, Z.; Paget, T.A. Proteases as markers for differentiation of pathogenic and nonpatogenic species of Acanthamoeba. J. Clin. Microbiol. 2000, 38, 2858–2861. [Google Scholar] [CrossRef] [Green Version]
- Alsam, S.; Sissons, J.; Jayasekera, S.; Khan, N.A. Extracellular proteases of Acanthamoeba castellanii (encephalitis isolate belonging to T1 genotype) contribute to increased permeability in an in vitro model of the human blood-brain barrier. J. Infect. 2005, 51, 150–156. [Google Scholar] [CrossRef]
- Omaña-Molina, M.; González Robles, A.; Salazar Villatoro, L.I.; Lorenzo Morales, J.; Cristóbal Ramos, A.R.; Hernández Ramírez, V.I. Reevaluating the role of Acanthamoeba proteases in tissue invasion: Observation of cytopathogenic mechanisms on MDCK cell. monolayers and hamster corneal cells. BioMed Res. Int. 2013, 2013, 461329. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; De, T.; Chakraborti, T. Leishmania donovani secretory serine protease alters macrophage inflammatory response via COX-2 mediated PGE-2 production. Indian J. Biochem. Biophys. 2014, 51, 542–551. [Google Scholar]
- Cano, A.; Mattana, A.; Henriquez, F.L.; Alexander, J.; Roberts, C.W. Acanthamoeba proteases contribute to macrophage activation through PAR1, but not PAR2. Parasite Immunol. 2019, 41, e12612. [Google Scholar] [CrossRef] [Green Version]
- Koussis, K.; Withers-Martinez, C.; Yeoh, S.; Child, M.; Hackett, F.; Knuepfer, E.; Juliano, L.; Woehlbier, U.; Bujard, H.; Blackman, M.J. A multifunctional serine protease primes the malaria parasite for red blood cell invasion. EMBO J. 2009, 28, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.R.; Hackett, F.; Strath, M.; Penzo, M.; Withers-Martinez, C.; Baker, D.; Blackman, M.J. Malaria parasite cGMP-dependent protein kinase regulates blood stage merozoite secretory organelle discharge and egress. PLoS Pathog. 2013, 9, e1003344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, C.R.; Hackett, F.; Atid, J.; Tan, M.S.Y.; Blackman, M.J. The Plasmodium falciparum pseudoprotease SERA5 regulates the kinetics and efficiency of malaria parasite egress from host erythrocytes. PLoS Pathog. 2017, 13, e1006453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackman, M.J. Proteases in host cell invasion by the malaria parasite. Cell Microbiol. 2004, 6, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Buguliskis, J.S.; Lee, T.D.; Sibley, L.D. Functional analysis of rhomboid proteases during Toxoplasma invasion. mBio 2014, 5, e01795-14. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, L.U.; Denise, H.; Coombs, G.H.; Alexander, J.; Mottram, J.C.; Scott, P. Cysteine protease B of Leishmania mexicana inhibits host Th1 responses and protective immunity. J. Immunol. 2003, 171, 3711–3717. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, R.; Das, P.; De, T.; Chakraborti, T. Immunolocalization and characterization of two novel proteases in Leishmania donovani: Putative roles in host invasion and parasite development. Biochimie 2010, 92, 1274–1286. [Google Scholar] [CrossRef]
