
Poison or Potion: Effects of Sunflower Phenolamides on Bumble Bees and Their Gut Parasite

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
Simple Summary
Abstract
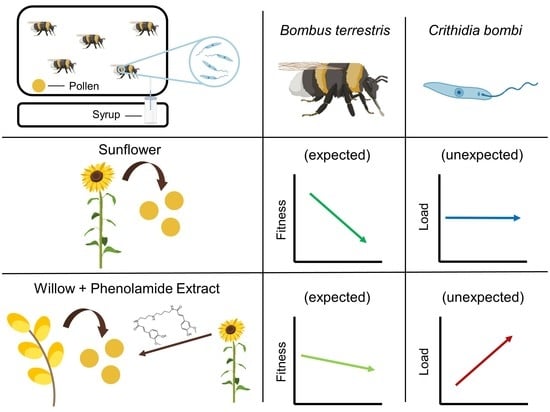
1. Introduction
2. Materials and Methods
2.1. Phenolamide Profiling in Sunflower
2.2. Bioassays
2.2.1. Experimental Design
2.2.2. Parasite Inoculation and Monitoring
2.2.3. Microcolony Parameters
2.2.4. Individual Parameters
3. Results
3.1. Phenolamides in Sunflower
3.2. Microcolony Performance
3.2.1. Resource Collection
3.2.2. Stress Responses
3.2.3. Development
3.3. Infection and Immunocompetence
3.3.1. Parasite Load
3.3.2. Fat Body Content
3.4. Phenotypic Variation
3.4.1. Centroid Size
3.4.2. Wing Shape
3.4.3. Fluctuating Asymmetry
4. Discussion
4.1. Phenolamide Allocation in Sunflower
4.2. Effects of Sunflower Pollen and Phenolamides on Bumble Bees
4.3. Infection Costs of a Gut Parasite on Bumble Bees
4.4. Effects of Sunflower Pollen and Phenolamides on a Gut Parasite
4.5. Focus for Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee nutrition and floral resource restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Palmer-Young, E.C.; Farrell, I.W.; Adler, L.S.; Milano, N.J.; Egan, P.A.; Junker, R.R.; Irwin, R.E.; Stevenson, P.C. Chemistry of floral rewards: Intra- and interspecific variability of nectar and pollen secondary metabolites across taxa. Ecol. Monogr. 2019, 89, e01335. [Google Scholar] [CrossRef]
- Westerkamp, C. Pollen in bee-flower relations some considerations on melittophily. Bot. Acta 1996, 109, 325–332. [Google Scholar] [CrossRef]
- Brochu, K.K.; van Dyke, M.T.; Milano, N.J.; Petersen, J.D.; McArt, S.H.; Nault, B.A.; Kessler, A.; Danforth, B.N. Pollen defenses negatively impact foraging and fitness in a generalist bee (Bombus impatiens: Apidae). Sci. Rep. 2020, 10, 3112. [Google Scholar] [CrossRef] [PubMed]
- Detzel, A.; Wink, M. Attraction, deterrence or intoxication of bees (Apis mellifera) by plant allelochemicals. Chemoecology 1993, 4, 8–18. [Google Scholar] [CrossRef]
- Arnold, S.E.J.; Idrovo, M.E.P.; Arias, L.J.L.; Belmain, S.R.; Stevenson, P.C. Herbivore defence compounds occur in pollen and reduce bumblebee colony fitness. J. Chem. Ecol. 2014, 40, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Hurst, V.; Stevenson, P.C.; Wright, G.A. Toxins induce ‘malaise’ behaviour in the honeybee (Apis mellifera). J. Comp. Physiol. A 2014, 200, 881–890. [Google Scholar] [CrossRef]
- Baracchi, D.; Brown, M.J.F.; Chittka, L. Behavioural evidence for self-medication in bumblebees? F1000Research 2015, 4, 73. [Google Scholar] [CrossRef] [PubMed]
- Rivest, S.; Forrest, J.R.K. Defence compounds in pollen: Why do they occur and how do they affect the ecology and evolution of bees? New Phytol. 2020, 225, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.C. For antagonists and mutualists: The paradox of insect toxic secondary metabolites in nectar and pollen. Phytochem. Rev. 2020, 19, 603–614. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Stabler, D.; Patch, H.M.; Tooker, J.F.; Grozinger, C.M.; Wright, G.A. Bumble bees regulate their intake of essential protein and lipid pollen macronutrients. J. Exp. Biol. 2016, 220, 507. [Google Scholar] [CrossRef] [PubMed]
