
Realising the Environmental Potential of Vertical Farming Systems through Advances in Plant Photobiology

 ,
, 
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
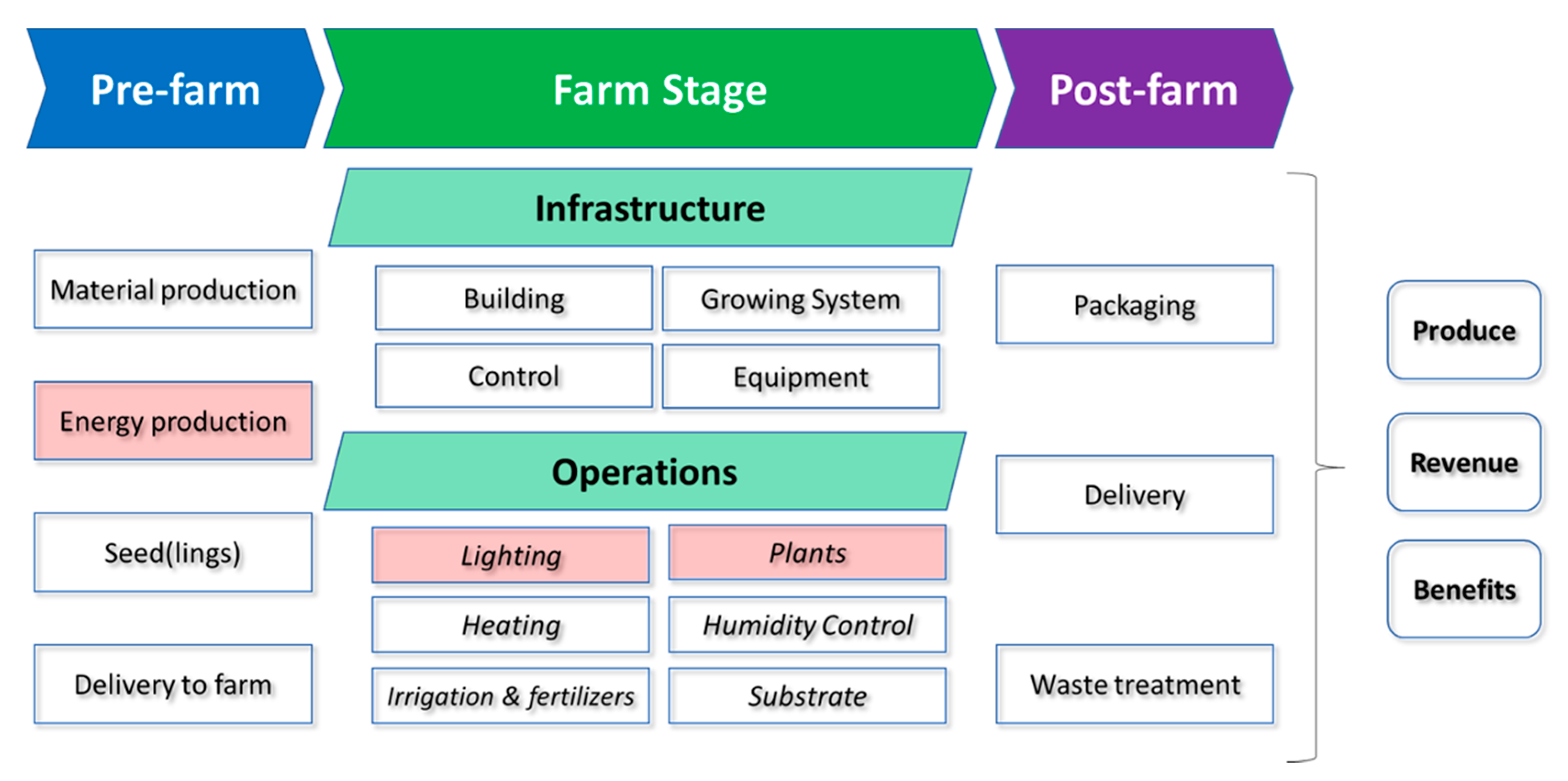
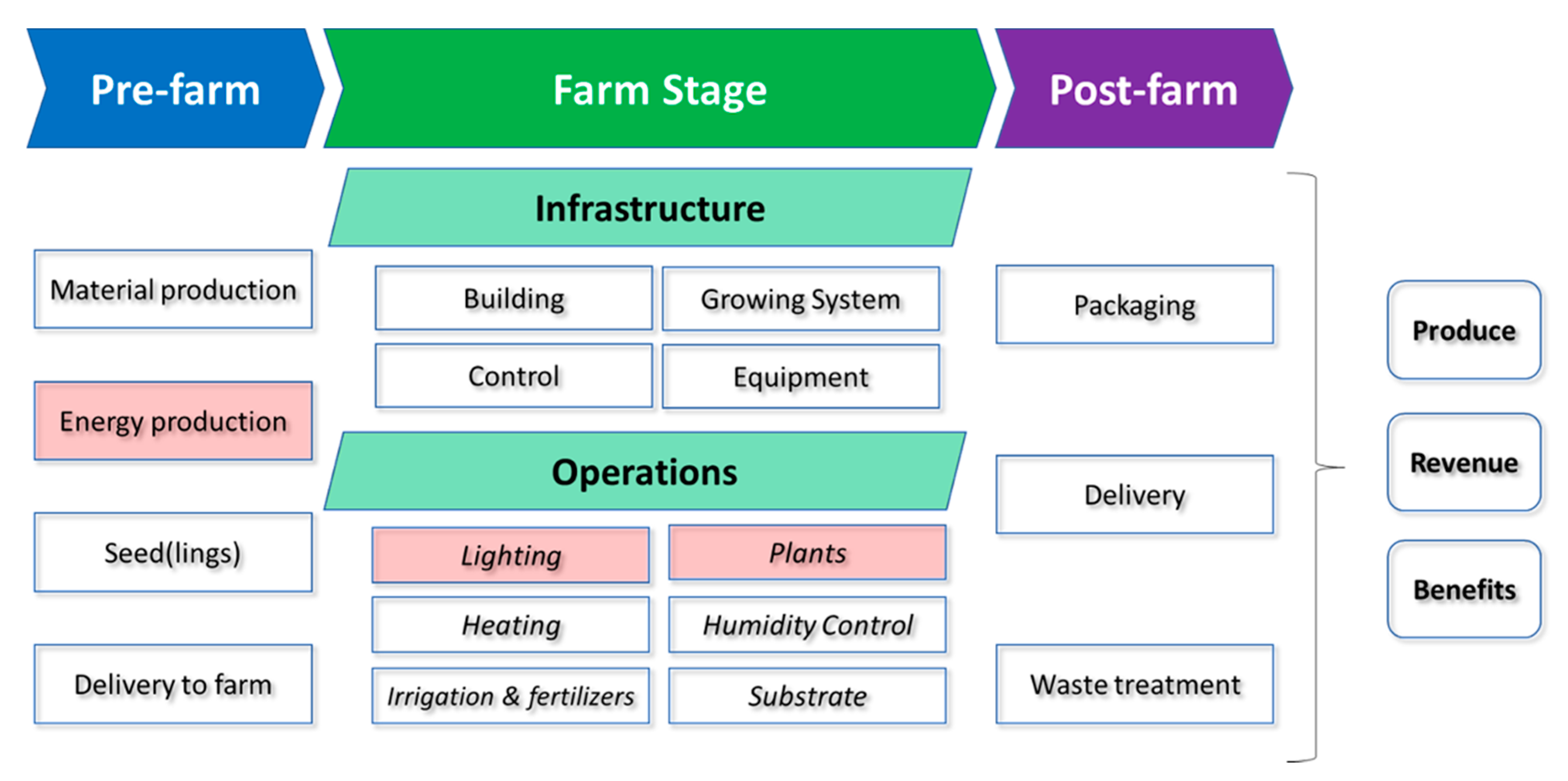
1.1. Defining Sources of Energy Usage and Carbon Emissions in VFS
1.2. Electrification of VFS by Renewable Energy Sources Provides an Opportunity to Contribute Sustainably to Food Production
2. Photobiology Strategies for Maximising Energy Efficiency and Yield
2.1. Daily and Seasonal Variations in Light Irradiation Are an Intrinsic Component of Plant Biology
2.1.1. Light Intensity
2.1.2. Light Quality
2.1.3. Photoperiod and the Control of Flowering
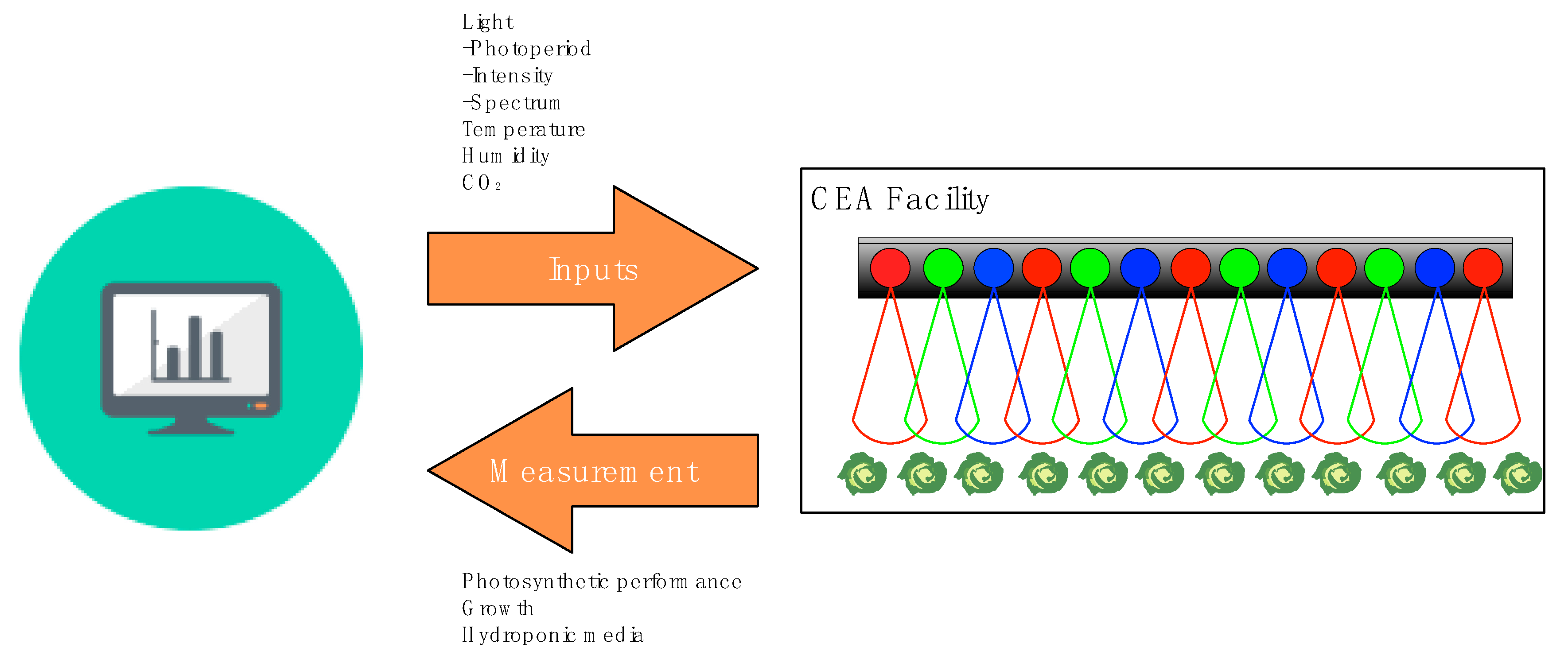
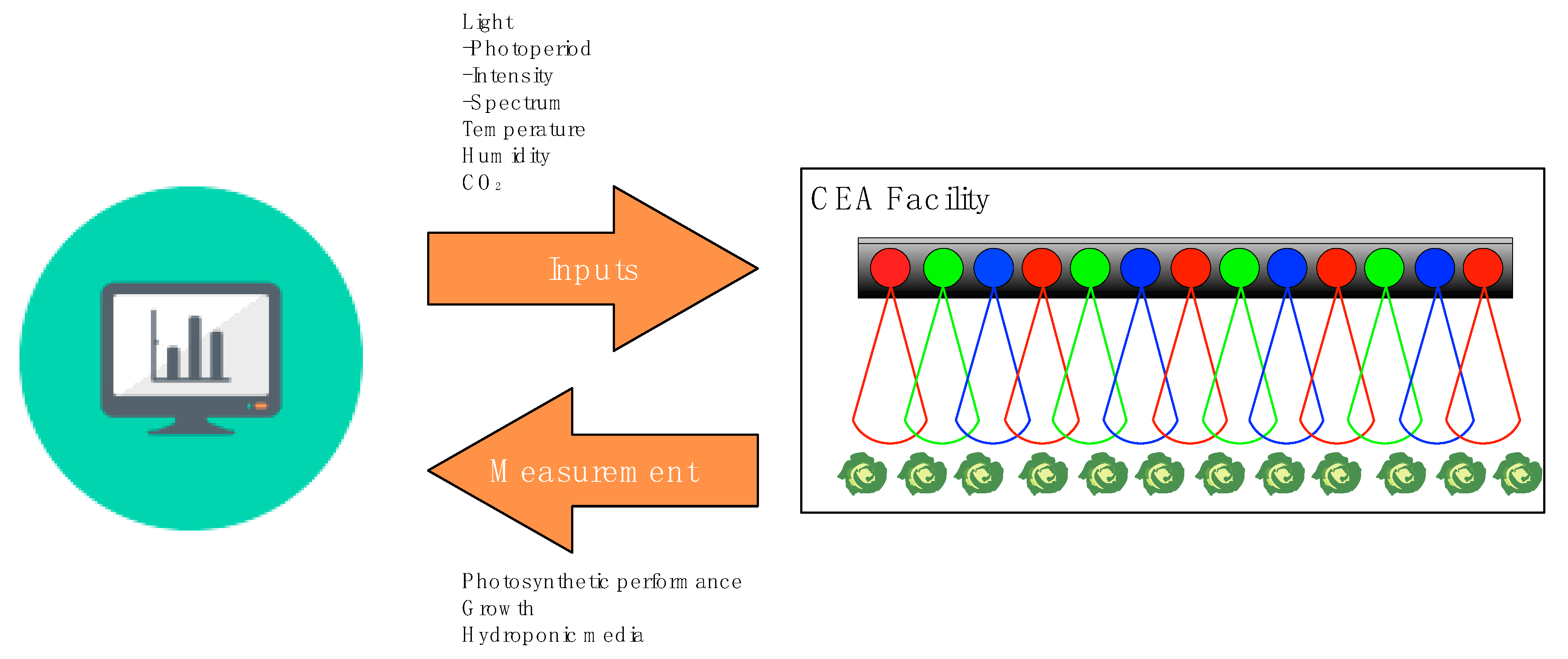
2.2. Variable Lighting with Consideration of Biological Parameters Provides Opportunities to Coordinate Energy Loads with Renewable Energy Grids
