
Hotspot siRNA Confers Plant Resistance against Viral Infection

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Infection and Plant Maintenance
2.2. Detection of ACMV siRNAs
2.3. RNAseq Profiling of ACMC vsiRNAs
2.4. In Vitro Production of siRNA
2.5. DNA Extraction and Southern Blot
3. Results
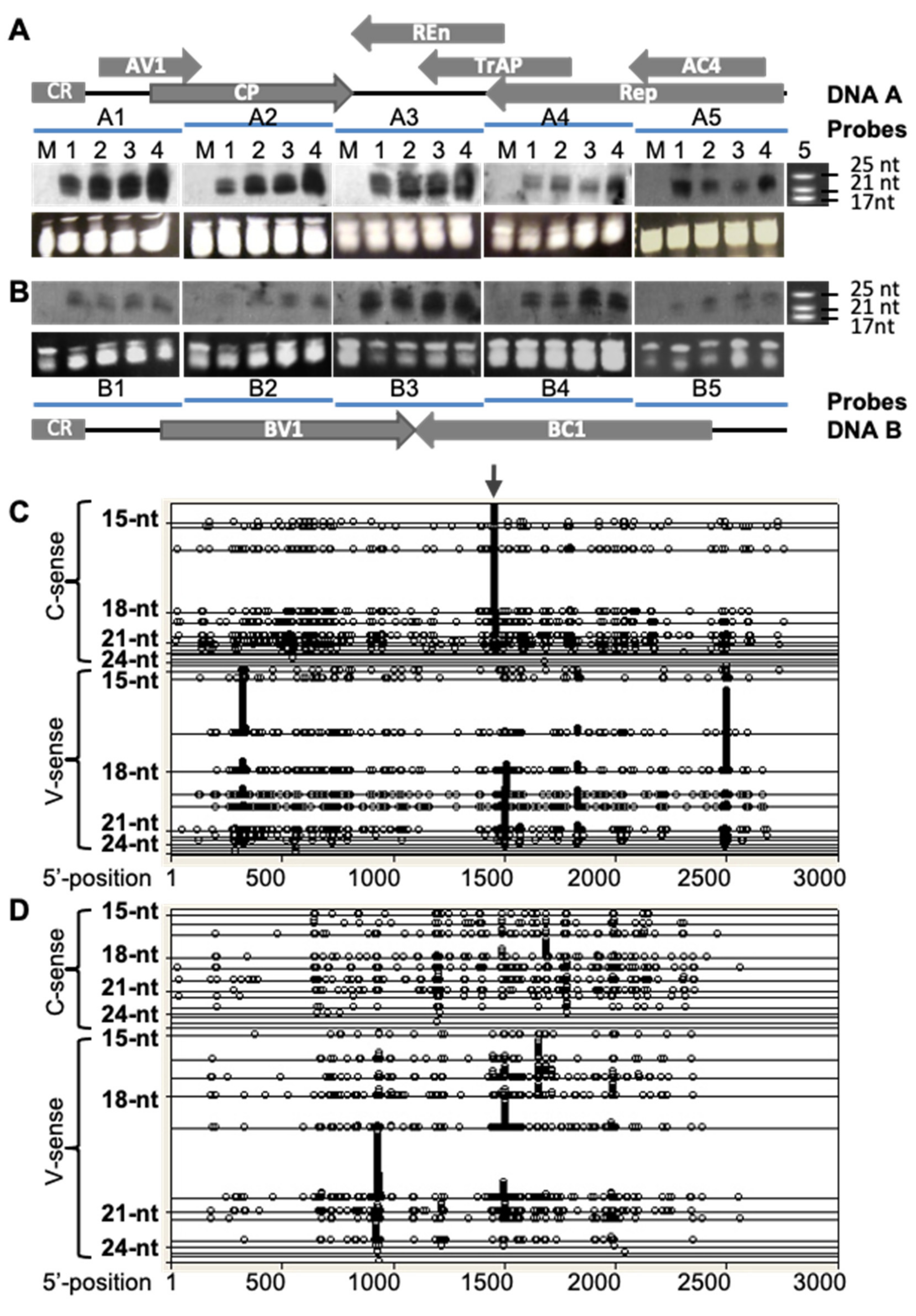
3.1. Profiling ACMV vsiRNAs
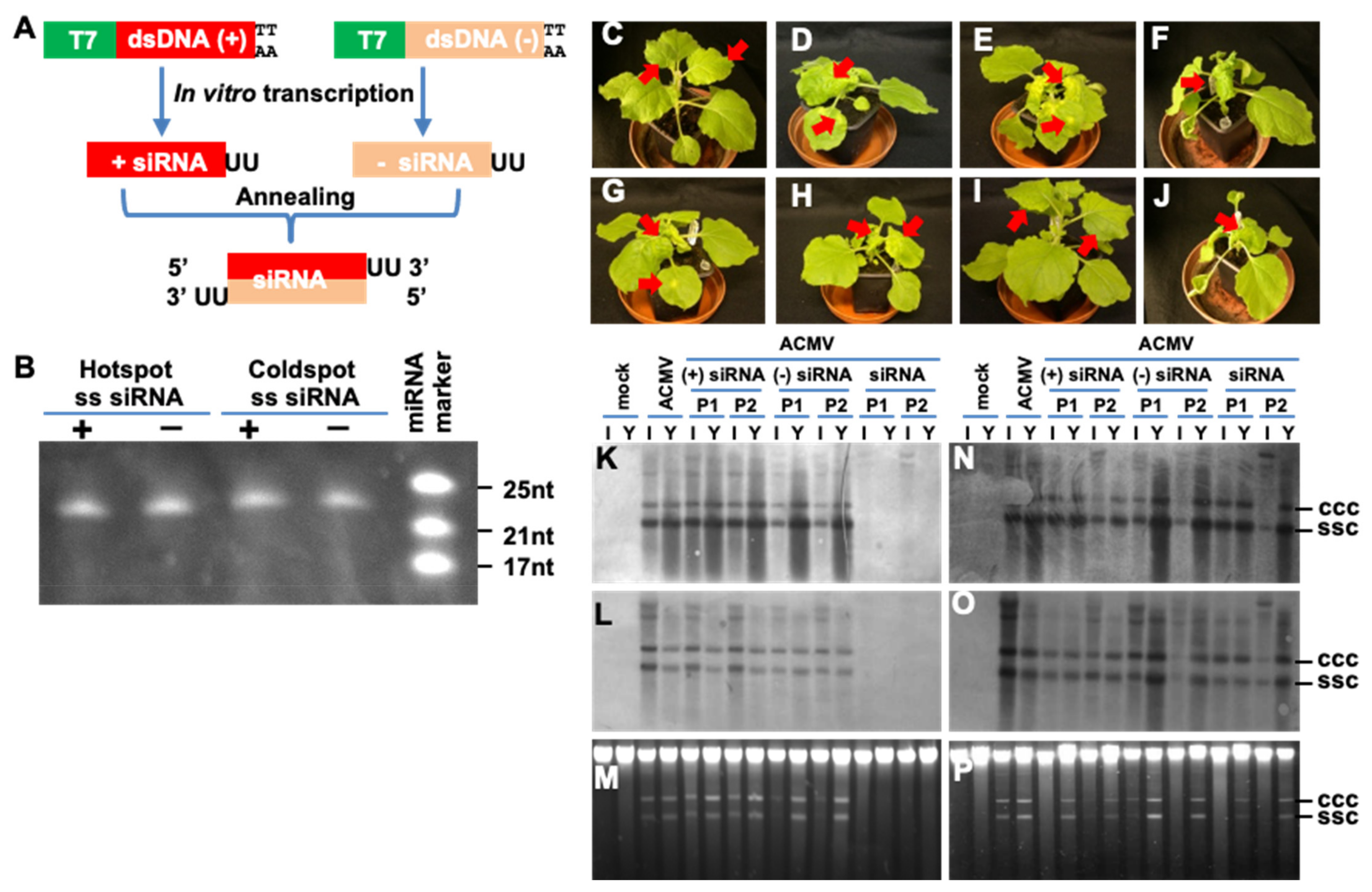
3.2. Hotspot siRNA-Mediated Viral Resistance
3.3. Threshold Effect of Hotspot ds-siRNA-Mediated ACMV Suppression
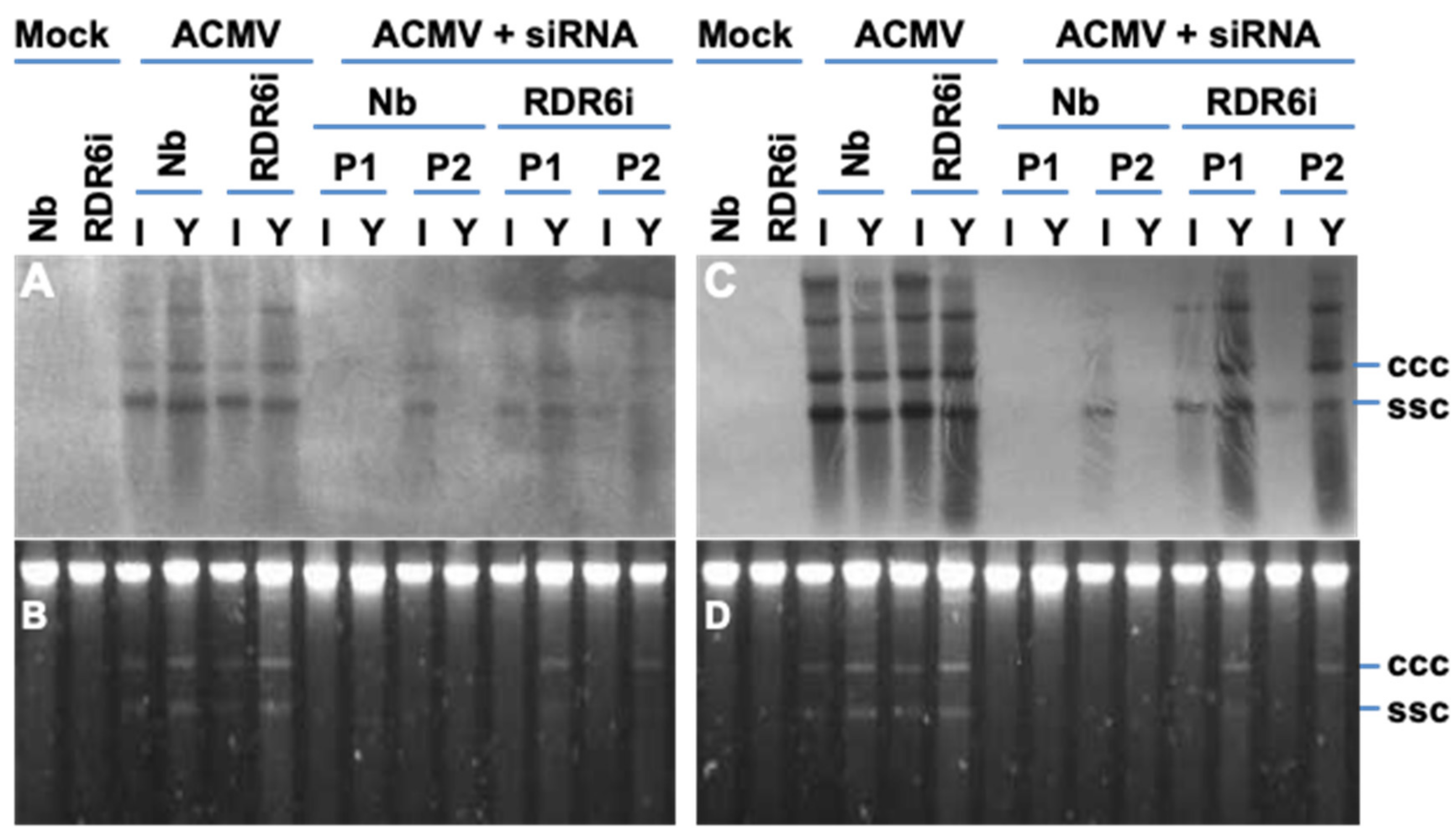
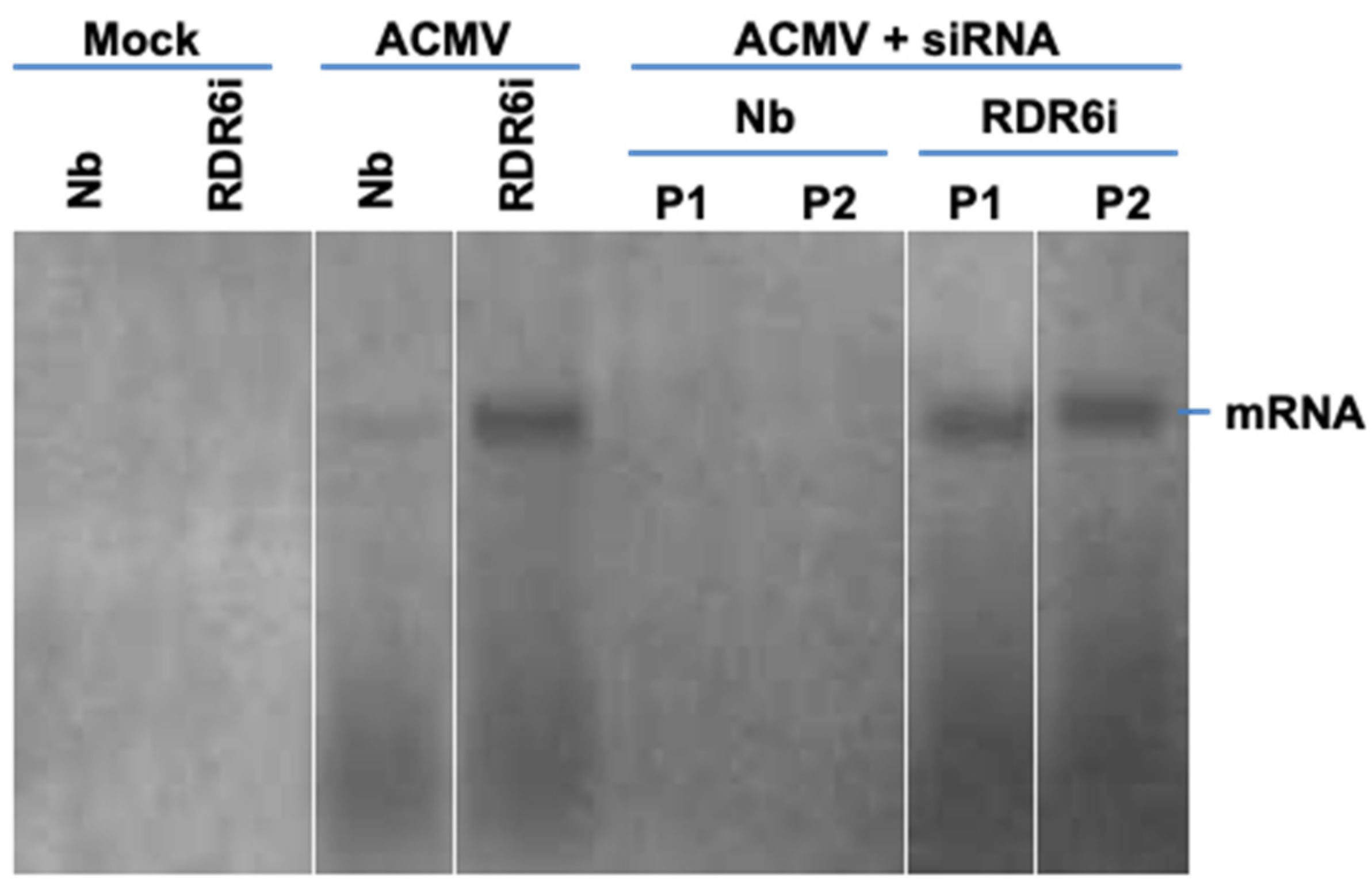
3.4. Involvement of RDR6 in Hotspot ds-siRNA-Mediated Antiviral Defence
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaire, L.; Wang, Y.; Gonzalez-Ibeas, D.; Mayer, K.F.; Aranda, M.A.; Llave, C. Deep-sequencing of plant viral small RNAs reveals effective and widespread targeting of viral genomes. Virology 2009, 392, 203–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M.; Boettcher, S.; Caudy, A.A.; Kobayashi, R.; Hannon, G.J. Argonaute2, a link between genetic and biochemical analyses of RNAi. Science 2001, 293, 1146–1150. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA deep sequencing reveals role for Arabidopsis thaliana RNA-dependent RNA polymerases in viral siRNA biogenesis. PLoS ONE 2009, 4, e4971. [Google Scholar] [CrossRef]
- Parameswaran, P.; Sklan, E.; Wilkins, C.; Burgon, T.; Samuel, M.A.; Lu, R.; Ansel, K.M.; Heissmeyer, V.; Einav, S.; Jackson, W.; et al. Six RNA viruses and forty-one hosts: Viral small RNAs and modulation of small RNA repertoires in vertebrate and invertebrate systems. PLoS Pathog. 2010, 6, e1000764. [Google Scholar] [CrossRef] [Green Version]