- Vanderplanck, M.; Martinet, B.; Carvalheiro, L.G.; Rasmont, P.; Barraud, A.; Renaudeau, C.; Michez, D. Ensuring access to high-quality resources reduces the impacts of heat stress on bees. Sci. Rep. 2019, 9, 12596. [Google Scholar] [CrossRef]
- Barraud, A.; Vanderplanck, M.; Nadarajah, S.; Michez, D. The impact of pollen quality on the sensitivity of bumblebees to pesticides. Acta Oecologica 2020, 105, 103552. [Google Scholar] [CrossRef]
- Figueroa, L.L.; Grincavitch, C.; McArt, S.H. Crithidia bombi can infect two solitary bee species while host survivorship depends on diet. Parasitology 2021, 148, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Folly, A.J.; Barton-Navarro, M.; Brown, M.J.F. Exposure to nectar-realistic sugar concentrations negatively impacts the ability of the trypanosome parasite (Crithidia bombi) to infect its bumblebee host. Ecol. Entomol. 2020, 45, 1495–1498. [Google Scholar] [CrossRef]
- Macfarlane, R.P.; Lipa, J.J.; Liu, H.J. Bumble bee pathogens and internal enemies. Bee World 1995, 76, 130–148. [Google Scholar] [CrossRef]
- Meeus, I.; Brown, M.J.; de Graaf, D.C.; Smagghe, G. Effects of invasive parasites on bumble bee declines. Conserv. Biol. 2011, 25, 662–671. [Google Scholar] [CrossRef]
- Gegear, R.J.; Otterstatter, M.C.; Thomson, J.D. Bumble-bee foragers infected by a gut parasite have an impaired ability to utilize floral information. Proc. R. Soc. B Biol. Sci. 2006, 273, 1073–1078. [Google Scholar] [CrossRef]
- Graystock, P.; Meeus, I.; Smagghe, G.; Goulson, D.; Hughes, W.O.H. The effects of single and mixed infections of Apicystis bombi and deformed wing virus in Bombus terrestris. Parasitology 2016, 143, 358–365. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Loosli, R.; Schmid-Hempel, P. Condition-dependent expression of virulence in a trypanosome infecting bumblebees. Oikos 2000, 91, 421–427. [Google Scholar] [CrossRef]
- Sadd, B.M.; Barribeau, S.M. Heterogeneity in infection outcome: Lessons from a bumblebee-trypanosome system. Parasite Immunol. 2013, 35, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Woodward, J.; Langat, M.K.; Brown, M.J.F.; Stevenson, P.C. Flagellum removal by a nectar metabolite inhibits infectivity of a bumblebee parasite. Curr. Biol. 2019, 29, 3494–3500. [Google Scholar] [CrossRef] [PubMed]
- Roger, N.; Michez, D.; Wattiez, R.; Sheridan, C.; Vanderplanck, M. Diet effects on bumblebee health. J. Insect Physiol. 2017, 96, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Folly, A.J.; Stevenson, P.C.; Brown, M.J.F. Age-related pharmacodynamics in a bumblebee-microsporidian system mirror similar patterns in vertebrates. J. Exp. Biol. 2020, 223, jeb217828. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Young, E.C.; Tozkar, C.Ö.; Schwarz, R.S.; Chen, Y.; Irwin, R.E.; Adler, L.S.; Evans, J.D. Nectar and pollen phytochemicals stimulate honey bee (Hymenoptera: Apidae) immunity to viral infection. J. Econ. Entomol. 2017, 110, 1959–1972. [Google Scholar] [CrossRef]
