
l-Serine Biosensor-Controlled Fermentative Production of l-Tryptophan Derivatives by Corynebacterium glutamicum




Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
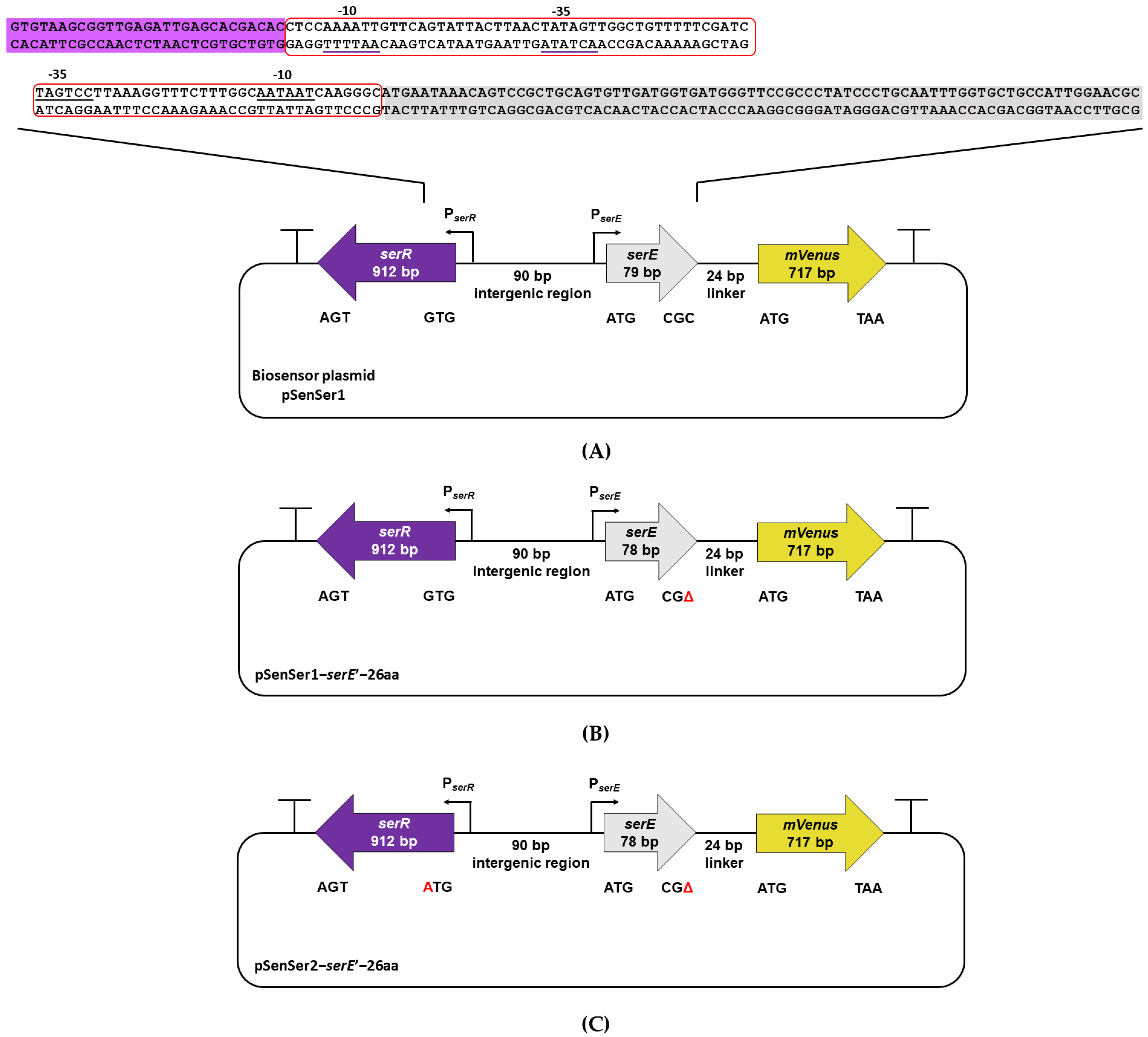
2.2. Molecular Genetic Techniques, Plasmids, and Strain Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmids | Description | Source |
|---|---|---|
| pGP2 | pVWEx1 [51]-derived E. coli-C. glutamicum expression vector without lacIq and tac promoter; KmR, pHM1519 oriVCg | [49] |
| pGP21 | pGP2-derived E. coli-C. glutamicum shuttle vector | This work |
| pSenSer1 | pGP21 containing serR, intergenic region of serR and serE, and transcriptional fusion of serE with mVenus | This work |
| pSenSer1-serE’-26 aa | pSenSer1 with single-point mutation in the serE-mVenus transcriptional fusion to yield a 78-bp serE | This work |
| pSenSer2-serE’-26 aa | pSenSer1-serE’-26 aa with start codon exchange of serR from GTG to ATG | This work |
| pSdS-trpB | pSenSer1 with mVenus replaced by trpB | This work |
| pK19mobsacB | KmR, oriVEc, sacB, lacZα; E. coli-C. glutamicum shuttle vector for construction of insertion and deletion mutants in C. glutamicum | [47] |
| pK19mobsacB ΔserA | pK19mobsacB with a construct for in-frame deletion of serA | This work |
| pK19mobsacB ΔtrpBA | pK19mobsacB with a construct for in-frame deletion of trpBA | This work |
| pEKEx3 | SpecR, Ptac lacIq, pBL1 oriVCg, sacB, lacZα; E. coli-C. glutamicum expression vector | [52] |
| pEKEx3-trpBA | pEKEx3 overexpressing trpBA | This work |
| pEKEx3-trpB | pEKEx3 overexpressing trpB | This work |
2.3. Quantification of l-Serine and Aromatic Compounds
3. Results
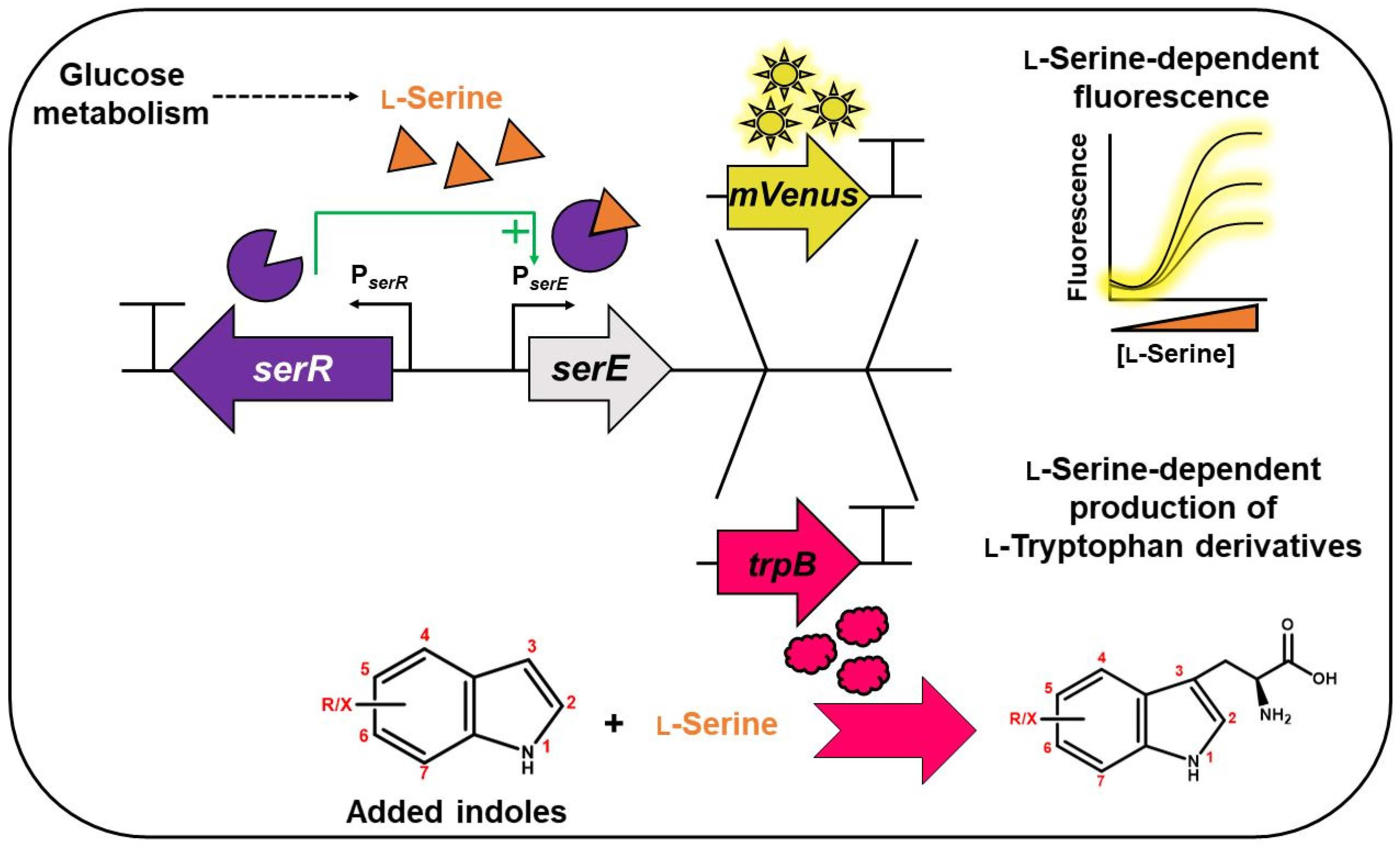
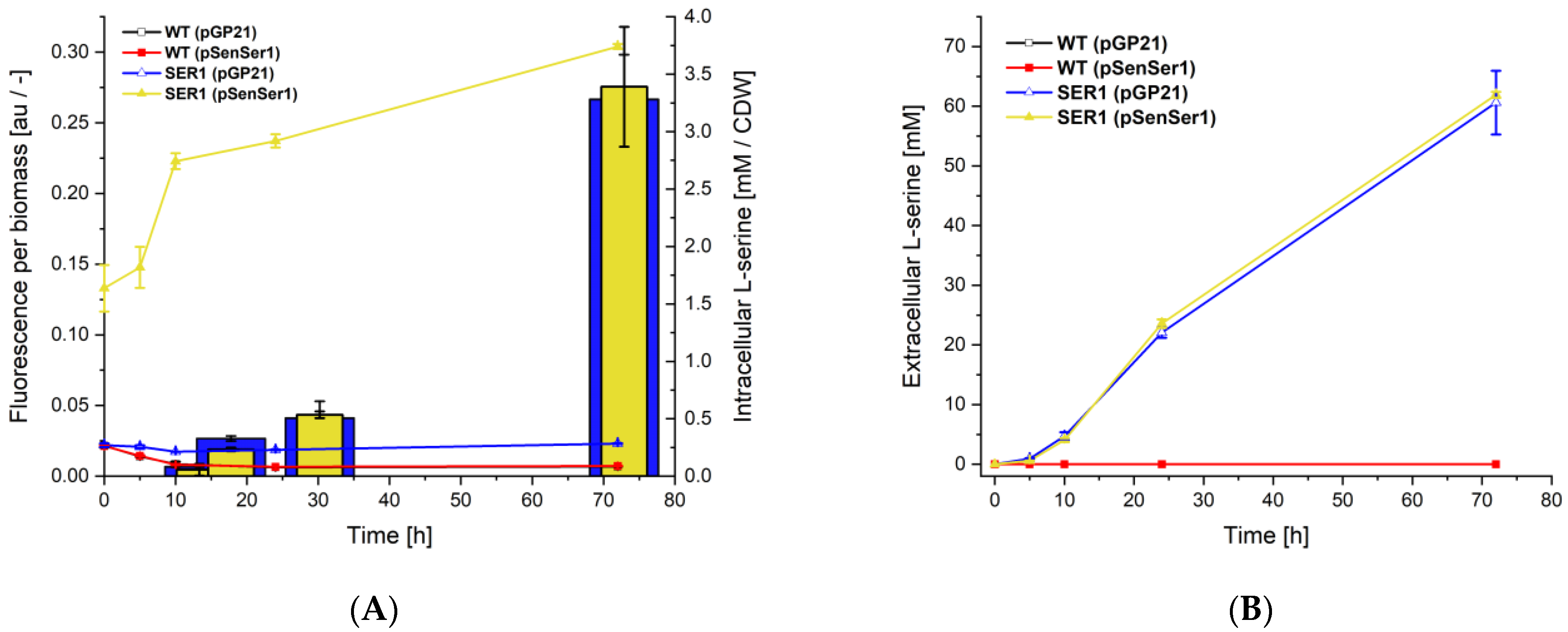
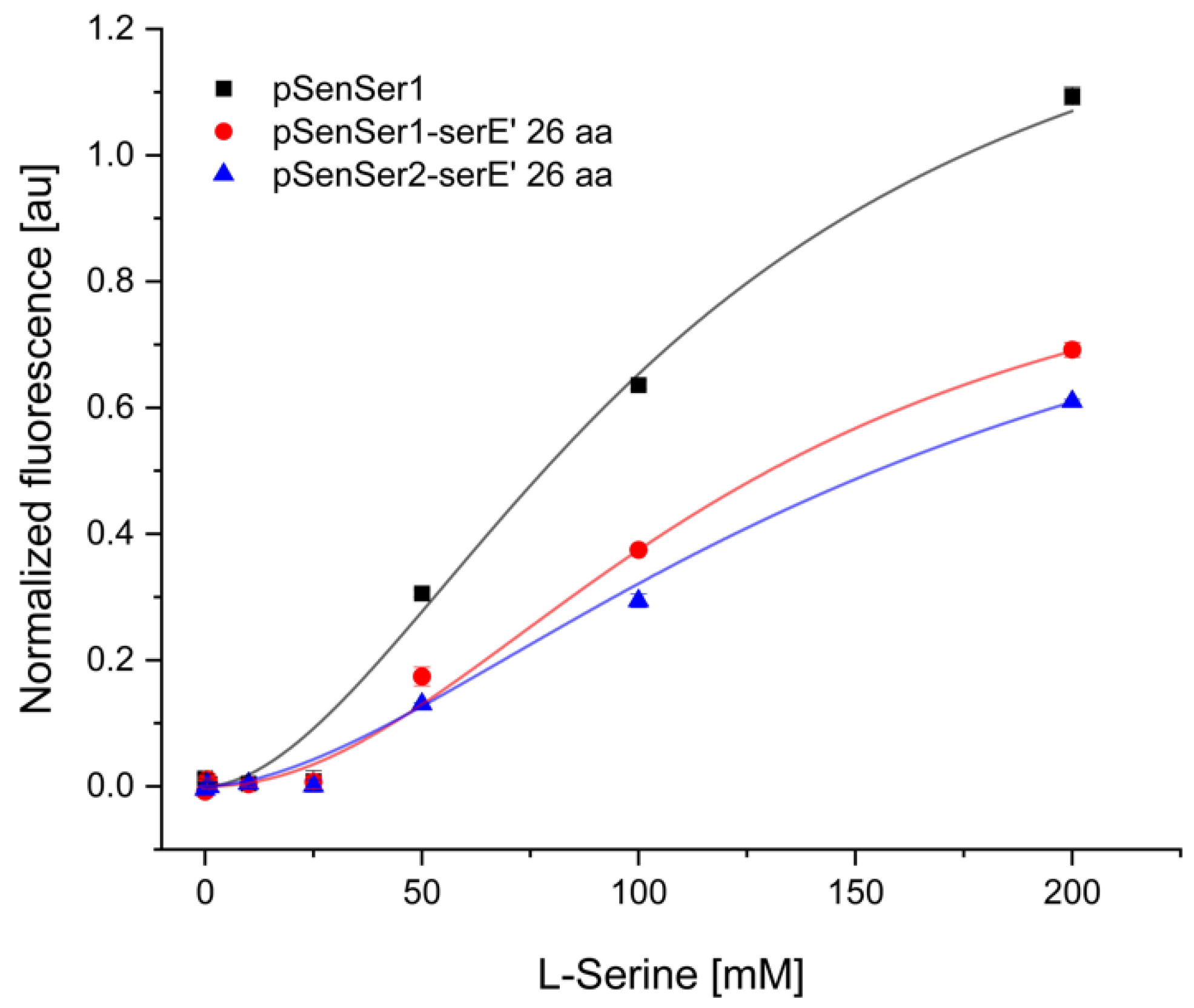
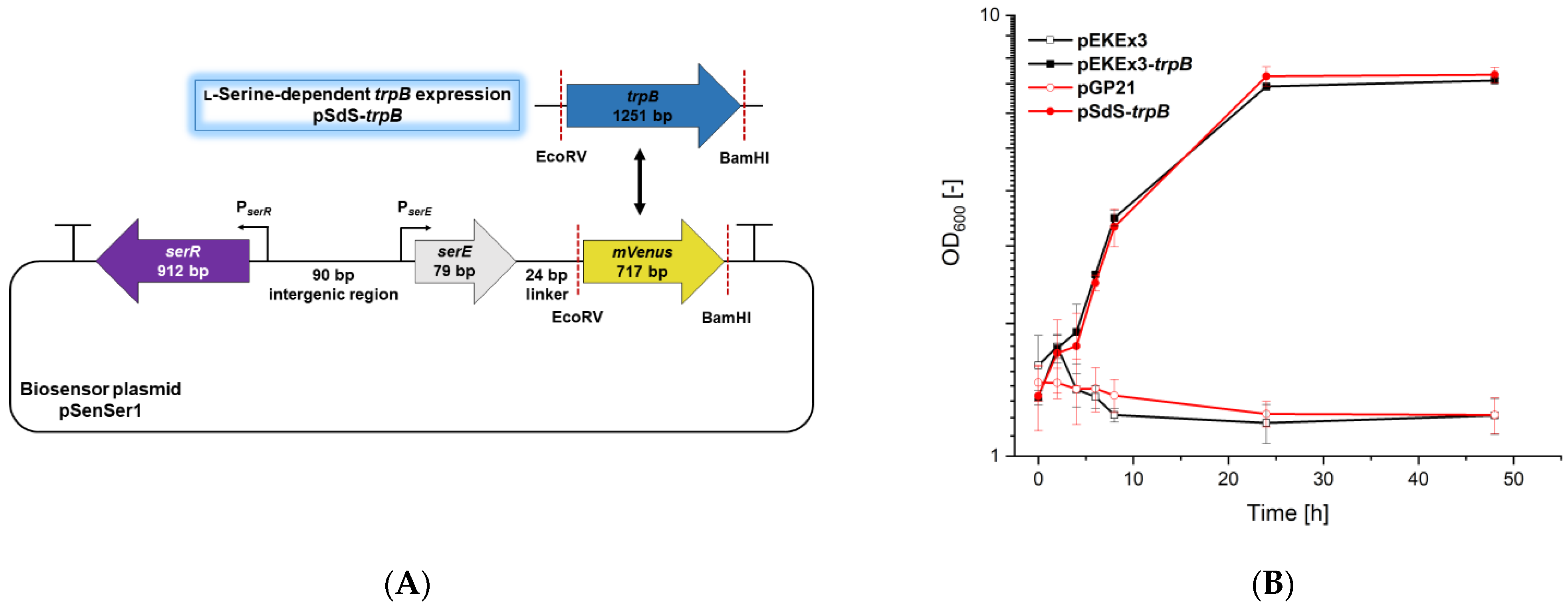
3.1. Design and Construction of an l-Serine-Responsive Biosensor
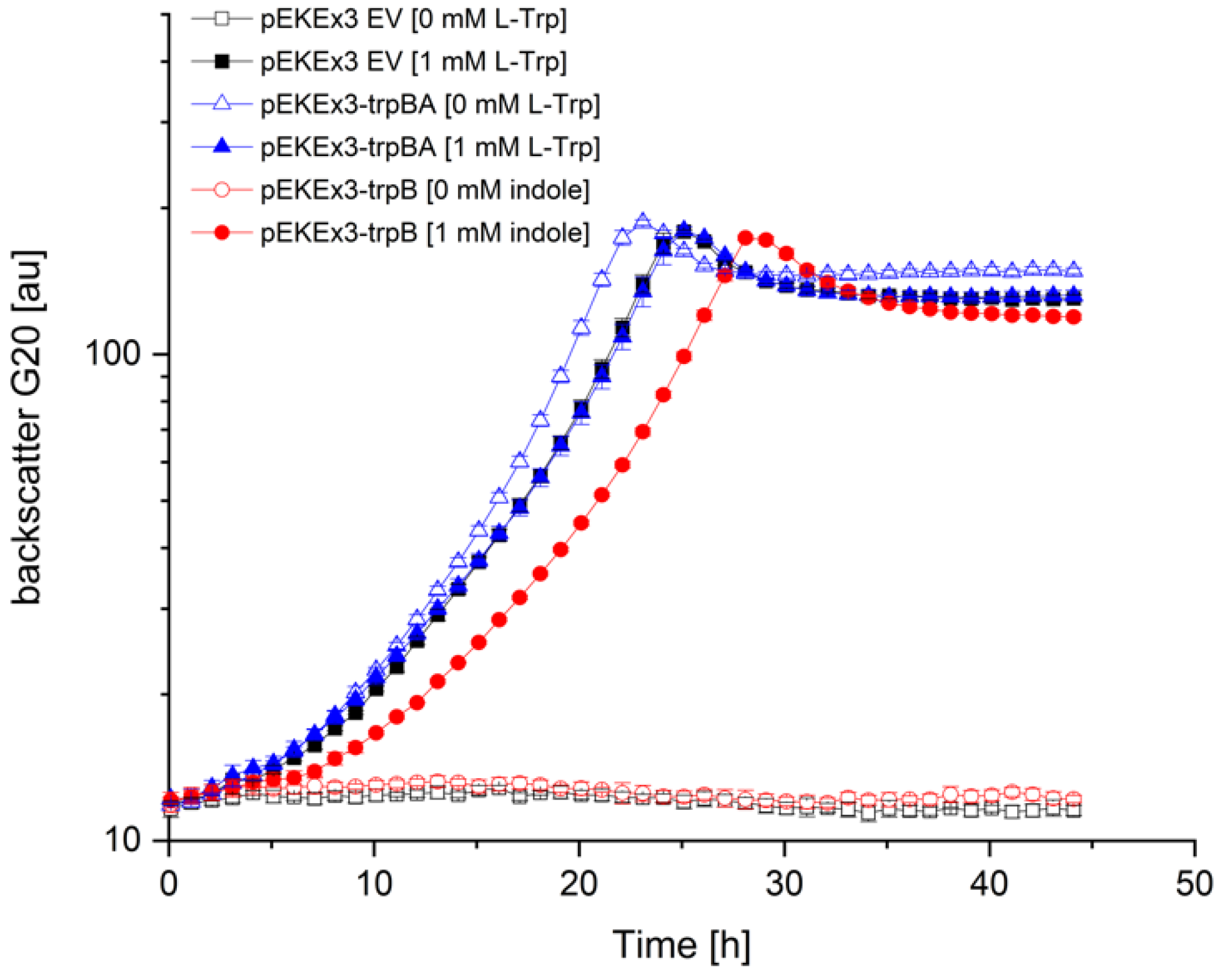
3.2. Construction of an Indole-Essential C. glutamicum Strain
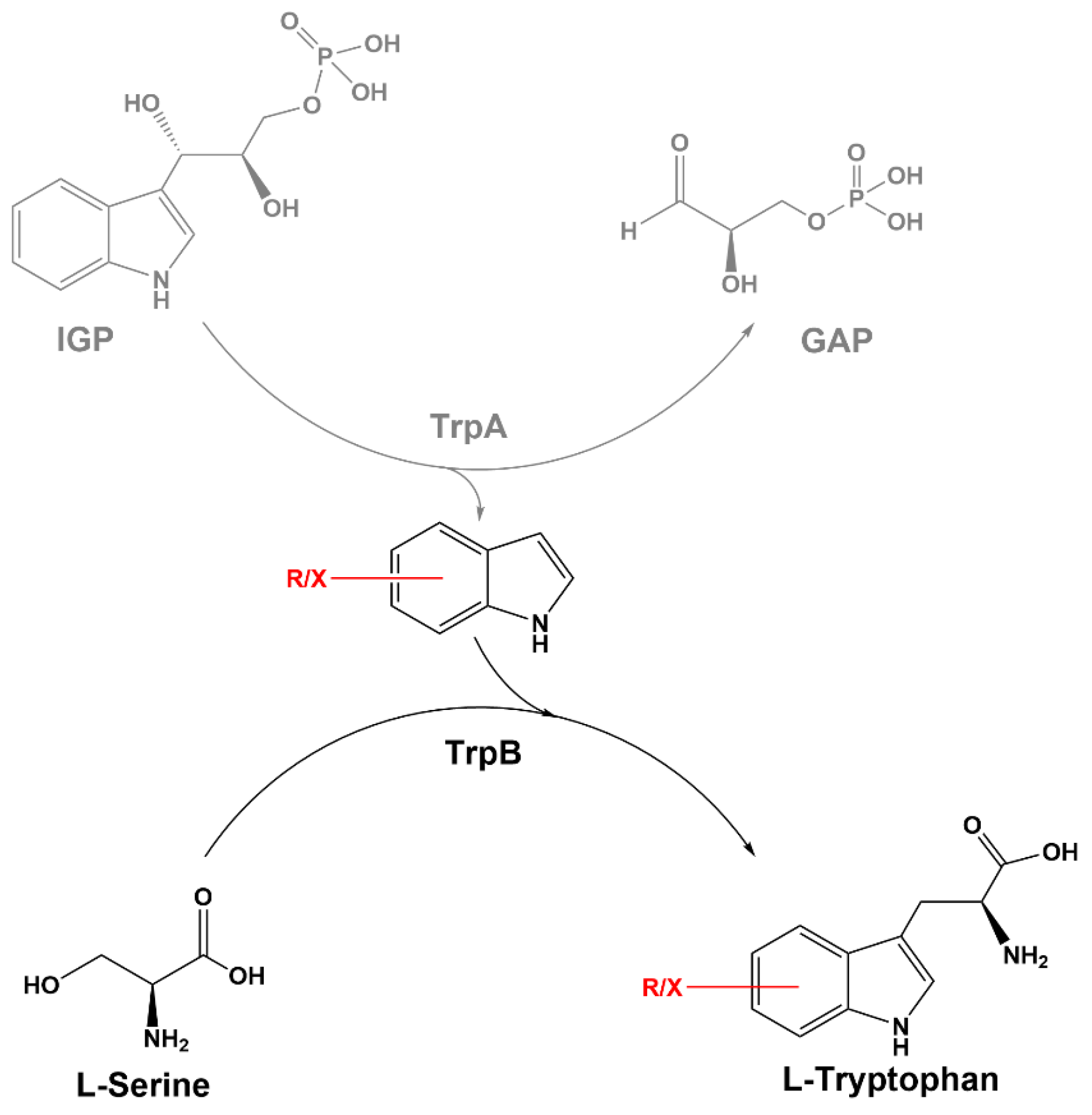
3.3. On-Demand Conversion of Indole and l-Serine to l-Tryptophan via l-Serine-Responsive Biosensor
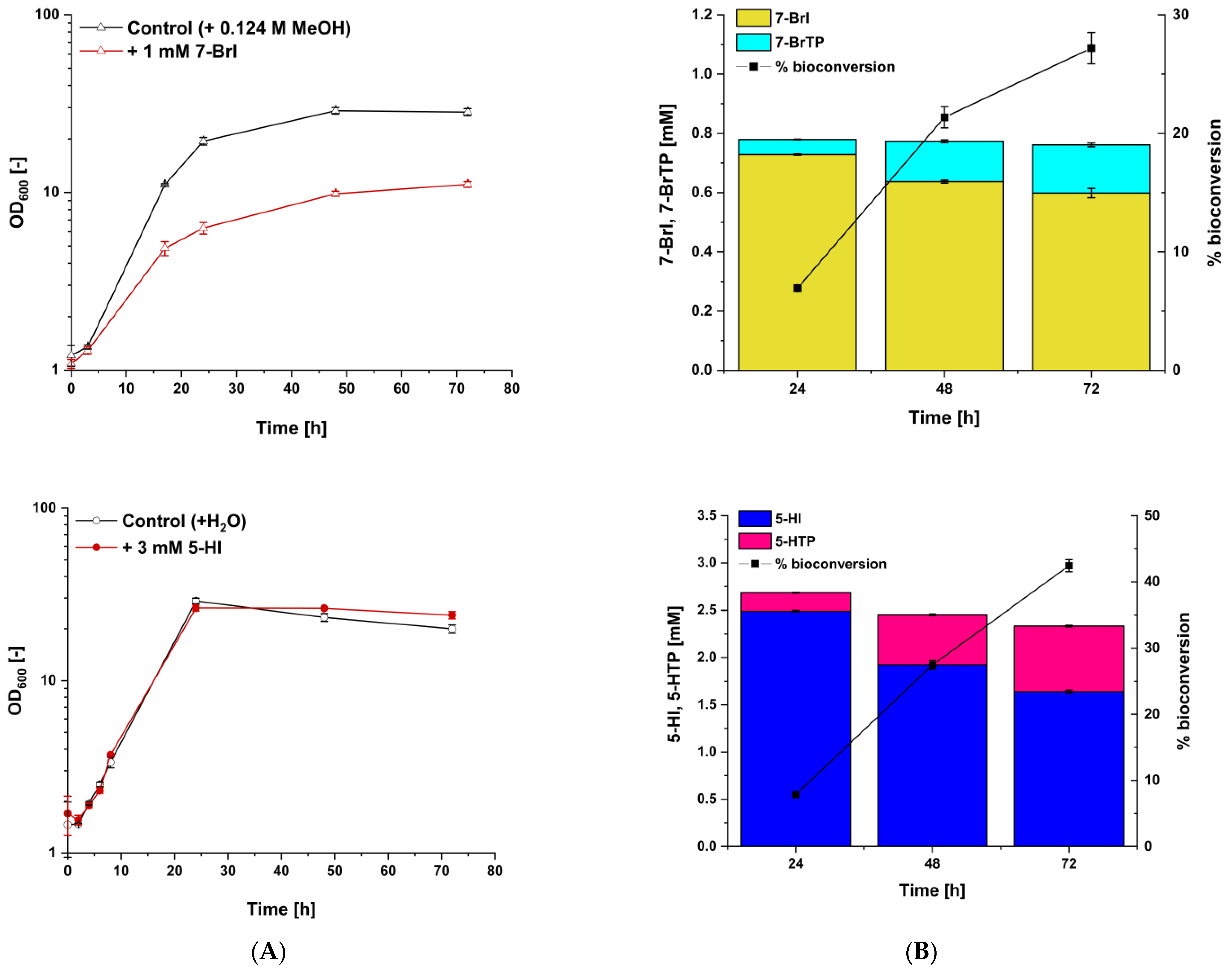
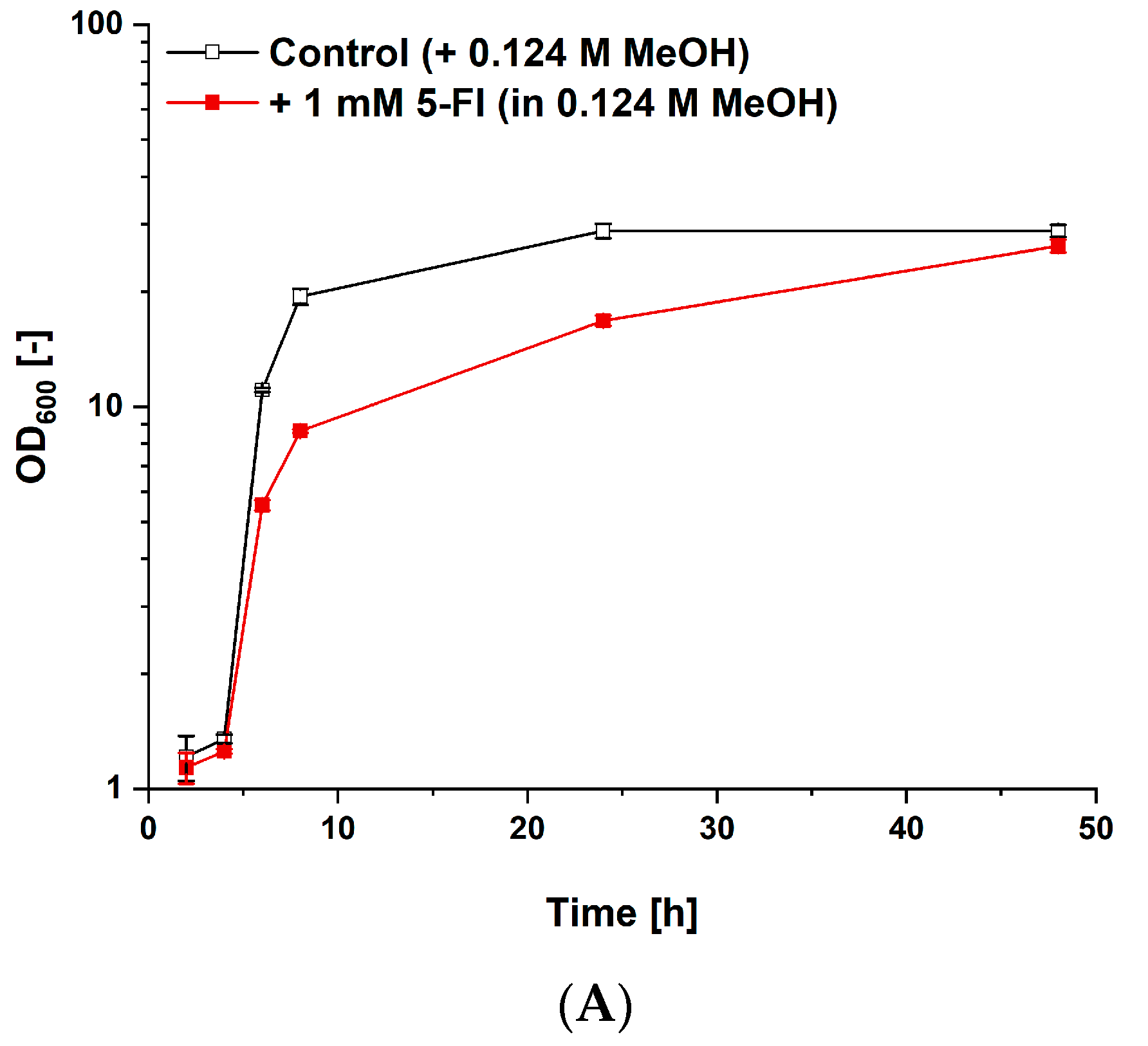
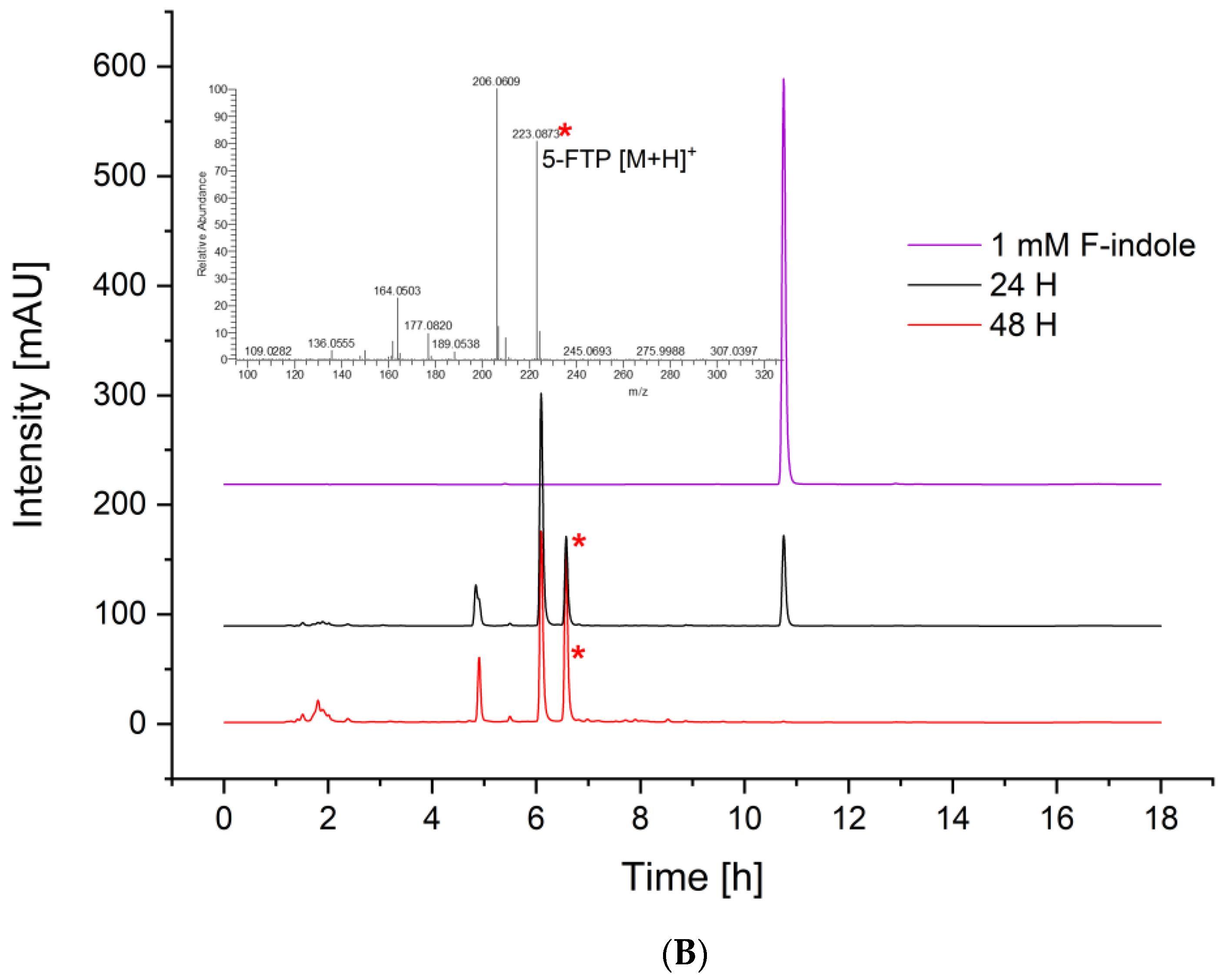
3.4. Fermentative Production of l-Tryptophan Derivatives by C. glutamicum SER2 (pSdS-trpB) from Indole Derivatives
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mindt, M.; Walter, T.; Kugler, P.; Wendisch, V.F. Microbial engineering for production of N-functionalized amino acids and amines. Biotechnol. J. 2020, 15, e1900451. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Granold, M.; Hajieva, P.; Toşa, M.I.; Irimie, F.-D.; Moosmann, B. Modern diversification of the amino acid repertoire driven by oxygen. Proc. Natl. Acad. Sci. USA 2017, 115, 41–46. [Google Scholar] [CrossRef]