- Stanley, J.; Gay, M.R. Nucleotide sequence of cassava latent virus DNA. Nature 1983, 301, 260–262. [Google Scholar] [CrossRef]
- Stanley, J. Infectivity of the cloned geminivirus genome requires sequences from both DNAs. Nature 1983, 305, 643–645. [Google Scholar] [CrossRef]
- Chellappan, P.; Vanitharani, R.; Pita, J.; Fauquet, C.M. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected hosts, and gene silencing targets specific viral sequences. J. Virol. 2004, 78, 7465–7477. [Google Scholar] [CrossRef] [Green Version]
- Akbergenov, R.; Si-Ammour, A.; Blevins, T.; Amin, I.; Kutter, C.; Vanderschuren, H.; Zhang, P.; Gruissem, W.; Meins, F., Jr.; Hohn, T.; et al. Molecular characterization of geminivirus-derived small RNAs in different plant species. Nucleic Acids Res. 2006, 34, 462–471. [Google Scholar] [CrossRef]
- Hong, Y.; Saunders, K.; Hartley, M.R.; Stanley, J. Resistance to geminivirus infection by virus-induced expression of dianthin in transgenic plants. Virology 1996, 220, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Van Wezel, R.; Dong, X.; Blake, P.; Stanley, J.; Hong, Y. Differential roles of geminivirus Rep and AC4 (C4) in the induction of necrosis in Nicotiana benthamiana. Mol. Plant Pathol. 2002, 3, 461–471. [Google Scholar] [CrossRef]