- Giacomini, J.J.; Leslie, J.; Tarpy, D.R.; Palmer-Young, E.C.; Irwin, R.E.; Adler, L.S. Medicinal value of sunflower pollen against bee pathogens. Sci. Rep. 2018, 8, 14394. [Google Scholar] [CrossRef] [PubMed]
- LoCascio, G.M.; Aguirre, L.; Irwin, R.E.; Adler, L.S. Pollen from multiple sunflower cultivars and species reduces a common bumblebee gut pathogen. R. Soc. Open Sci. 2019, 6, 190279. [Google Scholar] [CrossRef]
- LoCascio, G.M.; Pasquale, R.; Amponsah, E.; Irwin, R.E.; Adler, L.S. Effect of timing and exposure of sunflower pollen on a common gut pathogen of bumble bees. Ecol. Entomol. 2019, 44, 702–710. [Google Scholar] [CrossRef]
- Fowler, A.E.; Stone, E.C.; Irwin, R.E.; Adler, L.S. Sunflower pollen reduces a gut pathogen in worker and queen but not male bumble bees. Ecol. Entomol. 2020, 45, 1318–1326. [Google Scholar] [CrossRef]
- Giacomini, J.J.; Connon, S.J.; Marulanda, D.; Adler, L.S.; Irwin, R.E. The costs and benefits of sunflower pollen diet on bumble bee colony disease and health. Ecosphere 2021, 12, e03663. [Google Scholar] [CrossRef]
- Adler, L.S.; Fowler, A.E.; Malfi, R.L.; Anderson, P.R.; Coppinger, L.M.; Deneen, P.M.; Lopez, S.; Irwin, R.E.; Farrell, I.W.; Stevenson, P.C. Assessing chemical mechanisms underlying the effects of sunflower pollen on a gut pathogen in bumble bees. J. Chem. Ecol. 2020, 46, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, J.J.; Moore, N.; Adler, L.S.; Irwin, R.E. Sunflower pollen induces rapid excretion in bumble bees: Implications for host-pathogen interactions. J. Insect Physiol. 2022, 137, 104356. [Google Scholar] [CrossRef]
- Roumani, M.; Besseau, S.; Gagneul, D.; Robin, C.; Larbat, R. Phenolamides in plants: An update on their function, regulation, and origin of their biosynthetic enzymes. J. Exp. Bot. 2021, 72, 2334–2355. [Google Scholar] [CrossRef] [PubMed]
- Elejalde-Palmett, C.; de Bernonville, T.D.; Glevarec, G.; Pichon, O.; Papon, N.; Courdavault, V.; St-Pierre, B.; Giglioli-Guivarc’h, N.; Lanoue, A.; Besseau, S. Characterization of a spermidine hydroxycinnamoyltransferase in Malus domestica highlights the evolutionary conservation of trihydroxycinnamoyl spermidines in pollen coat of core Eudicotyledons. J. Exp. Bot. 2015, 66, 7271–7285. [Google Scholar] [CrossRef] [PubMed]
- Handrick, V.; Vogt, T.; Frolov, A. Profiling of hydroxycinnamic acid amides in Arabidopsis thaliana pollen by tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 2789–2801. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, C.; Zhao, X.; Xia, Y.; Sun, X.; Xie, W.; Ye, Y.; Lu, X.; Xu, G. Deep annotation of hydroxycinnamic acid amides in plants based on ultra-high-performance liquid chromatography-high-resolution mass spectrometry and its in silico database. Anal. Chem. 2018, 90, 14321–14330. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wilke, C.O. Cowplot: Streamlined Plot Theme and Plot Annotations for “ggplot2”. 2020. Available online: https://cran.r-project.org/web/packages/cowplot/index.html (accessed on 10 January 2022).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. 2020. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 10 January 2022).