- Serrano-Luna, J.; Piña-Vázquez, C.; Reyes-López, M.; Ortiz-Estrada, G.; de la Garza, M. Proteases from Entamoeba spp. and pathogenic free-living amoebae as virulence factors. J. Trop. Med. 2013, 2013, 890603. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.A.; Nam, Y.H.; Min, A.; Kim, K.A.; Nozaki, T.; Saito-Nakano, Y.; Mirelman, D.; Shin, M.H. Entamoeba histolytica-secreted cysteine proteases induce IL-8 production in human mast cells via a PAR2-independent mechanism. Parasite 2014, 21, 1. [Google Scholar] [CrossRef] [Green Version]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Betanzos, A.; Bañuelos, C.; Orozco, E. Host invasion by pathogenic amoebae: Epithelial disruption by parasite proteins. Genes 2019, 10, 618. [Google Scholar] [CrossRef] [Green Version]
- Bazzan, E.; Tinè, M.; Casara, A.; Biondini, D.; Semenzato, U.; Cocconcelli, E. Critical review of the evolution of extracellular vesicles’ knowledge: From 1946 to Today. Int. J. Mol. Sci. 2021, 22, 6417. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band a (~140 KDa) | Band b (~100 KDa) | Band c (~80 KDa) | ||||
|---|---|---|---|---|---|---|
| Accession | Description | Accession | Description | Accession | Description | |
| 1 | B5M6J9 | Actin (fragment) | A0A6A5C9S6 | Uncharacterized protein | A0A6A5BE22 | Methylmalonate-semialdehyde dehydrogenase |
| 2 | A0A6A5BG15 | Peptidase_S9 domain-containing protein | B5M6J9 | Actin (fragment) | B5M6J9 | Actin (fragment) |
| 3 | A0A6A5BDZ7 | Uncharacterized protein | A0A6A5BQ42 | Glucose-6-phosphate isomerase | A0A6A5BJW8 | Uncharacterized protein |
| 4 | A0A6A5BS42 | Uncharacterized protein | A0A6A5C1E4 | Uncharacterized protein | A0A6A5C7K8 | Cytosol_AP domain-containing protein |
| 5 | A0A6A5CCP5 | Uncharacterized protein | A0A6A5C7K8 | Cytosol AP domain-containing protein | A0A6A5BFY9 | Peptidase_M28 domain-containing protein |
| 6 | A0A6A5CD16 | VWFA domain-containing protein | A0A6A5C8E9 | Guanine nucleotide-binding protein subunit β-like protein | A0A6A5BNQ4 | Glutamate dehydrogenase |
| 7 | A0A6A5BUK4 | Uncharacterized protein | A0A6A5BXF2 | Histidine ammonia-lyase | A0A6A5BBK6 | Uncharacterized protein |
| 8 | A0A6A5C3B7 | Uncharacterized protein | A0A6A5CD16 | VWFA domain-containing protein | A0A6A5CD16 | VWFA domain-containing protein |
| 9 | A0A6A5BAB4 | Uncharacterized protein | A0A6A5BR20 | Uncharacterized protein | A0A6A5C9S6 | Uncharacterized protein |
| 10 | A0A6A5BQ87 | Uncharacterized protein | A0A6A5BHL2 | CCT-alpha | A0A6A5BJJ6 | Coronin |
| 11 | A0A6A5C150 | C2 domain-containing protein | A0A6A5CCP5 | Uncharacterized protein | A0A6A5C761 | Uncharacterized protein |
| 12 | A0A6A5CG58 | Uncharacterized protein | A0A6A5BT30 | Uncharacterized protein | A0A6A5B5S1 | ADP/ATP translocase |
| 13 | A0A6A5CHB1 | Uncharacterized protein | A0A6A5C7Z0 | Beta-hexosaminidase | A0A6A5CCP5 | Uncharacterized protein |
| 14 | A0A6A5BVL0 | Cytokinin dehydrogenase | A0A1L1XWF9 | Fowlerpain-2 | A0A1L1XWF9 | Fowlerpain-2 |
| 15 | A0A6A5BDH1 | Prolyl endopeptidase | A0A6A5BP20 | Pept_C1 domain-containing protein | A0A6A5BUU2 | Bifunctional dihydrofolate reductase-thymidylate synthase |
| 16 | A0A6A5BXC9 | Peptidase_S9 domain-containing protein | A0A6A5CCE5 | SHOCT domain-containing protein | A0A6A5BUH9 | Dihydrolipoyl dehydrogenase |
| 17 | A0A6A5BF81 | Fructose-bisphosphate aldolase | A0A6A5C1F7 | PRK domain-containing protein | A0A6A5BYK8 | Adenosylhomocysteinase |
| 18 | D2W4E3 | Leucine aminopeptidase ** | D2V4W4 | Mitochondrial chaperonin cpn60 ** | A0A6A5BWP7 | Catalase |
| 19 | A0A6A5BKE4 | Alpha-mannosidase | A0A6A5C3B7 | Uncharacterized protein | A0A6A5BF81 | Fructose-bisphosphate aldolase |
| 20 | A0A6A5BP34 | Uncharacterized protein | A0A6A5B5S1 | ADP/ATP translocase | A0A6A5C539 | WH2 domain-containing protein |