- Van Wezel, R.; Dong, X.; Liu, H.; Tien, P.; Stanley, J.; Hong, Y. Mutation of three cysteine residues in Tomato yellow leaf curl virus-China C2 protein causes dysfunction in pathogenesis and posttranscriptional gene-silencing suppression. Mol. Plant Microbe Interact. 2002, 15, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.; Wang, H.; Pallett, D.; Dalmay, T. Evidence for targeting common siRNA hotspots and GC preference by plant Dicer-like proteins. FEBS Lett. 2007, 581, 3267–3272. [Google Scholar] [CrossRef]
- Hong, Y.; Stanley, J.; Van Wezel, R. Novel system for the simultaneous analysis of geminivirus DNA replication and plant interactions in Nicotiana benthamiana. J. Virol. 2003, 77, 13315–13322. [Google Scholar] [CrossRef] [Green Version]
- Vanitharani, R.; Chellappan, P.; Fauquet, C.M. Short interfering RNA-mediated interference of gene expression and viral DNA accumulation in cultured plant cells. Proc. Natl. Acad. Sci. USA 2003, 100, 9632–9636. [Google Scholar] [CrossRef] [Green Version]
- Lindbo, J.A.; Silva-Rosales, L.; Proebsting, W.M.; Dougherty, W.G. Induction of a highly specific antiviral state in transgenic plants: Implications for regulation of gene expression and virus resistance. Plant Cell 1993, 5, 1749–1759. [Google Scholar] [CrossRef]
- Dalmay, T.; Hamilton, A.; Rudd, S.; Angell, S.; Baulcombe, D.C. An RNA-dependent RNA polymerase gene in Arabidopsis is required for posttranscriptional gene silencing mediated by a transgene but not by a virus. Cell 2000, 101, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Mourrain, P.; Béclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Jouette, D.; Lacombe, A.M.; Nikic, S.; Picault, N.; et al. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Fan, B.; Chen, C.; Chen, Z. An important role of an inducible RNA-dependent RNA polymerase in plant antiviral defense. Proc. Natl. Acad. Sci. USA 2001, 98, 6516–6521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwach, F.; Vaistij, F.E.; Jones, L.; Baulcombe, D.C. An RNA-dependent RNA polymerase prevents meristem invasion by potato virus X and is required for the activity but not the production of a systemic silencing signal. Plant Physiol. 2005, 138, 1842–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muangsan, N.; Beclin, C.; Vaucheret, H.; Robertson, D. Geminivirus VIGS of endogenous genes requires SGS2/SDE1 and SGS3 and defines a new branch in the genetic pathway for silencing in plants. Plant J. 2004, 38, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Hou, G.; Sato, S.; Clemente, T.E.; Morris, T.J. RDR6 has a broad-spectrum but temperature-dependent antiviral defense role in Nicotiana benthamiana. J. Virol. 2005, 79, 15209–15217. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Shi, N.; Gu, M.; Zhang, H.; Li, B.; Shen, J.; Mohammed, A.; Ryabov, E.; Li, C.; Wang, H.; et al. Involvement of RDR6 in short-range intercellular RNA silencing in Nicotiana benthamiana. Sci Rep. 2012, 2, 467. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhang, X.; Fan, Y.; Li, B.; Ryabov, E.; Shi, N.; Zhao, M.; Yu, Z.; Qin, C.; Zheng, Q.; et al. A genetic network for systemic RNA silencing in plants. Plant Physiol. 2018, 176, 2700–2719. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef]