- R Core Team Stats. The R Stats Package. 2020. Available online: https://cran.r-project.org/web/packages/STAT/index.html (accessed on 10 January 2022).
- Husson, F.; Josse, J.; Le, S.; Mazet, J. FactoMineR: Multivariate Exploratory Data Analysis and Data Mining. 2017. Available online: https://cran.r-project.org/web/packages/FactoMineR/index.html (accessed on 10 January 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 10 January 2022).
- Martinez Arbizu, P. Pairwiseadonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 0.4. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 10 January 2022).
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology. 2019. Available online: https://cran.r-project.org/web/packages/labdsv/index.html (accessed on 10 January 2022).
- Regali, A.; Rasmont, P. Nouvelles méthodes de test pour l’évaluation du régime alimentaire chez des colonies orphelines de Bombus terrestris (L) (Hymenoptera, Apidae). Apidologie 1995, 26, 273–281. [Google Scholar] [CrossRef]
- Logan, A.; Ruiz-González, M.X.; Brown, M.J.F. The impact of host starvation on parasite development and population dynamics in an intestinal trypanosome parasite of bumble bees. Parasitology 2005, 130, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, P.; Reber Funk, C. The distribution of genotypes of the trypanosome parasite, Crithidia bombi, in populations of its host, Bombus terrestris. Parasitology 2004, 129, 147–158. [Google Scholar] [CrossRef]
- Cole, R.J. The application of the “triangulation” method to the purification of Nosema spores from insect tissues. J. Invertebr. Pathol. 1970, 15, 193–195. [Google Scholar] [CrossRef]
- Martin, C.D.; Fountain, M.T.; Brown, M.J.F. Bumblebee olfactory learning affected by task allocation but not by a trypanosome parasite. Sci. Rep. 2018, 8, 5809. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.; Kristensen, K.; van Benthem, K.; Magnusson, A.; Berg, C.; Nielsen, A.; Skaug, H.; Mächler, M.; Bolker, B. Modeling zero-inflated count data with glmmTMB. BioRxiv 2017, 12, 132753. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2021. Available online: https://cran.r-project.org/web/packages/DHARMa/index.html (accessed on 10 January 2022).
- Bartoń, K. MuMIn: Multi-Model Inference. 2020. Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 10 January 2022).
- Du Prel, J.-B.; Hommel, G.; Röhrig, B.; Blettner, M. Confidence interval or p-value? Part 4 of a series on evaluation of scientific publications. Dtsch. Arztebl. Int. 2009, 106, 335–339. [Google Scholar] [CrossRef]
- Sculfort, O.; Gérard, M.; Gekière, A.; Nonclercq, D.; Gerbaux, P.; Duez, P.; Vanderplanck, M. Specialized metabolites in floral resources: Effects and detection in buff-tailed bumblebees. Front. Ecol. Evol. 2021, 9, 669352. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear Mixed-Effects Models Using “Eigen” and S4. 2021. Available online: https://cran.r-project.org/web/packages/lme4/index.html (accessed on 10 January 2022).
- Therneau, T.M. Coxme: Mixed Effects Cox Models. 2020. Available online: https://cran.r-project.org/web/packages/coxme/index.html (accessed on 10 January 2022).
- Therneau, T.M. Survival: Survival Analysis. 2021. Available online: https://cran.r-project.org/web/packages/survival/index.html (accessed on 10 January 2022).