- Walsh, C.T. Biological matching of chemical reactivity: Pairing indole nucleophilicity with electrophilic isoprenoids. ACS Chem. Biol. 2014, 9, 2718–2728. [Google Scholar] [CrossRef]
- Mindt, M.; Kashkooli, A.B.; Suarez-Diez, M.; Ferrer, L.; Jilg, T.; Bosch, D.; Martins Dos Santos, V.; Wendisch, V.F.; Cankar, K. Production of indole by Corynebacterium glutamicum microbial cell factories for flavor and fragrance applications. Microb. Cell Fact. 2022, 21, 45. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, L.; Mindt, M.; Suarez-Diez, M.; Jilg, T.; Zagorščak, M.; Lee, J.-H.; Gruden, K.; Wendisch, V.F.; Cankar, K. Fermentative indole production via bacterial tryptophan synthase alpha subunit and plant indole-3-glycerol phosphate lyase nzymes. J. Agric. Food Chem. 2022, 70, 5634–5645. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.-H.; Jung, S.; Lee, S.; Cheon, C.-I.; Song, E.; Lee, S.-S.; Shin, J.; Lee, M.-S. Purification and characterization of an arginine regulatory protein, ArgR, in Corynebacterium glutamicum. J. Ind. Microbiol. Biotechnol. 2011, 38, 1911–1920. [Google Scholar] [CrossRef]
- Bellmann, A.; Vrljić, M.; Pátek, M.; Sahm, H.; Krämer, R.; Eggeling, L. Expression control and specificity of the basic amino acid exporter LysE of Corynebacterium glutamicum. Microbiology 2001, 147, 1765–1774. [Google Scholar] [CrossRef]
- Kubota, T.; Tanaka, Y.; Takemoto, N.; Hiraga, K.; Yukawa, H.; Inui, M. Identification and expression analysis of a gene encoding a shikimate transporter of Corynebacterium glutamicum. Microbiology 2015, 161, 254–263. [Google Scholar] [CrossRef]
- Schweitzer, J.E.; Stolz, M.; Diesveld, R.; Etterich, H.; Eggeling, L. The serine hydroxymethyltransferase gene glyA in Corynebacterium glutamicum is controlled by GlyR. J. Biotechnol. 2009, 139, 214–221. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Xu, G.; Zhang, X.; Shi, J.; Xu, Z. Integration of ARTP mutagenesis with biosensor-mediated high-throughput screening to improve L-serine yield in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2018, 102, 5939–5951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, Y.; Chen, Z.; Xu, G.; Zhang, X.; Li, H.; Shi, J.; Koffas, M.A.G.; Xu, Z. High-yield production of L-serine through a novel identified exporter combined with synthetic pathway in Corynebacterium glutamicum. Microb. Cell Fact. 2020, 19, 115. [Google Scholar] [CrossRef] [PubMed]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; National Renewable Energy Lab.: Golden, CO, USA, 2004. [Google Scholar]
- Lai, S.; Zhang, Y.; Liu, S.; Liang, Y.; Shang, X.; Chai, X.; Wen, T. Metabolic engineering and flux analysis of Corynebacterium glutamicum for L-serine production. Sci. China Life Sci. 2012, 55, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Simic, P.; Willuhn, J.; Sahm, H.; Eggeling, L. Identification of glyA (encoding serine hydroxymethyltransferase) and its use together with the exporter ThrE To increase L-threonine accumulation by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2002, 68, 3321–3327. [Google Scholar] [CrossRef] [PubMed]