- Castanotto, D.; Rossi, J.J. The promises and pitfalls of RNA-interference-based therapeutics. Nature 2009, 457, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Li, B.; Fan, Y.; Zhang, X.; Yu, Z.; Ryabov, E.; Zhao, M.; Wang, H.; Shi, N.; Zhang, P.; et al. Roles of Dicer-Like Proteins 2 and 4 in Intra- and Intercellular Antiviral Silencing. Plant Physiol. 2017, 174, 1067–1081. [Google Scholar] [CrossRef]
- Zhang, X.; Lai, T.; Zhang, P.; Zhang, X.; Yuan, C.; Jin, Z.; Li, H.; Yu, Z.; Qin, C.; Tör, M.; et al. Mini review: Revisiting mobile RNA silencing in plants. Plant Sci. 2019, 278, 113–117. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, Q.; Wu, Y.; Huang, F.; Ismayil, A.; Zhang, D.; Li, H.; Gu, H.; Ludman, M.; Fátyol, K.; et al. A calmodulin-binding transcription factor links calcium signaling to antiviral RNAi defense in plants. Cell Host Microbe 2021, 29, 1393–1406.e7. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, Q.; Jin, Z.; Liu, Y.; Hong, Y. Linking calcium and RNAi signaling in plants. Trends Plant Sci. 2022, 27, 328–330. [Google Scholar] [CrossRef]
- Bilir, Ö.; Telli, O.; Norman, C.; Budak, H.; Hong, Y.; Tör, M. Small RNA inhibits infection by downy mildew pathogen Hyaloperonospora arabidopsidis. Mol. Plant. Pathol. 2019, 20, 1523–1534. [Google Scholar] [CrossRef] [Green Version]
- Abdellatef, E.; Kamal, N.M.; Tsujimoto, H. Tuning Beforehand: A foresight on RNA interference (RNAi) and in vitro-derived dsRNAs to enhance crop resilience to biotic and abiotic Stresses. Int. J. Mol. Sci. 2021, 22, 7687. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences | Application |
|---|---|---|
| PP684 | 5′TTGGAGACACTCAACTAGAGACACTCTTGAGCAT3′ | Probe A1 |
| PP685 | 5′TGACGTGGACAGTGGGGGCAGTAGCACGGTTCCTG3′ | Probe A1 |
