
Phenotypic and Genotypic Detection of Biofilm-Forming Staphylococcus aureus from Different Food Sources in Bangladesh


 ,
,  ,
,  , ,
, ,  , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Size Calculation
2.2. Study Area and Sampling
2.3. Sample Processing
2.4. Isolation of S. aureus
2.5. Molecular Detection of S. aureus
2.6. Biofilm Formation of S. aureus
2.6.1. Phenotypic Analysis of Biofilm Formation
2.6.2. Quantitative Analysis of Biofilm Formation
2.6.3. Genotypic Analysis of Biofilm Formation
2.7. Statistical Analysis
2.7.1. Descriptive Analysis
2.7.2. Bivariate Analysis
2.7.3. Heatmap Analysis
3. Results
3.1. Prevalence of S. aureus Isolates
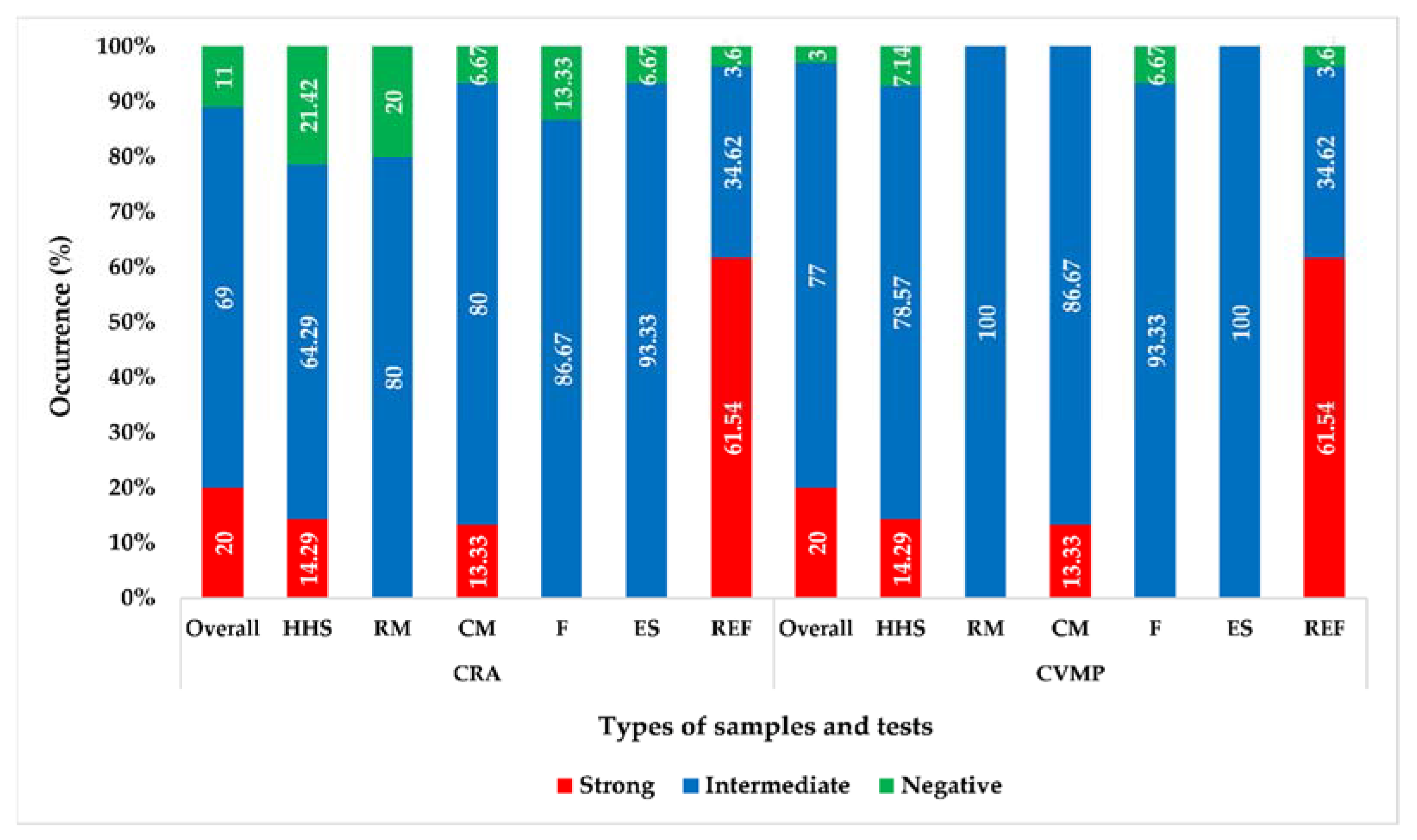
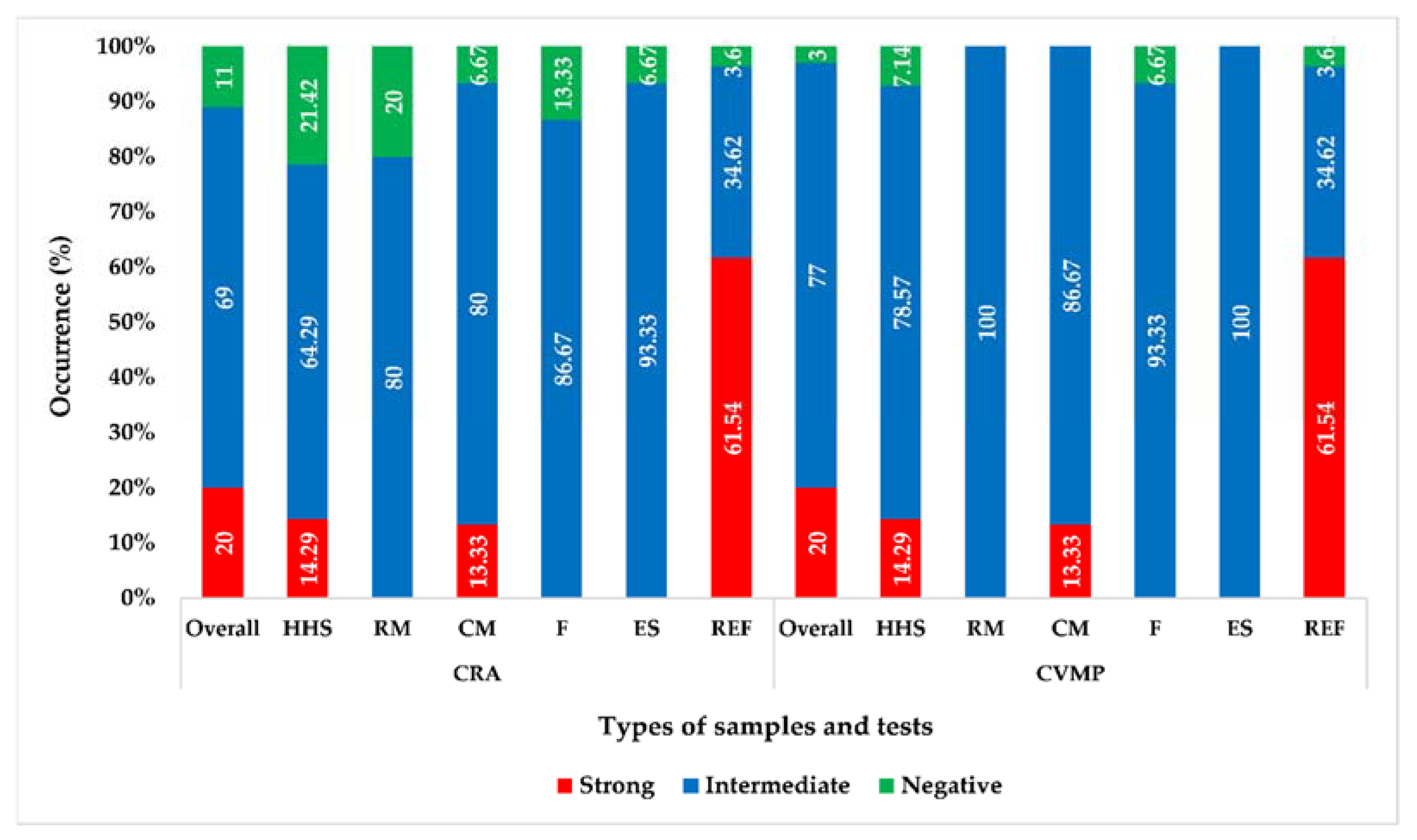
3.2. Phenotypic Biofilm Formation
3.3. Quantification of Biofilm Formation
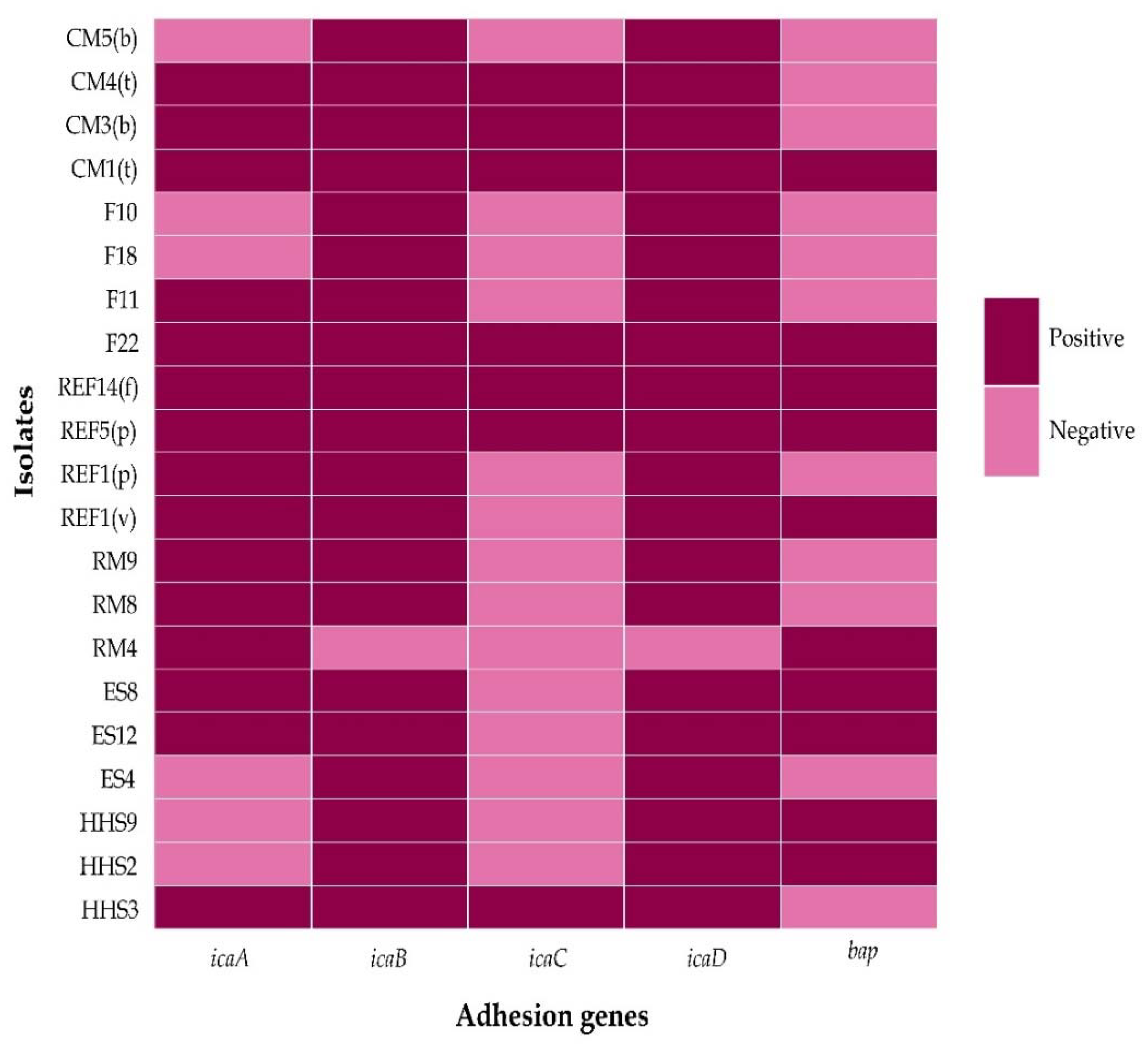
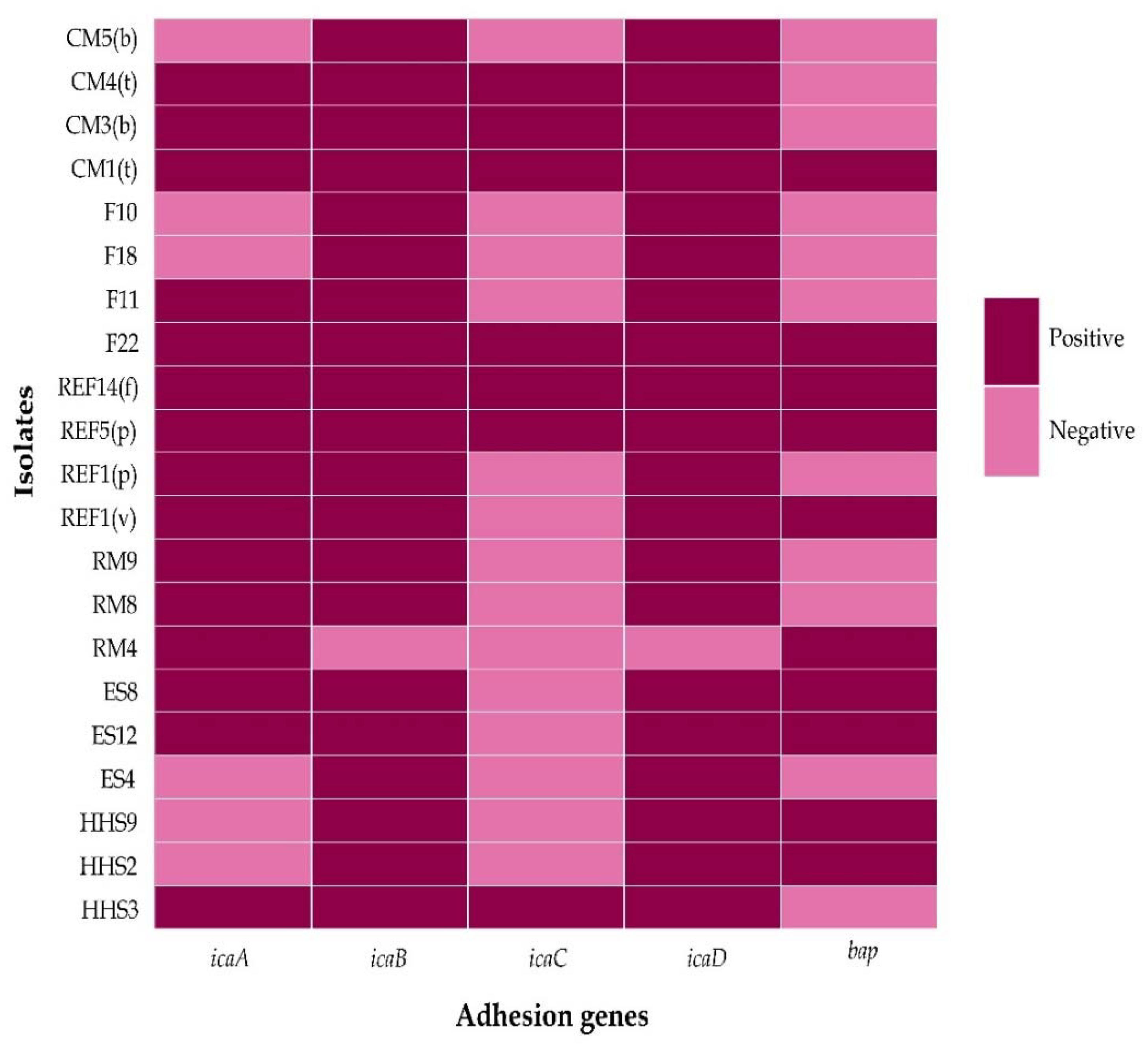
3.4. Genotypic Biofilm Formation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thiran, E.; Di Ciccio, P.; Graber, H.; Zanardi, E.; Ianieri, A.; Hummerjohann., J. Biofilm formation of Staphylococcus aureus dairy isolates representing different genotypes. J. Dairy Sci. 2018, 101, 1000–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urmi, M.R.; Ansari, W.K.; Islam, M.S.; Sobur, M.A.; Rahman, M.; Rahman, M.T. Antibiotic resistance patterns of Staphylococcus spp. isolated from fast foods sold in different restaurants of Mymensingh, Bangladesh. J. Adv. Vet. Anim. Res. 2021, 8, 274. [Google Scholar] [CrossRef]
- Chen, Q.; Xie, S.; Lou, X.; Cheng, S.; Liu, X.; Zheng, W.; Zheng, Z.; Wang, H. Biofilm formation and prevalence of adhesion genes among Staphylococcus aureus isolates from different food sources. Microbiologyopen 2020, 9, e00946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, O.Y.; Raaijmakers, J.M.; Kuramae, E.E. Microbial extracellular polymeric substances: Ecological function and impact on soil aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, E.L.; Meira, Q.G.; Barbosa, I.D.; Athayde, A.J.; Conceição, M.L.; Siqueira Júnior, J.P. Biofilm formation by Staphylococcus aureus from food contact surfaces in a meat-based broth and sensitivity to sanitizers. Braz. J. Microbiol. 