2.3. Selective Breeding Provides Opportunities to Optimise Crops for VFS
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The share of food systems in total greenhouse gas emissions. Global, regional and country trends, 1990–2019. In Analytical Brief Series No. 31; FAOSTAT, Ed.; FAO: Rome, Italy, 2021. [Google Scholar]
- UK Consumers Want the Future of Agriculture Discussed at COP26 in Glasgow. Available online: https://www.agriland.co.uk/farming-news/uk-consumers-want-the-future-of-agriculture-discussed-at-cop26-in-glasgow/ (accessed on 15 June 2022).
- UNDP. United Nations Development Programme Report; UNDP: New York, NY, USA, 2019. [Google Scholar]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating Targets for Sustainable Intensification. Bioscience 2017, 67, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Ellingsen, E. The Vertical Farm—The origin of a 21st century Architectural Typology. CTBUH J. 2008, 3, 26–34. [Google Scholar]
- Forbes, H.; Fisher, K.; Parry, A. UK Food System GHG Emissions: Total UK Food & Drink Consumption Footprint and Pathway to a 50% Reduction by 2030; WRAP: Banbury, UK, 2021. [Google Scholar]
- UK-Government. 2021 Government Greenhouse Gas Conversion Factors for Company Reporting; E.I.S. Department for Business: London, UK, 2021. [Google Scholar]
- Noguer, C.G. AgriFood Innovation Helping to Mitigate the Impact of Climate Change; UKRI: Swindon, UK, 2022. [Google Scholar]
- van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; et al. Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef]
- Graamans, L.; Baeza, E.; van den Dobbelsteen, A.; Tsafaras, I.; Stanghellini, C. Plant factories versus greenhouses: Comparison of resource use efficiency. Agric. Syst. 2018, 160, 31–43. [Google Scholar] [CrossRef]
- UN-WUP. World Urbanization Prospects—The 2018 Revision; United Nations: New York, NY, USA, 2018. [Google Scholar]
- Kalantari, F.; Tahir, O.M.; Joni, R.A.; Fatemi, E. Opportunities and Challenges in Sustainability of Vertical Farming: A Review. J. Landsc. Ecol. 2018, 11, 35–60. [Google Scholar] [CrossRef] [Green Version]
- WayBeyond Ltd.; Agritecture LLC. 2021 Global CEA Census Report; WayBeyond Ltd.: Auckland, New Zealand; Agritecture LLC: Auckland, New Zealand, 2021. [Google Scholar]
- Gordon-Smith, H. Vertical Farming Is Headed for the ‘Trough of Disillusionment’. Here’s Why That’s a Good Thing. Available online: https://agfundernews.com/vertical-farming-is-headed-for-the-trough-of-disillusionment-heres-why-thats-a-good-thing/ (accessed on 15 June 2022).