| PP686 | 5′ACTGTCCACGTCACAAATCGAAAACGG3′ | Probe A2 |
| PP687 | 5′TCATACTTCCCTGCCTCCTGATGATTGTATGTCAC3′ | Probe A2 |
| PP688 | 5′GGCAGGGAAGTATGAGAATCACACAGAGAATG3′ | Probe A3 |
| PP689 | 5′GGATGTTCATATTACCTCCATATCAACTGCTTCAA3′ | Probe A3 |
| PP690 | 5′GTAATATGAACATCCACAGACAAGATCCACTCT3′ | Probe A4 |
| PP691 | 5′TTCCCATGTTCTTCCTTTGACCAAGTTCCTGTTGA3′ | Probe A4 |
| PP692 | 5′TCAAAGGAAGAACATGGGAAGGGAGAAACATAAG3′ | Probe A5 |
| PP693 | 5′TCTAGTTGAGTGTCTCCAATTGACTTGGTCAACATG3′ | Probe A5 |
| PP758 | 5′TAGAGAGAGACGCTCTCAACTGGAGACACACTTG3′ | Probe B1 |
| PP759 | 5′TGTATTTCGCACCCTATATATAATACATAA3′ | Probe B1 |
| PP760 | 5′ATACAGGAGTTGGAGAATATCATTTATTGGAAGTA3′ | Probe B2 |
| PP761 | 5′TATTCAATAATCTTAAATTAACGTAACAAGCGGAA3′ | Probe B2 |
| PP762 | 5′GAATAATCAACAGCATCGATATAGGGTATT3′ | Probe B3 |
| PP763 | 5′CCACAATTGGGCGCTATACAAGCATGAGAT3′ | Probe B3 |
| PP764 | 5′TGTGGATTTTGTAGCCCATGTTTCTCCTGGTT3′ | Probe B4 |
| PP765 | 5′TTCCTTCGACCCTTGAGAGAACAAGGGTACGTAT3′ | Probe B4 |
| PP766 | 5′AGGAAATTGAAGTGTAATCGGCGATTCATC3′ | Probe B5 |
| PP767 | 5′TCCAGTTGAGAGCGTCTCTCTCTAACTTTCTCTC3′ | Probe B5 |
| PP768 | 5′TAATACGACTCACTATAGCTGAGGAAGGAGCTGGGGACTT3′ | Hotspot (+) strand siRNA |
| PP769 | 3′ATTATGCTGAGTGATATCGACTCCTTCCTCGACCCCTGAA5′ | Hotspot (+) strand siRNA |
| PP770 | 5′TAATACGACTCACTATAGTCCCCAGCTCCTTCCTCAGCTT3′ | Hotspot (−) strand siRNA |
| PP771 | 3′ATTATGCTGAGTGATATCAGGGGTCGAGGAAGGAGTCGAA5′ | Hotspot (−) strand siRNA |
| PP772 | 5′TAATACGACTCACTATAGCAAGTAATTAAATACTTGGCTT3′ | Coldspot (+) strand siRNA |
| PP773 | 3′ATTATGCTGAGTGATATCGTTCATTAATTTATGAACCGAA5′ | Coldspot (+) strand siRNA |