2014, 45, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaguler, T.; Kahraman, H.; Tuter, M. Analyzing effects of ELF electromagnetic fields on removing bacterial biofilm. Biocybern. Biomed. Eng. 2017, 37, 336–340. [Google Scholar] [CrossRef]
- Payne, D.E.; Boles, B.R. Emerging interactions between matrix components during biofilm development. Curr. Genet. 2016, 62, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Lin, S.; Soteyome, T.; Peters, B.M.; Li, Y.; Chen, H.; Su, J.; Li, L.; Li, B.; Xu, Z. Biofilm formation of Staphylococcus aureus under food heat processing conditions: First report on CML production within biofilm. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Xie, J.; Peters, B.M.; Li, B.; Li, L.; Yu, G.; Xu, Z.; Shirtliff, M.E. Clinical features and antimicrobial resistance profiles of important Enterobacteriaceae pathogens in Guangzhou representative of Southern China, 2001–2015. Microb. Pathog. 2017, 107, 206–211. [Google Scholar] [CrossRef]
- Avila-Novoa, M.-G.; Iñíguez-Moreno, M.; Solís-Velázquez, O.-A.; González-Gomez, J.-P.; Guerrero-Medina, P.-J.; Gutiérrez-Lomelí, M.J. Biofilm formation by Staphylococcus aureus isolated from food contact surfaces in the dairy industry of Jalisco, Mexico. J. Food Qual. 2018, 2018, 1746139. [Google Scholar] [CrossRef] [Green Version]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation, and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thrusfeld, M. Veterinary Epidemiology, 2nd ed.; Blackwell Science Ltd.: London, UK, 1995; p. 83. [Google Scholar]
- Sobur, M.; Islam, M.; Haque, Z.F.; Orubu, E.S.F.; Toniolo, A.; Choudhury, M.; Rahman, M. Higher seasonal temperature enhances the occurrence of methicillin resistance of Staphylococcus aureus in house flies (Musca domestica) under hospital and environmental settings. Folia Microbiol. 