- ISO 14040: 2006; Environmental Management—Life Cycle Assessment—Principles and Framework. International Standards Organization: Geneva, Switzerland, 2006.
- GHGprotocol. The Greenhouse Gas Protocol—A Corporate Accounting and Reporting Standard; World Business Council for Sustainable Development, Ed.; World Resources Institute, World Business Council for Sustainable Development: Geneva, Switzerland, 2004. [Google Scholar]
- Dorr, E.; Goldstein, B.; Horvath, A.; Aubry, C.; Gabrielle, B. Environmental impacts and resource use of urban agriculture: A systematic review and meta-analysis. Environ. Res. Lett. 2021, 16, 093002. [Google Scholar] [CrossRef]
- Jack, L.; Florez-Lopez, R.; Ramon-Jeronimo, J.M. Accounting, performance measurement and fairness in UK fresh produce supply networks. Account. Organ. Soc. 2018, 64, 17–30. [Google Scholar] [CrossRef] [Green Version]
- RICS. Whole Life Carbon Assessment for the Built Environment; Royale Institution of Chartered Surveyors (RICS): London, UK, 2018. [Google Scholar]
- AUK. Agriculture in the United Kingdom; Department for Environment, Food & Rural Affairs: London, UK, 2020. [Google Scholar]
- 2020 Greenest Year on Record for Britain. Available online: https://www.nationalgrid.com/stories/journey-to-net-zero-stories/2020-greenest-year-record-britain (accessed on 15 June 2022).
- OneFarm. CO2 Emission Scoping Report. Comparison between Different Farming Methods in Lettuce Production; OneFarm Ltd.: London, UK, 2018. [Google Scholar]
- Bharathi, C.; Rekha, D.; Vijayakumar, V. Genetic Algorithm Based Demand Side Management for Smart Grid. Wirel. Pers. Commun. 2017, 93, 481–502. [Google Scholar] [CrossRef]
- Castro, P.M.; Sun, L.; Harjunkoski, I. Resource–Task Network Formulations for Industrial Demand Side Management of a Steel Plant. Ind. Eng. Chem. Res. 2013, 52, 13046–13058. [Google Scholar] [CrossRef] [Green Version]
- Gahm, C.; Denz, F.; Dirr, M.; Tuma, A. Energy-efficient scheduling in manufacturing companies: A review and research framework. Eur. J. Oper. Res. 2016, 248, 744–757. [Google Scholar] [CrossRef]
- Kumar, T.S.; Venkatesan, T. A Survey on Demand Response in Smart Power Distribution Systems. In Proceedings of the International Conference on Power, Energy, Control and Transmission Systems (ICPECTS), Chennai, India, 10–11 December 2020. [Google Scholar]
- Shoreh, M.H.; Siano, P.; Shafie-Khah, M.; Loia, V.; Catalão, J.P.S. A survey of industrial applications of Demand Response. Electr. Power Syst. Res. 2016, 141, 31–49. [Google Scholar] [CrossRef]
- Avgoustaki, D.D.; Xydis, G. How energy innovation in indoor vertical farming can improve food security, sustainability, and food safety? Adv. Food Secur. Sustain. 2020, 5, 1–51. [Google Scholar]
- Millar, A.J. The Intracellular Dynamics of Circadian Clocks Reach for the Light of Ecology and Evolution. Annu. Rev. Plant Biol. 2016, 67, 595–618. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.A. Using light to improve commercial value. Hortic. Res. 2018, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Steed, G.; Ramirez, D.C.; Hannah, M.A.; Webb, A.A.R. Chronoculture, harnessing the circadian clock to improve crop yield and sustainability. Science 2021, 372, eabc9141. [Google Scholar] [CrossRef]
- Aphalo, P.J.; Kotilainen, T. The best light spectrum for plant cultivation: A source of colourful arguments. EcoEvoRxiv 2022. [Google Scholar] [CrossRef]