| 21 | A0A6A5C4U4 | Peptidase_M3 domain-containing protein | A0A6A5C052 | Uncharacterized protein | D2V5W7 | Vacuolar proton pump subunit B |
| 22 | A0A6A5C7J0 | Pyruvate carboxylase | A0A6A5C6S5 | M20_dimer domain-containing protein | A0A6A5BFD6 | UDP-glucose 6-dehydrogenase |
| 23 | A0A6A5C9S6 | Uncharacterized protein | A0A6A5BBK6 | Uncharacterized protein | A0A6A5BXJ0 | AMP_N domain-containing protein |
| 24 | A0A1L1XWF9 | Fowlerpain-2 | A0A6A5BSJ3 | Peptidase S53 domain-containing protein | A0A6A5BA58 | Uncharacterized protein |
| 25 | D2VDR3 | Methylcrotonyl-CoA carboxylase ** | A0A6A5BFH5 | Uncharacterized protein | A0A6A5CIW0 | Non-specific serine/threonine protein kinase |
| 26 | A0A6A5C8W6 | Uncharacterized protein | A0A6A5BUK4 | Uncharacterized protein | A0A6A5BWG6 | Uncharacterized protein |
| 27 | A0A4V8H039 | Glyceraldehyde-3-phosphate dehydrogenase | Q6B3P1 | Hsp70 | A0A6A5BYB7 | Uncharacterized protein |
| 28 | A0A6A5CIU9 | Peptidase_S8 domain-containing protein | A0A6A5CFU9 | Peptidase A1 domain-containing protein | D2V4W4 | Mitochondrial chaperonin cpn60 |
| 29 | A0A6A5CBE6 | Uncharacterized protein | Q94626 | Cpn-60 (fragment) | ||
| 30 | A0A6A5CHD7 | Uncharacterized protein | A0A4V8H039 | Glyceraldehyde-3-phosphate dehydrogenase | ||
| 31 | A0A6A5BGJ7 | Imidazolonepropionate hydrolase | A0A6A5CAR5 | Elongation factor 1-alpha | ||
| 32 | A0A6A5C721 | RGS domain-containing protein | A0A6A5C052 | Uncharacterized protein | ||
| 33 | Q25548 | Penicillin amidase homolog (fragment) | A0A6A5BQM0 | Isocitrate dehydrogenase | ||
| 34 | A0A6A5BBK6 | Uncharacterized protein | A0A6A5C3B7 | Uncharacterized protein | ||
| 35 | A0A6A5B2H1 | Lysine--tRNA ligase | A0A6A5BUK8 | Uncharacterized protein | ||
| 36 | A0A6A5BY41 | Uncharacterized protein | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Retana Moreira, L.; Steller Espinoza, M.F.; Chacón Camacho, N.; Cornet-Gomez, A.; Sáenz-Arce, G.; Osuna, A.; Lomonte, B.; Abrahams Sandí, E. Characterization of Extracellular Vesicles Secreted by a Clinical Isolate of Naegleria fowleri and Identification of Immunogenic Components within Their Protein Cargo. Biology 2022, 11, 983. https://doi.org/10.3390/biology11070983
Retana Moreira L, Steller Espinoza MF, Chacón Camacho N, Cornet-Gomez A, Sáenz-Arce G, Osuna A, Lomonte B, Abrahams Sandí E. Characterization of Extracellular Vesicles Secreted by a Clinical Isolate of Naegleria fowleri and Identification of Immunogenic Components within Their Protein Cargo. Biology. 2022; 11(7):983. https://doi.org/10.3390/biology11070983
Chicago/Turabian StyleRetana Moreira, Lissette, María Fernanda Steller Espinoza, Natalia Chacón Camacho, Alberto Cornet-Gomez, Giovanni Sáenz-Arce, Antonio Osuna, Bruno Lomonte, and Elizabeth Abrahams Sandí. 2022. "Characterization of Extracellular Vesicles Secreted by a Clinical Isolate of Naegleria fowleri and Identification of Immunogenic Components within Their Protein Cargo" Biology 11, no. 7: 983. https://doi.org/10.3390/biology11070983
APA StyleRetana Moreira, L., Steller Espinoza, M. F., Chacón Camacho, N., Cornet-Gomez, A., Sáenz-Arce, G., Osuna, A., Lomonte, B., & Abrahams Sandí, E. (2022). Characterization of Extracellular Vesicles Secreted by a Clinical Isolate of Naegleria fowleri and Identification of Immunogenic Components within Their Protein Cargo. Biology, 11(7), 983. https://doi.org/10.3390/biology11070983