2022, 67, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Kalorey, D.R.; Shanmugam, Y.; Kurkure, N.V.; Chousalkar, K.K.; Barbuddhe, S.B. PCR-based detection of genes encoding virulence determinants in Staphylococcus aureus from bovine subclinical mastitis cases. J. Vet. Sci. 2007, 8, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Rohde, H.; Knobloch, J.K.; Horstkotte, M.A.; Mack, D. Correlation of Staphylococcus aureus icaADBC genotype and biofilm expression phenotype. J. Clin. Microbiol. 2001, 39, 4595–4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Xu, Y.; Yestrepsky, B.D.; Sorenson, R.J.; Chen, M.; Larsen, S.D.; Sun, H. Novel inhibitors of Staphylococcus aureus virulence gene expression and biofilm formation. PLoS ONE 2012, 7, e47255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiem, S.; Oh, W.S.; Peck, K.R.; Lee, N.Y.; Lee, J.-Y.; Song, J.-H.; Hwang, E.S.; Kim, E.-C.; Cha, C.Y.; Choe, K.-W. Phase variation of biofilm formation in Staphylococcus aureus by IS256 insertion and its impact on the capacity adhering to polyurethane surface. J. Korean Med. Sci. 2004, 19, 779–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucarella, C.; Tormo, M.A.; Ubeda, C.; Trotonda, M.P.; Monzón, M.; Peris, C.; Amorena, B.; Lasa, Í.; Penadés, J.R. Role of biofilm-associated protein bap in the pathogenesis of bovine Staphylococcus Aureus. Infect. Immun. 2004, 72, 2177–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.; Nayeem, M.; Hasan, M.; Sobur, M.; Ievy, S.; Rahman, S.; Kafi, M.; Ashour, H.M.; Rahman, M. Virulence determinants and multidrug resistance of Escherichia coli isolated from migratory birds. Antibiotics 2021, 10, 190. [Google Scholar] [CrossRef]
- Tawyabur, M.; Islam, M.; Sobur, M.; Hossain, M.; Mahmud, M.; Paul, S.; Hossain, M.T.; Ashour, H.M.; Rahman, M. Isolation and characterization of multidrug-resistant Escherichia coli and Salmonella spp. from healthy and diseased turkeys. Antibiotics 2020, 9, 770. [Google Scholar] [CrossRef]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. Presence of icaA and icaD genes and slime production in a collection of staphylococcal strains from catheter-associated infections. J. Clin. Microbiol. 2001, 39, 2151–2156. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Campoccia, D.; Gamberini, S.; Baldassarri, L.; Montanaro, L. Prevalence of can, fnbA and fnbB adhesin genes among Staphylococcus aureus isolates from orthopedic infections associated to different types of implant. FEMS Microbiol. Lett. 2005, 246, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouidhi, B.; Zmantar, T.; Hentati, H.; Bakhrouf, A. Cell surface hydrophobicity, biofilm formation, adhesives properties and molecular detection of adhesins genes in Staphylococcus aureus associated to dental caries. Microb. Pathog. 2010, 49, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.D.; Cai, T.T.; DasGupta, A. Interval estimation for a binomial proportion. Stat. Sci. 2001, 16, 101–133. [Google Scholar] [CrossRef]
- Rahman, M.; Sobur, M.; Islam, M.; Ievy, S.; Hossain, M.; El Zowalaty, M.E.; Rahman, A.; Ashour, H.M. Zoonotic diseases: Etiology, impact, and control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef]
- Islam, M.A.; Parveen, S.; Rahman, M.; Huq, M.; Nabi, A.; Khan, Z.U.M.; Ahmed, N.; Wagenaar, J.A. Occurrence and characterization of methicillin resistant Staphylococcus aureus in processed raw foods and ready-to-eat foods in an urban setting of a developing country. Front. Microbiol. 2019, 10, 503. [Google Scholar] [CrossRef]
- Jahan, M.; Rahman, M.; Parvej, M.S.; Chowdhury, S.M.Z.H.; Haque, M.E.; Talukder, M.A.K.; Ahmed, S. Isolation and characterization of Staphylococcus aureus from raw cow milk in Bangladesh. J. Adv. Vet. Anim. Res. 2015, 2, 49–55. [Google Scholar] [CrossRef]
- Pondit, A.; Haque, Z.F.; Sabuj, A.A.M.; Khan, M.S.R.; Saha, S. Characterization of Staphylococcus aureus isolated from chicken and quail eggshell. J. Adv. Vet. Anim. Res. 2018, 5, 466. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, J.; Wu, Q.; Zhang, J.; Zhang, F.; Yang, X.; Wu, H.; Zeng, H.; Chen, M.; Ding, Y. Staphylococcus aureus isolated from retail meat and meat products in China: Incidence, antibiotic resistance and genetic diversity. Front. Microbiol. 2018, 9, 2767. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Zhang, F.; Wu, Q.; Zhang, J.; Pang, R.; Zeng, H.; Yang, X.; Chen, M.; Wang, J. Prevalence and characterization of food-related methicillin-resistant Staphylococcus aureus (MRSA) in China. Front. Microbiol. 2019, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Regasa, S.; Mengistu, S.; Abraha, A. Milk safety assessment, isolation, and antimicrobial susceptibility profile of Staphylococcus aureus in selected dairy farms of mukaturi and sululta town, oromia region, Ethiopia. Vet. Med. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Kukułowicz, A.; Steinka, I.; Siwek, A. Presence of antibiotic-resistant Staphylococcus aureus in fish and seafood originating from points of sale in the tri-city area (Poland). J. Food Prot. 2021, 84, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Shaltout, F.A.E.; Farouk, M.; Ibrahim, H.A.; Afifi, M.E. Incidence of coliform and Staphylococcus aureus in ready to eat fast foods. Benha Vet. Med. J. 2017, 32, 13–17. [Google Scholar] [CrossRef]
- Syed, M.A.; Shah, S.H.H.; Sherafzal, Y.; Shafi-ur-Rehman, S.; Khan, M.A.; Barrett, J.B.; Woodley, T.A.; Jamil, B.; Abbasi, S.A.; Jackson, C.R. Detection and molecular characterization of methicillin-resistant Staphylococcus aureus from table eggs in Haripur, Pakistan. Foodborne Pathog. Dis. 2018, 15, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Gadaga, T.H.; Samende, B.K.; Musuna, C.; Chibanda, D. The microbiological quality of informally vended foods in Harare, Zimbabwe. Food Control. 2008, 19, 829–832. [Google Scholar] [CrossRef]
- Kasturwar, N.B.; Shafee, M. Knowledge, practices and prevalence of MRSA among food handlers. Int. J. Biol. Med. Res. 2011, 2, 889–894. [Google Scholar]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Torlak, E.; Korkut, E.; Uncu, A.T.; Şener, Y. Biofilm formation by Staphylococcus aureus isolates from a dental clinic in Konya, Turkey. J. Infect. Public Health 2017, 10, 809–813. [Google Scholar] [CrossRef]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Høiby, N. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 2002, 148, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, T.D.L.; Pereira, E.M.; Dos Santos, K.R.N.; Maciel, E.L.N.; Schuenck, R.P.; Nunes, A.P.F. Modification of the Congo red agar method to detect biofilm production by Staphylococcus epidermidis. Diagn. Microbiol. Infect. Dis. 2013, 75, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Kroning, I.S.; Iglesias, M.A.; Sehn, C.P.; Gandra, T.K.V.; Mata, M.M.; da Silva, W.P. Staphylococcus aureus isolated from handmade sweets: Biofilm formation, enterotoxigenicity and antimicrobial resistance. Food Microbiol. 2016, 58, 105–111. [Google Scholar] [CrossRef]
- Atshan, S.S.; Shamsudin, M.N.; Karunanidhi, A.; van Belkum, A.; Lung, L.T.T.; Sekawi, Z.; Nathan, J.J.; Ling, K.H.; Seng, J.S.C.; Ali, A.M. Quantitative PCR analysis of genes expressed during biofilm development of methicillin resistant Staphylococcus aureus (MRSA). Infect. Genet. Evol. 2013, 18, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Stanley, N.R.; Lazazzera, B.A. Environmental signals and regulatory pathways that influence biofilm formation. Mol. Microbiol. 2004, 52, 917–924. [Google Scholar] [CrossRef]
- El-Nagdy, A.H.; Abdel-Fattah, G.M.; Emarah, Z. Detection and control of biofilm formation by Staphylococcus aureus from febrile neutropenic patient. Infect. Drug Resist. 2020, 13, 3091. [Google Scholar] [CrossRef] [PubMed]
- Szczuka, E.; Urbańska, K.; Pietryka, M.; Kaznowski, A. Biofilm density and detection of biofilm-producing genes in methicillin-resistant Staphylococcus aureus strains. Folia Microbiol. 2013, 58, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Chen, J.; Li, H.; Zeng, P.; Li, J. Characterization of adhesin genes, staphylococcal nuclease, hemolysis, and biofilm formation among Staphylococcus aureus strains isolated from different sources. Foodborne Pathog. Dis. 2013, 10, 757–763. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Delgado, S.; Vázquez-Sánchez, D.; Martínez, B.; Cabo, M.L.; Rodríguez, A.; Herrera, J.J.; García, P. Incidence of Staphylococcus aureus and analysis of associated bacterial communities on food industry surfaces. Appl. Environ. Microbiol. 2012, 78, 8547–8554. [Google Scholar] [CrossRef] [Green Version]