- Stolz, M.; Peters-Wendisch, P.; Etterich, H.; Gerharz, T.; Faurie, R.; Sahm, H.; Fersterra, H.; Eggeling, L. Reduced folate supply as a key to enhanced L-serine production by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2007, 73, 750–755. [Google Scholar] [CrossRef]
- Mustafi, N.; Grünberger, A.; Kohlheyer, D.; Bott, M.; Frunzke, J. The development and application of a single-cell biosensor for the detection of L-methionine and branched-chain amino acids. Metab. Eng. 2012, 14, 449–457. [Google Scholar] [CrossRef]
- Mahr, R.; Gätgens, C.; Gätgens, J.; Polen, T.; Kalinowski, J.; Frunzke, J. Biosensor-driven adaptive laboratory evolution of L-valine production in Corynebacterium glutamicum. Metab. Eng. 2015, 32, 184–194. [Google Scholar] [CrossRef]
- Lange, C.; Mustafi, N.; Frunzke, J.; Kennerknecht, N.; Wessel, M.; Bott, M.; Wendisch, V.F. Lrp of Corynebacterium glutamicum controls expression of the brnFE operon encoding the export system for L-methionine and branched-chain amino acids. J. Biotechnol. 2012, 158, 231–241. [Google Scholar] [CrossRef]
- Binder, S.; Siedler, S.; Marienhagen, J.; Bott, M.; Eggeling, L. Recombineering in Corynebacterium glutamicum combined with optical nanosensors: A general strategy for fast producer strain generation. Nucleic Acids Res. 2013, 41, 6360–6369. [Google Scholar] [CrossRef]
- Kortmann, M.; Mack, C.; Baumgart, M.; Bott, M. Pyruvate carboxylase variants enabling improved lysine production from glucose identified by biosensor-based high-throughput fluorescence-activated cell sorting screening. ACS Synth. Biol. 2019, 8, 274–281. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, B.; Liu, Y.-M.; Yang, K.-Q.; Liu, S.-J. New intracellular shikimic acid biosensor for monitoring shikimate synthesis in Corynebacterium glutamicum. ACS Synth. Biol. 2018, 7, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Schulte, J.; Baumgart, M.; Bott, M. Development of a single-cell GlxR-based cAMP biosensor for Corynebacterium glutamicum. J. Biotechnol. 2017, 258, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Binder, S.; Schendzielorz, G.; Stäbler, N.; Krumbach, K.; Hoffmann, K.; Bott, M.; Eggeling, L. A high-throughput approach to identify genomic variants of bacterial metabolite producers at the single-cell level. Genome Biol. 2012, 13, R40. [Google Scholar] [CrossRef] [PubMed]