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Ellers, J. Fat and eggs: An alternative method to measure the trade-off between survival and reproduction in insect parasitoids. Neth. J. Zool. 1995, 46, 227–235. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Michez, D.; Albrecht, M.; Attridge, E.; Babin, A.; Bottero, I.; Breeze, T.; Brown, M.; Chauzat, M.-P.; Cini, E.; et al. Monitoring bee health in European agro-ecosystems using wing morphology and fat bodies. One Ecosyst. 2021, 6, e63653. [Google Scholar] [CrossRef]
- Gérard, M.; Michez, D.; Debat, V.; Fullgrabe, L.; Meeus, I.; Piot, N.; Sculfort, O.; Vastrade, M.; Smagghe, G.; Vanderplanck, M. Stressful conditions reveal decrease in size, modification of shape but relatively stable asymmetry in bumblebee wings. Sci. Rep. 2018, 8, 15169. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. tpsUtil Program. Version 1.47. 2013. Available online: http://www.sbmorphometrics.org/soft-utility.html (accessed on 5 January 2022).
- Rohlf, F.J. tpsDig. Version 2.30. 2013. Available online: http://sbmorphometrics.org/soft-dataacq.html (accessed on 5 January 2022).
- Adams, D.; Collyer, M.; Kaliontzopoulou, A.; Baken, E. Geomorph: Geometric Morphometric Analyses of 2D/3D Landmark Data. 2021. Available online: https://cran.r-project.org/web/packages/geomorph/index.html (accessed on 5 January 2022).
- Collyer, M.; Adams, D. RRPP: Linear Model Evaluation with Randomized Residuals in a Permutation Procedure. 2021. Available online: https://cran.r-project.org/web/packages/RRPP/index.html (accessed on 5 January 2022).
- Kyselka, J.; Bleha, R.; Dragoun, M.; Bialasová, K.; Horáčková, Š.; Schätz, M.; Sluková, M.; Filip, V.; Synytsya, A. Antifungal polyamides of hydroxycinnamic acids from sunflower bee pollen. J. Agric. Food Chem. 2018, 66, 11018–11026. [Google Scholar] [CrossRef] [PubMed]
- Sammataro, D.; Erickson, E.H.; Garment, M.B. Ultrastructure of the sunflower nectary. J. Apic. Res. 1985, 24, 150–160. [Google Scholar] [CrossRef]
- Clearwater, M.J.; Revell, M.; Noe, S.; Manley-Harris, M. Influence of genotype, floral stage, and water stress on floral nectar yield and composition of mānuka (Leptospermum scoparium). Ann. Bot. 2018, 121, 501–512. [Google Scholar] [CrossRef]
- Vogt, T. Unusual spermine-conjugated hydroxycinnamic acids on pollen: Function and evolutionary advantage. J. Exp. Bot. 2018, 69, 5311–5315. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Manson, J.S.; Gardner, D.R.; Welch, K.D.; Irwin, R.E. Norditerpene alkaloid concentrations in tissues and floral rewards of larkspurs and impacts on pollinators. Biochem. Syst. Ecol. 2013, 48, 123–131. [Google Scholar] [CrossRef]
- Stegemann, T.; Kruse, L.H.; Brütt, M.; Ober, D. Specific distribution of pyrrolizidinea alkaloids in floral parts of comfrey (Symphytum officinale) and its implications for flower ecology. J. Chem. Ecol. 2019, 45, 128–135. [Google Scholar] [CrossRef] [PubMed]
- McAulay, M.K.; Forrest, J.R.K. How do sunflower pollen mixtures affect survival of queenless microcolonies of bumblebees (Bombus impatiens)? Arthropod. Plant. Interact. 2019, 13, 517–529. [Google Scholar] [CrossRef]