- Beenken, K.E.; Dunman, P.M.; McAleese, F.; Macapagal, D.; Murphy, E.; Projan, S.J.; Blevins, J.S.; Smeltzer, M.S. Global gene expression in Staphylococcus aureus biofilms. J. Bacteriol. 2004, 186, 4665–4684. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Kim, C.-H.; Hacker, J.; Ziebuhr, W.; Lee, B.-K.; Cho, S.-h. Molecular characterization of regulatory genes associated with biofilm variation in a Staphylococcus aureus strain. J. Microbiol. Biotechnol. 2008, 18, 28–34. [Google Scholar]
- O’Gara, J.P. Ica and beyond: Biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Ziebuhr, W.; Krimmer, V.; Rachid, S.; Lößner, I.; Götz, F.; Hacker, J. A novel mechanism of phase variation of virulence in Staphylococcus epidermidis: Evidence for control of the polysaccharide intercellular adhesin synthesis by alternating insertion and excision of the insertion sequence element IS256. Mol. Microbiol. 1999, 32, 345–356. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Targeted Genes | Primer Sequence (5′–3′) | Annealing Temperature | Amplicon Size (bp) | References |
|---|---|---|---|---|
| nuc | F: GCGATTGATGGTGATACGGT R: AGCCAAGCCTTGACGAACTAAAGC | 55 | 279 | [14] |
| icaA | F: GACCTCGAAGTCAATAGAGGT R: CCCAGTATAACGTTGGATACC | 56 | 814 | [15] |
| icaD | F: AGGCAATATCCAACGGTAA R: GTCACGACCTTTCTTATATT | 59 | 526 | [16] |
| icaB | F: ATCGCTTAAAGCACACGACGC R: TATCGGCATCTGGTGTGACAG | 59 | 526 | [17] |
| icaC | F: ATAAACTTGAATTAGTGTATT R: ATATATAAAACTCTCTTAACA | 45 | 989 | [17] |
| bap | F: CCCTATATCGAAGGTGTAGAATTGCAC R: GCTGTTGAAGTTAATACTGTACCTGC | 53 | 971 | [18] |
| Name of Sample | Positive Isolates (%) | 95% CI | p-Value |
|---|---|---|---|
| Human hand swab (n = 60) | 14 (23.33) | 14.44–35.44 | 0.992 |
| Raw milk (n = 60) | 15 (25) | 15.78–37.23 | |
| Chicken muscle (n = 60) | 15 (25) | 15.78–37.23 | |
| Fish (n = 60) | 15 (25) | 15.78–37.23 | |
| Egg surface (n = 60) | 15 (25) | 15.78–37.23 | |
| Ready-to-eat foods (n = 120) | 26 (21.67) | 15.24–29.85 | |