- Brune, I.; Jochmann, N.; Brinkrolf, K.; Hüser, A.T.; Gerstmeir, R.; Eikmanns, B.J.; Kalinowski, J.; Pühler, A.; Tauch, A. The IclR-type transcriptional repressor LtbR regulates the expression of leucine and tryptophan biosynthesis genes in the amino acid producer Corynebacterium glutamicum. J. Bacteriol. 2007, 189, 2720–2733. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Matsui, K. Structure and function of the trp operon control regions of Brevibacterium lactofermentum, a glutamic-acid-producing bacterium. Gene 1987, 53, 191–200. [Google Scholar] [CrossRef]
- Matsui, K.; Sano, K.; Ohtsubo, E. Sequence analysis of the Brevibacterium lactofermentum trp operon. Mol. Gen. Genet. 1987, 209, 299–305. [Google Scholar] [CrossRef]
- Purwanto, H.S.; Kang, M.-S.; Ferrer, L.; Han, S.S.; Lee, J.Y.; Kim, H.S.; Lee, J.H. Rational engineering of the shikimate and related pathways in Corynebacterium glutamicum for 4-hydroxybenzoate production. J. Biotechnol. 2018, 282, 92–100. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.; Kim, B.; Ferrer, L.; Choi, J.; Wendisch, V.F.; Lee, J. Engineered Corynebacterium glutamicum as platform for the production of aromatic aldehydes. Front. Bioeng. Biotechnol. 2022. [Google Scholar] [CrossRef]
- Ikeda, M. Towards bacterial strains overproducing L-tryptophan and other aromatics by metabolic engineering. Appl. Microbiol. Biotechnol. 2006, 69, 615–626. [Google Scholar] [CrossRef]
- Ikeda, M.; Katsumata, R. Hyperproduction of tryptophan by Corynebacterium glutamicum with the modified pentose phosphate pathway. Appl. Environ. Microbiol. 1999, 65, 2497–2502. [Google Scholar] [CrossRef]
- Katsumata, R.; Ikeda, M. Hyperproduction of tryptophan in Corynebacterium glutamicum by pathway engineering. Nat. Biotechnol. 1993, 11, 921–925. [Google Scholar] [CrossRef]
- Phillips, R.S. Synthetic applications of tryptophan synthase. Tetrahedron Asymmetry 2004, 15, 2787–2792. [Google Scholar] [CrossRef]
- Goss, R.J.M.; Newill, P.L.A. A convenient enzymatic synthesis of L-halotryptophans. Chem. Commun. 2006, 47, 4924–4925. [Google Scholar] [CrossRef] [PubMed]
- Murciano-Calles, J.; Romney, D.K.; Brinkmann-Chen, S.; Buller, A.R.; Arnold, F.H. A panel of TrpB biocatalysts derived from tryptophan synthase through the transfer of mutations that mimic allosteric activation. Angew. Chem.-Int. Ed. 2016, 55, 11577–11581. [Google Scholar] [CrossRef]
- Inman, M.; Moody, C. Indole synthesis- something old, something new. Chem. Sci. 2013, 4, 29–41. [Google Scholar] [CrossRef]
- Vaswani, R.G.; Albrecht, B.K.; Audia, J.E.; Côte, A.; Dakin, L.A.; Duplessis, M.; Gehling, V.S.; Harmange, J.-C.; Hewitt, M.C.; Leblanc, Y.; et al. A practical synthesis of indoles via a Pd-Catalyzed C−N ring formation. Org. Lett. 2014, 16, 4114–4117. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Manabe, K. One-pot synthesis of 2,4-disubstituted indoles from N-Tosyl-2,3-dichloroaniline using palladium-dihydroxyterphenylphosphine. Org. Lett. 2014, 16, 2386–2389. [Google Scholar] [CrossRef]