- Nicolson, S.W.; Human, H. Chemical composition of the ‘low quality’ pollen of sunflower (Helianthus annuus, Asteraceae). Apidologie 2013, 44, 144–152. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Gilles, H.; Nonclercq, D.; Duez, P.; Gerbaux, P. Asteraceae paradox: Chemical and mechanical protection of Taraxacum pollen. Insects 2020, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Blackmore, S.; Wortley, A.H.; Skvarla, J.J.; Rowley, J.R. Pollen wall development in flowering plants. New Phytol. 2007, 174, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.-A.; von Roepenack-Lahaye, E.; Parr, A.; Daniels, M.J.; Dow, J.M. Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas campestris, a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms. Mol. Plant-Microbe Interact. 2001, 14, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef]
- Chahine, S.; O’Donnell, M.J. Interactions between detoxification mechanisms and excretion in Malpighian tubules of Drosophila melanogaster. J. Exp. Biol. 2011, 214, 462–468. [Google Scholar] [CrossRef]
- Yu, S.J. Detoxification mechanisms in insects. In Encyclopedia of Entomology; Springer: Dordrecht, The Netherlands, 2008; pp. 1252–1254. [Google Scholar]
- Gérard, M.; Marshall, L.; Martinet, B.; Michez, D. Impact of landscape fragmentation and climate change on body size variation of bumblebees during the last century. Ecography 2021, 44, 255–264. [Google Scholar] [CrossRef]
- Moerman, R.; Vanderplanck, M.; Fournier, D.; Jacquemart, A.-L.; Michez, D. Pollen nutrients better explain bumblebee colony development than pollen diversity. Insect Conserv. Divers. 2017, 10, 171–179. [Google Scholar] [CrossRef]
- Beasley, D.A.E.; Bonisoli-Alquati, A.; Mousseau, T.A. The use of fluctuating asymmetry as a measure of environmentally induced developmental instability: A meta-analysis. Ecol. Indic. 2013, 30, 218–226. [Google Scholar] [CrossRef]
- Debat, V.; David, P. Mapping phenotypes: Canalization, plasticity and developmental stability. Trends Ecol. Evol. 2001, 16, 555–561. [Google Scholar] [CrossRef]
- Møller, A.P. A review of developmental instability, parasitism and disease. Infect. Genet. Evol. 2006, 6, 133–140. [Google Scholar] [CrossRef]
- Schmid-Hempel, P.; Wilfert, L.; Schmid-Hempel, R. Pollinator diseases: The Bombus-Crithidia system. In Wildlife Disease Ecology: Linking Theory to Data and Application; Cambridge University Press: Cambridge, UK, 2019; pp. 3–31. ISBN 9781316479964. [Google Scholar]
- Richardson, L.L.; Adler, L.S.; Leonard, A.S.; Andicoechea, J.; Regan, K.H.; Anthony, W.E.; Manson, J.S.; Irwin, R.E. Secondary metabolites in floral nectar reduce parasite infections in bumblebees. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142471. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Moret, Y.; Schmid-Hempel, P. Activation of host constitutive immune defence by an intestinal trypanosome parasite of bumble bees. Parasitology 2003, 126, 253–260. [Google Scholar] [CrossRef][Green Version]
- Folly, A.J.; Koch, H.; Stevenson, P.C.; Brown, M.J.F. Larvae act as a transient transmission hub for the prevalent bumblebee parasite Crithidia bombi. J. Invertebr. Pathol. 2017, 148, 81–85. [Google Scholar] [CrossRef]
- Fowler, A.E.; Giacomini, J.J.; Connon, S.J.; Irwin, R.E.; Adler, L.S. Sunflower pollen reduces a gut pathogen in the model bee species, Bombus impatiens, but has weaker effects in three wild congeners. Proc. R. Soc. B Biol. Sci. 2022, 289, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.J.; Palmer-Young, E.C.; Irwin, R.E.; Adler, L.S. Food limitation affects parasite load and survival of Bombus impatiens (Hymenoptera: Apidae) infected with Crithidia (Trypanosomatida: Trypanosomatidae). Environ. Entomol. 2016, 45, 1212–1219. [Google Scholar] [CrossRef]