- Kelly, N.; Choe, D.; Meng, Q.; Runkle, E.S. Promotion of lettuce growth under an increasing daily light integral depends on the combination of the photosynthetic photon flux density and photoperiod. Sci. Hortic. 2020, 272, 109565. [Google Scholar] [CrossRef]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Paucek, I.; Appolloni, E.; Pennisi, G.; Quaini, S.; Gianquinto, G.; Orsini, F. LED Lighting Systems for Horticulture: Business Growth and Global Distribution. Sustainability 2020, 12, 7516. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; He, D.; Niu, G.; Yan, Z.; Song, J. Effects of environment lighting on the growth, photosynthesis, and quality of hydroponic lettuce in a plant factory. Int. J. Agric. Biol. Eng. 2018, 11, 33–40. [Google Scholar] [CrossRef]
- Cheng, M.C.; Kathare, P.K.; Paik, I.; Huq, E. Phytochrome Signaling Networks. Annu. Rev. Plant Biol. 2021, 72, 217–244. [Google Scholar] [CrossRef]
- Christie, J.M. Phototropin Blue-Light Receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Lin, C. Mechanisms of Cryptochrome-Mediated Photoresponses in Plants. Annu. Rev. Plant Biol. 2020, 71, 103–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, S.; van Iersel, M.; Bugbee, B. Why Far-Red Photons Should Be Included in the Definition of Photosynthetic Photons and the Measurement of Horticultural Fixture Efficacy. Front. Plant Sci. 2021, 12, 693445. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.I. Photomorphogenic responses to ultraviolet-B light. Plant Cell Environ. 2017, 40, 2544–2557. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, O.K. Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf Morphology, Photosynthetic Performance, Chlorophyll Fluorescence, Stomatal Development of Lettuce (Lactuca sativa L.) Exposed to Different Ratios of Red Light to Blue Light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-L.; Li, Y.-L.; Wang, L.-C.; Guo, W.-Z. Red and blue wavelengths affect the morphology, energy use efficiency and nutritional content of lettuce (Lactuca sativa L.). Sci. Rep. 2021, 11, 8374. [Google Scholar] [CrossRef]
- Pennisi, G.; Orsini, F.; Blasioli, S.; Cellini, A.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Resource use efficiency of indoor lettuce (Lactuca sativa L.) cultivation as affected by red:blue ratio provided by LED lighting. Sci. Rep. 2019, 9, 14127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Shi, Y.; Piao, F.; Sun, Z. Effects of different LED sources on the growth and nitrogen metabolism of lettuce. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 231–240. [Google Scholar] [CrossRef]
- Battle, M.W.; Vegliani, F.; Jones, M.A. Shades of green: Untying the knots of green photoperception. J. Exp. Bot. 2020, 71, 5764–5770. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.; Cheng, R.; Wang, Y.; Yang, Q.; Lu, C. Effect of green light on nitrate reduction and edible quality of hydroponically grown lettuce (Lactuca sativa L.) under short-term continuous light from red and blue light-emitting diodes. Environ. Exp. Bot. 2018, 153, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.; Yang, Q.; Li, T.; Cheng, R.; Barnett, Y.; Lu, C. Study of the beneficial effects of green light on lettuce grown under short-term continuous red and blue light-emitting diodes. Physiol. Plant. 2018, 164, 226–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.L.; McAusland, L.; Murchie, E. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Urbina, J.L.; Heuvelink, E.; Marcelis, L.F.M. Adding Far-Red to Red-Blue Light-Emitting Diode Light Promotes Yield of Lettuce at Different Planting Densities. Front. Plant Sci. 2021, 11, 609977. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and physiological properties of indoor cultivated lettuce in response to additional far-red light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Nhi, P.N. Effects of Color Led Light Intensities and Different Photoperiod Regimes on Growth of Hydroponic Lettuce (Latuca sativa L.). Can Tho Univ. J. Sci. 2016, 2, 1–7. [Google Scholar]
- Zha, L.; Zhang, Y.; Liu, W. Dynamic Responses of Ascorbate Pool and Metabolism in Lettuce to Long-term Continuous Light Provided by Red and Blue LEDs. Environ. Exp. Bot. 2019, 163, 15–23. [Google Scholar] [CrossRef]