- Peters-Wendisch, P.; Stolz, M.; Etterich, H.; Kennerknecht, N.; Sahm, H.; Eggeling, L. Metabolic engineering of Corynebacterium glutamicum for L-serine production. Appl. Environ. Microbiol. 2005, 71, 7139–7144. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on Transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Eggeling, L.; Bott, M. Handbook of Corynebacterium Glutamicum; CRC Press, Taylor Francis Group: Boca Raton, FL, USA, 2005; ISBN 978-1-4200-3969-6. [Google Scholar]
- Funke, M.; Diederichs, S.; Kensy, F.; Müller, C.; Büchs, J. The baffled microtiter plate: Increased oxygen transfer and improved online monitoring in small scale fermentations. Biotechnol. Bioeng. 2009, 103, 1118–1128. [Google Scholar] [CrossRef]
- Kensy, F.; Zang, E.; Faulhammer, C.; Tan, R.-K.; Büchs, J. Validation of a high-throughput fermentation system based on online monitoring of biomass and fluorescence in continuously shaken microtiter plates. Microb. Cell Fact. 2009, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization sytem for in vivo genetic engineering: Transposon mutagenesis in Gram negative bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Baumgart, M.; Unthan, S.; Kloß, R.; Radek, A.; Polen, T.; Tenhaef, N.; Müller, M.F.; Küberl, A.; Siebert, D.; Brühl, N.; et al. Corynebacterium glutamicum chassis C1∗: Building and testing a novel platform host for synthetic biology and industrial biotechnology. ACS Synth. Biol. 2018, 7, 132–144. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbachb, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Craig Venter, J.; Hutchison III, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Kugler, P.; Fröhlich, D.; Wendisch, V.F. Development of a biosensor for crotonobetaine-CoA ligase screening based on the elucidation of Escherichia coli carnitine metabolism. ACS Synth. Biol. 2020, 9, 2460–2471. [Google Scholar] [CrossRef]
- Gauttam, R.; Desiderato, C.; Jung, L.; Shah, A.; Eikmanns, B.J. A step forward: Compatible and dual-inducible expression vectors for gene co-expression in Corynebacterium glutamicum. Plasmid 2019, 101, 20–27. [Google Scholar] [CrossRef]
- Peters-Wendisch, P.; Kreutzer, C.; Kalinowski, J.; Pátek, M.; Sahm, H.; Eikmanns, B.J. Pyruvate carboxylase from Corynebacterium glutamicum: Characterization, expression and inactivation of the pyc gene. Microbiology 1998, 144, 915–927. [Google Scholar] [CrossRef]
- Stansen, C.; Uy, D.; Delaunay, S.; Eggeling, L.; Goergen, J.-L.; Wendisch, V.F. Characterization of a Corynebacterium glutamicum lactate utilization operon induced during temperature-triggered glutamate production. Appl. Environ. Microbiol. 2005, 71, 5920–5928. [Google Scholar] [CrossRef]
- Liu, H.; Naismith, J.H. An efficient one-step site-directed deletion, insertion, single and multiple-site plasmid mutagenesis protocol. BMC Biotechnol. 2008, 8, 91. [Google Scholar] [CrossRef]