| PP774 | 5′TAATACGACTCACTATAGCCAAGTATTTAATTACTTGCTT3′ | Coldspot (−) strand siRNA |
| PP775 | 3′ATTATGCTGAGTGATATCGGTTCATAAATTAATGAACGAA5′ | Coldspot (−) strand siRNA |
| Experiment | I | II | ||
|---|---|---|---|---|
| Hotspot | Coldspot | Hotspot | Coldspot | |
| Mock | 0/2 | 0/2 | 0/3 | 0/3 |
| ACMV | 2/2 | 2/2 | 3/3 | 3/3 |
| ACMV/(+) strand | 2/2 | 2/2 * | 3/3 | 3/3 |
| ACMV/(−) strand | 2/2 | 2/2 | 3/3 | 3/3 |
| ACMV/siRNA | 0/2 | 2/2 | 0/4 | 3/3 |
| Experiment | I | II | III | |
|---|---|---|---|---|
| Mock | Nb | 0/2 | 0/2 | 0/4 |
| NbRDR6i | 0/2 | 0/2 | 0/4 | |
| ACMV | Nb | 3/3 | 4/4 | 4/4 |
| NbRDR6i | 3/3 | 4/4 | 4/4 | |
| ACMV/ siRNA | Nb | 0/3 * | 0/4 * | 0/3 * |
| NbRDR6i | 3/3 | 4/4 | 3/3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, A.; Jin, Z.; Osman, T.; Shi, N.; Tör, M.; Jackson, S.; Hong, Y. Hotspot siRNA Confers Plant Resistance against Viral Infection. Biology 2022, 11, 714. https://doi.org/10.3390/biology11050714
Mohamed A, Jin Z, Osman T, Shi N, Tör M, Jackson S, Hong Y. Hotspot siRNA Confers Plant Resistance against Viral Infection. Biology. 2022; 11(5):714. https://doi.org/10.3390/biology11050714
Chicago/Turabian StyleMohamed, Atef, Zhenhui Jin, Toba Osman, Nongnong Shi, Mahmut Tör, Stephen Jackson, and Yiguo Hong. 2022. "Hotspot siRNA Confers Plant Resistance against Viral Infection" Biology 11, no. 5: 714. https://doi.org/10.3390/biology11050714
APA StyleMohamed, A., Jin, Z., Osman, T., Shi, N., Tör, M., Jackson, S., & Hong, Y. (2022). Hotspot siRNA Confers Plant Resistance against Viral Infection. Biology, 11(5), 714. https://doi.org/10.3390/biology11050714