- Zhou, W.L.; Liu, W.K.; Yang, Q.C. Quality changes in hydroponic lettuce grown under pre-harvest short-duration continuous light of different intensities. J. Hortic. Sci. Biotechnol. 2015, 87, 429–434. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanoue, J.; Zheng, J.; Little, C.; Thibodeau, A.; Grodzinski, B.; Hao, X. Alternating Red and Blue Light-Emitting Diodes Allows for Injury-Free Tomato Production with Continuous Lighting. Front. Plant Sci. 2019, 10, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgoustaki, D.D.; Bartzanas, T.; Xydis, G. Minimising the energy footprint of indoor food production while maintaining a high growth rate: Introducing disruptive cultivation protocols. Food Control 2021, 130, 108290. [Google Scholar] [CrossRef]
- Palmer, S.; van Iersel, M.W. Increasing Growth of Lettuce and Mizuna under Sole-Source LED Lighting Using Longer Photoperiods with the Same Daily Light Integral. Agronomy 2020, 10, 1659. [Google Scholar] [CrossRef]
- Rajendran, S.; Heo, J.; Kim, Y.; Kim, D.; Ko, K.; Lee, Y.; Oh, S.; Kim, C.; Bae, J.; Park, S. Optimization of Tomato Productivity Using Flowering Time Variants. Agronomy 2021, 11, 285. [Google Scholar] [CrossRef]
- Leijten, W.; Koes, R.; Roobeek, I.; Frugis, G. Translating Flowering Time from Arabidopsis thaliana to Brassicaceae and Asteraceae Crop Species. Plants 2018, 7, 111. [Google Scholar] [CrossRef] [Green Version]
- Jones, M. Entrainment of the Arabidopsis circadian clock. J. Plant Biol. 2009, 52, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Eaves, J.; Eaves, S. Comparing the Profitability of a Greenhouse to a Vertical Farm in Quebec. Can. J. Agric. Econ. Rev. Can. D’Agroecon. 2018, 66, 43–54. [Google Scholar] [CrossRef]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The Molecular Genetics of Crop Domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [Green Version]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
- McClung, C.R. Circadian Clock Components Offer Targets for Crop Domestication and Improvement. Genes 2021, 12, 374. [Google Scholar] [CrossRef] [PubMed]
- Koytsoumpa, E.I.; Bergins, C.; Buddenberg, T.; Wu, S.; Sigurbjörnsson, Ó.; Tran, K.C.; Kakaras, E. The Challenge of Energy Storage in Europe: Focus on Power to Fuel. J. Energy Resour. Technol. 2016, 138, 042002. [Google Scholar] [CrossRef]
- Cohen, A.R.; Chen, G.; Berger, E.M.; Warrier, S.; Lan, G.; Grubert, E.; Dellaert, F.; Chen, Y. Dynamically Controlled Environment Agriculture: Integrating Machine Learning and Mechanistic and Physiological Models for Sustainable Food Cultivation. ACS ES&T Eng. 2022, 2, 3–19. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Carbonnel, M.; Stormonth-Darling, J.M.; Liu, W.; Kuziak, D.; Jones, M.A. Realising the Environmental Potential of Vertical Farming Systems through Advances in Plant Photobiology. Biology 2022, 11, 922. https://doi.org/10.3390/biology11060922
de Carbonnel M, Stormonth-Darling JM, Liu W, Kuziak D, Jones MA. Realising the Environmental Potential of Vertical Farming Systems through Advances in Plant Photobiology. Biology. 2022; 11(6):922. https://doi.org/10.3390/biology11060922
Chicago/Turabian Stylede Carbonnel, Matthieu, John M. Stormonth-Darling, Weiqi Liu, Dmytro Kuziak, and Matthew Alan Jones. 2022. "Realising the Environmental Potential of Vertical Farming Systems through Advances in Plant Photobiology" Biology 11, no. 6: 922. https://doi.org/10.3390/biology11060922
APA Stylede Carbonnel, M., Stormonth-Darling, J. M., Liu, W., Kuziak, D., & Jones, M. A. (2022). Realising the Environmental Potential of Vertical Farming Systems through Advances in Plant Photobiology. Biology, 11(6), 922. https://doi.org/10.3390/biology11060922