| Overall (n = 420) | 100 (23.81) | 19.99–28.11 |
| icaA | icaB | icaC | icaD | bap | ||
|---|---|---|---|---|---|---|
| icaA | Pearson Correlation | 1 | ||||
| Sig. (2-tailed) | - | |||||
| icaB | Pearson Correlation | 0.770 ** | 1 | |||
| Sig. (2-tailed) | 0.000 | - | ||||
| icaC | Pearson Correlation | 0.653 ** | 0.549 ** | 1 | ||
| Sig. (2-tailed) | 0.000 | 0.000 | - | |||
| icaD | Pearson Correlation | 0.770 ** | 1.000 ** | 0.549 ** | 1 | |
| Sig. (2-tailed) | 0.000 | 0.000 | 0.000 | - | ||
| bap | Pearson Correlation | 0.607 ** | 0.583 ** | 0.431 ** | 0.583 ** | 1 |
| Sig. (2-tailed) | 0.000 | 0.000 | 0.000 | 0.000 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballah, F.M.; Islam, M.S.; Rana, M.L.; Ferdous, F.B.; Ahmed, R.; Pramanik, P.K.; Karmoker, J.; Ievy, S.; Sobur, M.A.; Siddique, M.P.; et al. Phenotypic and Genotypic Detection of Biofilm-Forming Staphylococcus aureus from Different Food Sources in Bangladesh. Biology 2022, 11, 949. https://doi.org/10.3390/biology11070949
Ballah FM, Islam MS, Rana ML, Ferdous FB, Ahmed R, Pramanik PK, Karmoker J, Ievy S, Sobur MA, Siddique MP, et al. Phenotypic and Genotypic Detection of Biofilm-Forming Staphylococcus aureus from Different Food Sources in Bangladesh. Biology. 2022; 11(7):949. https://doi.org/10.3390/biology11070949
Chicago/Turabian StyleBallah, Fatimah Muhammad, Md. Saiful Islam, Md. Liton Rana, Farhana Binte Ferdous, Rokeya Ahmed, Pritom Kumar Pramanik, Jarna Karmoker, Samina Ievy, Md. Abdus Sobur, Mahbubul Pratik Siddique, and et al. 2022. "Phenotypic and Genotypic Detection of Biofilm-Forming Staphylococcus aureus from Different Food Sources in Bangladesh" Biology 11, no. 7: 949. https://doi.org/10.3390/biology11070949
APA StyleBallah, F. M., Islam, M. S., Rana, M. L., Ferdous, F. B., Ahmed, R., Pramanik, P. K., Karmoker, J., Ievy, S., Sobur, M. A., Siddique, M. P., Khatun, M. M., Rahman, M., & Rahman, M. T. (2022). Phenotypic and Genotypic Detection of Biofilm-Forming Staphylococcus aureus from Different Food Sources in Bangladesh. Biology, 11(7), 949. https://doi.org/10.3390/biology11070949