- Mockler, B.K.; Kwong, W.K.; Moran, N.A.; Koch, H. Microbiome structure influences infection by the parasite Crithidia bombi in bumble bees. Appl. Environ. Microbiol. 2018, 84, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bassard, J.E.; Ullmann, P.; Bernier, F.; Werck-Reichhart, D. Phenolamides: Bridging polyamines to the phenolic metabolism. Phytochemistry 2010, 71, 1808–1824. [Google Scholar] [CrossRef]
- Thorburn, L.P.; Adler, L.S.; Irwin, R.E.; Palmer-Young, E.C. Variable effects of nicotine and anabasine on parasitized bumble bees. F1000Research 2015, 4, 880. [Google Scholar] [CrossRef]
- Palmer-Young, E.C.; Thursfield, L. Pollen extracts and constituent sugars increase growth of a trypanosomatid parasite of bumble bees. PeerJ 2017, 5, e3297. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J. Self-medication in insects: Current evidence and future perspectives. Ecol. Entomol. 2014, 39, 273–280. [Google Scholar] [CrossRef]
- De Roode, J.C.; Hunter, M.D. Self-medication in insects: When altered behaviors of infected insects are a defense instead of a parasite manipulation. Curr. Opin. Insect Sci. 2018, 33, 1–6. [Google Scholar] [CrossRef]
- Reagon, M.; Snow, A.A. Cultivated Helianthus annuus (Asteraceae) volunteers as a genetic “bridge” to weedy sunflower populations in North America. Am. J. Bot. 2006, 93, 127–133. [Google Scholar] [CrossRef]
- Strange, K. USDA Foreign Agricultural Service Gain Report: Oilseeds and Products Annual 2016; GAIN, USDA FAS: Washington, DC, USA, 2016.
- Murat Asla, M.; Yavuksuz, C. Effect of honey bee (Apis mellifera L.) and bumblebee (Bombus terrestris L.) pollinators on yield and yield factors in sunflower (Helianthus annuus L.) production areas. J. Anim. Vet. Adv. 2010, 9, 332–335. [Google Scholar] [CrossRef]
- Meynié, S.; Bernard, R. Efficacité comparée de la pollinisation d’espèces sauvages d’Helianthus par plusieurs genres d’insectes. Agronomie 1997, 17, 43–51. [Google Scholar] [CrossRef]
- Fell, R.D. Foraging behaviors of Apis mellifera L. and Bombus spp. on oilseed sunflower (Helianthus annuus L.). J. Kans. Entomol. Soc. 1986, 59, 72–81. [Google Scholar]
- Macoy, D.M.; Kim, W.-Y.; Lee, S.Y.; Kim, M.G. Biosynthesis, physiology, and functions of hydroxycinnamic acid amides in plants. Plant Biotechnol. Rep. 2015, 9, 269–278. [Google Scholar] [CrossRef]
- Anyanga, M.O.; Farman, D.I.; Ssemakula, G.N.; Mwanga, R.O.M.; Stevenson, P.C. Effects of hydroxycinnamic acid esters on sweetpotato weevil feeding and oviposition and interactions with Bacillus thuringiensis proteins. J. Pest Sci. 2021, 94, 783–794. [Google Scholar] [CrossRef]
- Blagbrough, I.S.; Brackley, P.T.H.; Bruce, M.; Bycroft, B.W.; Mather, A.J.; Millington, S.; Sudan, H.L.; Usherwood, P.N.R. Arthropod toxins as leads for novel insecticides: An assessment of polyamine amides as glutamate antagonists. Toxicon 1992, 30, 303–322. [Google Scholar] [CrossRef]
- Fixon-Owoo, S.; Levasseur, F.; Williams, K.; Sabado, T.N.; Lowe, M.; Klose, M.; Joffre Mercier, A.; Fields, P.; Atkinson, J. Preparation and biological assessment of hydroxycinnamic acid amides of polyamines. Phytochemistry 2003, 63, 315–334. [Google Scholar] [CrossRef]
- Rasmont, P.; Coppee, A.; Michez, D.; De Meulemeester, T. An overview of the Bombus terrestris (L. 1758) subspecies (Hymenoptera: Apidae). Ann. Société Entomol. Fr. 2008, 44, 243–250. [Google Scholar] [CrossRef]
- Velthuis, H.H.W.; van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef]
- Pendrel, B.A.; Plowright, R.C. Larval feeding by adult bumble bee workers (Hymenoptera: Apidae). Behav. Ecol. Sociobiol. 1981, 8, 71–76. [Google Scholar] [CrossRef]
- Goulson, D. Bumblebees: Behaviour, Ecology, and Conservation; Oxford University Press: New York, NY, USA, 2010.