- Walter, T.; Al Medani, N.; Burgardt, A.; Cankar, K.; Ferrer, L.; Kerbs, A.; Lee, J.-H.; Mindt, M.; Risse, J.M.; Wendisch, V.F. Fermentative N-methylanthranilate production by engineered Corynebacterium glutamicum. Microorganisms 2020, 8, 866. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Guo, W.; Wang, F.; Peng, F.; Yang, Y.; Dai, X.; Liu, X.; Bai, Z. Transcriptome and multivariable data analysis of Corynebacterium glutamicum under different dissolved oxygen conditions in bioreactors. PLoS ONE 2016, 11, e0167156. [Google Scholar] [CrossRef] [PubMed]
- Bolten, C.J.; Kiefer, P.; Letisse, F.; Portais, J.-C.; Wittmann, C. Sampling for metabolome analysis of microorganisms. Anal. Chem. 2007, 79, 3843–3849. [Google Scholar] [CrossRef] [PubMed]
- Kremers, G.-J.; Goedhart, J.; van Munster, E.B.; Gadella, T.W.J. Cyan and yellow super fluorescent proteins with improved brightness, protein folding, and FRET Förster radius. Biochemistry 2006, 45, 6570–6580. [Google Scholar] [CrossRef]
- Netzer, R.; Peters-Wendisch, P.; Eggeling, L.; Sahm, H. Cometabolism of a nongrowth substrate: L-serine utilization by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2004, 70, 7148–7155. [Google Scholar] [CrossRef]
- Niks, D.; Hilario, E.; Dierkers, A.; Ngo, H.; Borchardt, D.; Neubauer, T.J.; Fan, L.; Mueller, L.J.; Dunn, M.F. Allostery and substrate channeling in the tryptophan synthase bienzyme complex: Evidence for two subunit conformations and four quaternary states. Biochemistry 2013, 52, 6396–6411. [Google Scholar] [CrossRef][Green Version]
- Watkins-Dulaney, E.; Straathof, S.; Arnold, F. Tryptophan synthase: Biocatalyst extraordinaire. ChemBioChem 2021, 22, 5–16. [Google Scholar] [CrossRef]
- Veldmann, K.H.; Dachwitz, S.; Risse, J.M.; Lee, J.-H.; Sewald, N.; Wendisch, V.F. Bromination of L-tryptophan in a fermentative process with Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2019, 7, 219. [Google Scholar] [CrossRef]
- Alkhalaf, L.M.; Ryan, K.S. Biosynthetic manipulation of tryptophan in bacteria: Pathways and mechanisms. Chem. Biol. 2015, 22, 317–328. [Google Scholar] [CrossRef]
- Buller, A.R.; Brinkmann-Chen, S.; Romney, D.K.; Herger, M.; Murciano-Calles, J.; Arnold, F.H. Directed evolution of the tryptophan synthase β-subunit for stand-alone function recapitulates allosteric activation. Proc. Natl. Acad. Sci. USA 2015, 112, 14599–14604. [Google Scholar] [CrossRef]
- Buller, A.R.; Van Roye, P.; Cahn, J.K.B.; Scheele, R.A.; Herger, M.; Arnold, F.H. Directed evolution mimics allosteric activation by stepwise tuning of the conformational ensemble. J. Am. Chem. Soc. 2018, 140, 7256–7266. [Google Scholar] [CrossRef] [PubMed]
- Salwiczek, M.; Nyakatura, E.K.; Gerling, U.I.M.; Ye, S.; Koksch, B. Fluorinated amino acids: Compatibility with native protein structures and effects on protein-protein interactions. Chem. Soc. Rev. 2012, 41, 2135–2171. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Huang, F.; Deng, H.; Schaffrath, C.; Spencer, J.B.; O’Hagan, D.; Naismith, J.H. Crystal structure and mechanism of a bacterial fluorinating enzyme. Nature 2004, 427, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Miles, Z.D.; Winter, J.M.; Eustáquio, A.S.; El Gamal, A.A.; Moore, B.S. Enzymatic halogenation and dehalogenation reactions: Pervasive and mechanistically diverse. Chem. Rev. 2017, 117, 5619–5674. [Google Scholar] [CrossRef] [PubMed]
- Koguchi, Y.; Kohno, J.; Nishio, M.; Takahashi, K.; Okuda, T.; Ohnuki, T.; Komatsubara, S. TMC-95A, B, C, and D, novel proteasome inhibitors produced by Apiospora montagnei Sacc. TC 1093. Taxonomy, production, isolation, and biological activities. J. Antibiot. 2000, 53, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Vergnolle, N. Protease inhibition as new therapeutic strategy for GI diseases. Gut 2016, 65, 1215–1224. [Google Scholar] [CrossRef]
- Hamdan, F.F.; Ribeiro, P. Characterization of a stable form of tryptophan hydroxylase from the human parasite Schistosoma mansoni. J. Biol. Chem. 1999, 274, 21746–21754. [Google Scholar] [CrossRef]
- Xu, L.; Li, T.; Huo, Z.; Chen, Q.; Xia, Q.; Jiang, B. Directed evolution improves the enzymatic synthesis of L-5-hydroxytryptophan by an engineered tryptophan synthase. Appl. Biochem. Biotechnol. 2021, 193, 3407–3417. [Google Scholar] [CrossRef]
- Kirschner, K.; Wiskocil, R.L.; Foehn, M.; Rezeau, L. The tryptophan synthase from Escherichia coli. An improved purification procedure for the alpha-subunit and binding studies with substrate analogues. Eur. J. Biochem. 1975, 60, 513–523. [Google Scholar] [CrossRef]
- Lane, A.N.; Kirschner, K. The catalytic mechanism of tryptophan synthase from Escherichia coli. Eur. J. Biochem. 1983, 129, 571–582. [Google Scholar] [CrossRef]
- Brown, E.A.; D’Ari, R.; Newman, E.B. A relationship between L-serine degradation and methionine biosynthesis in Escherichia coli K12. J. Gen. Microbiol. 1990, 136, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C.F. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Mannan, A.A.; Liu, D.; Zhang, F.; Oyarzún, D.A. Fundamental design principles for transcription-factor-based metabolite biosensors. ACS Synth. Biol. 2017, 6, 1851–1859. [Google Scholar] [CrossRef] [PubMed]









| Strain | Description | Source |
|---|---|---|
| Escherichia coli | ||
| DH5α | ΔlacU169 (ØdlacZΔM15), supE44, hsdR1d7, recA1, endA1, gyrA96, thi-1, relA1 | [40] |
| S17-1 | recA pro hsdR RP4-2-Tc::Mu-Km::Tn7 | [44] |
| Corynebacterium glutamicum | ||
| ATCC 13032 | C. glutamicum wild-type strain | ATCC |
| ATCC 13032 ΔserA | ATCC 13032 derivative with serA in-frame deletion, l-serine auxotroph | This work |
| C1* | Genome-reduced chassis strain of ATCC 13032 | [45] |