- Shykoff, J.A.; Schmid-Hempel, P. Incidence and effects of four parasites in natural populations of bumble bees in Switzerland. Apidologie 1991, 22, 117–125. [Google Scholar] [CrossRef]
- Graystock, P.; Goulson, D.; Hughes, W.O.H. Parasites in bloom: Flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151371. [Google Scholar] [CrossRef]
- Fauser, A.; Sandrock, C.; Neumann, P.; Sadd, B.M. Neonicotinoids override a parasite exposure impact on hibernation success of a key bumblebee pollinator. Ecol. Entomol. 2017, 42, 306–314. [Google Scholar] [CrossRef]
- Goulson, D.; O’Connor, S.; Park, K.J. The impacts of predators and parasites on wild bumblebee colonies. Ecol. Entomol. 2018, 43, 168–181. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Schmid-Hempel, R.; Schmid-Hempel, P. Strong context-dependent virulence in a host-parasite system: Reconciling genetic evidence with theory. J. Anim. Ecol. 2003, 72, 994–1002. [Google Scholar] [CrossRef]
- Brunner, F.S.; Schmid-Hempel, P.; Barribeau, S.M. Immune gene expression in Bombus terrestris: Signatures of infection despite strong variation among populations, colonies, and sister workers. PLoS ONE 2013, 8, e68181. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf | Corolla | Nectar | Pollen | |
|---|---|---|---|---|
| Plant 1 | 0 (<LOD) | 0 (<LOD) | 9.19 | 10.39 |
| Plant 2 | 0 (<LOD) | 0 (<LOD) | 24.8 | 10.94 |
| Plant 3 | 0 (<LOD) | 0 (<LOD) | 0.83 | 25.38 |
| Plant 4 | 0 (<LOD) | 0 (<LOD) | 21.07 | 11.92 |
| Plant 5 | 0 (<LOD) | 0 (<LOD) | 4.77 | 12.62 |
| Mean ± SD Median (Min–Max) | 0 0 a | 0 0 a | 12.12 ± 10.38 9.19 (0.83–24.8) b | 14.25 ± 6.18 11.92 (10.39–25.38) b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gekière, A.; Semay, I.; Gérard, M.; Michez, D.; Gerbaux, P.; Vanderplanck, M. Poison or Potion: Effects of Sunflower Phenolamides on Bumble Bees and Their Gut Parasite. Biology 2022, 11, 545. https://doi.org/10.3390/biology11040545
Gekière A, Semay I, Gérard M, Michez D, Gerbaux P, Vanderplanck M. Poison or Potion: Effects of Sunflower Phenolamides on Bumble Bees and Their Gut Parasite. Biology. 2022; 11(4):545. https://doi.org/10.3390/biology11040545
Chicago/Turabian StyleGekière, Antoine, Irène Semay, Maxence Gérard, Denis Michez, Pascal Gerbaux, and Maryse Vanderplanck. 2022. "Poison or Potion: Effects of Sunflower Phenolamides on Bumble Bees and Their Gut Parasite" Biology 11, no. 4: 545. https://doi.org/10.3390/biology11040545
APA StyleGekière, A., Semay, I., Gérard, M., Michez, D., Gerbaux, P., & Vanderplanck, M. (2022). Poison or Potion: Effects of Sunflower Phenolamides on Bumble Bees and Their Gut Parasite. Biology, 11(4), 545. https://doi.org/10.3390/biology11040545