| C1* ΔtrpBA | C1* derivative with trpBA in-frame deletion, l-tryptophan auxotroph | This work |
| SER1 | ATCC13032 derivative with in-frame deletion of sdaA and pabABC and overexpression of serA(fbr), serB, serC, tetracycline-resistant, l-serine overproducer | [16] |
| SER2 | SER1 with in-frame deletion of native trpBA | This work |
| Name | Oligonucleotide Sequence (5′ to 3′) |
|---|---|
| pGP21-F | CAACAGAGTTTGTAGAAACGCAAAAAGGCC |
| pGP21-R | GCTCGGTACCCGGGGATCCTAGCCCGCGGAGTACCGC |
| mVen-F | GGTGCTGCCATTGGAACGCTGAACTTTAAGAAGGAGATATCATATGAGTAAAGGAGAAGAACTTTTCACTGGAGTTGTCC |
| mVen-R | CGCGGTACTCCGCGGGCTAGCGCGGATCCGCGTTATTGTATAGTTCATCCA TGCCATGTGTAATCCCAG |
| serR-F | ATTCGAGCTCGGTACCCGGGTCACTCTACTAGACGAGCCTCCAAATAAGAT CTTTG |
| serE-R | AGTTCTTCTCCTTTACTCATATGATATCTCCTTCTTAAAGTTCAGCGTTCCAA TGGCAGCACCAAATTG |
| SenstrpB-F | CTGAACTTTAAGAAGGAGATATCATATGACTGAAAAAGAAAACTTGGGCG GCTC |
| SenstrpB-R | GTACTCCGCGGGCTAGCGCGGATCCGCGTCATCGGTTGTCCTTCAGGATC AGTTCTG |
| SDM1-F | CTCCTTCTTAAAGTTCACGTTCCAATGGCAGCAC |
| SDM1-R | GATATCTCCTTCTTAAAGTTCACGTTCCAATGGCAGCAC |
| SDM2-F | TGAACAATTTTGGAGGTGTCATGCTCAATCTCAA |
| SDM2-R | TTGAGATTGAGCATGACACCTCCAAAATTGTTCA |
| serAup-F | GCATGCCTGCAGGTCGACTCTAGAGTACGAGACCAGACACACGTGACAAA AAAT |
| serAup-R | GGACTCAACACGCAGGATCAGGACACGTCAACCCAACGGACTTCTACTGC |
| serAdn-F | GCAGTAGAAGTCCGTTGGGTTGACGTGTCCTGATCCTGCGTGTTGAGTC |
| serAdn-R | AGTGAATTCGAGCTCGGTACCCGGGTTAAGCACCGTTGTCAGACCAGTCG |
| serA-conf-F | TAAAATGGCGTTAAATGGCGCGAAATGG |
| serA-conf-R | GGGTAAAGTGCATGAAACTCACGAGTGAG |
| trpBA-F | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGACTGAAAAAGAAAACTTGGGCGGC |
| trpBA-R | AAAACGACGGCCAGTGAATTCTAAACCTTCTTGGTCGCTGCCTTCA |
| trpB-F | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGACTGAAAAAGAAAACTTGGGCGGC |
| trpB-R | AAAACGACGGCCAGTGAATTTCATCGGTTGTCCTTCAGGATCAGTTCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer, L.; Elsaraf, M.; Mindt, M.; Wendisch, V.F. l-Serine Biosensor-Controlled Fermentative Production of l-Tryptophan Derivatives by Corynebacterium glutamicum. Biology 2022, 11, 744. https://doi.org/10.3390/biology11050744
Ferrer L, Elsaraf M, Mindt M, Wendisch VF. l-Serine Biosensor-Controlled Fermentative Production of l-Tryptophan Derivatives by Corynebacterium glutamicum. Biology. 2022; 11(5):744. https://doi.org/10.3390/biology11050744
Chicago/Turabian StyleFerrer, Lenny, Mahmoud Elsaraf, Melanie Mindt, and Volker F. Wendisch. 2022. "l-Serine Biosensor-Controlled Fermentative Production of l-Tryptophan Derivatives by Corynebacterium glutamicum" Biology 11, no. 5: 744. https://doi.org/10.3390/biology11050744
APA StyleFerrer, L., Elsaraf, M., Mindt, M., & Wendisch, V. F. (2022). l-Serine Biosensor-Controlled Fermentative Production of l-Tryptophan Derivatives by Corynebacterium glutamicum. Biology, 11(5), 744. https://doi.org/10.3390/biology11050744