
Digestive Enzyme Activities and Gut Emptying Are Correlated with the Reciprocal Regulation of TRPA1 Ion Channel and Serotonin in the Gut of the Sea Urchin Strongylocentrotus intermedius



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sea Urchins
2.2. Chemicals
2.3. Sample Collection for TRPA1 Expression, 5-HT Concentration and Enzyme Activities
2.4. Total RNA Extraction and cDNA Synthesis
2.5. Transcriptional Analysis of TRPA1
2.6. Analysis of 5-HT Concentration
2.7. Analysis of Digestive Enzyme Activities
2.8. Statistical Analysis
3. Results
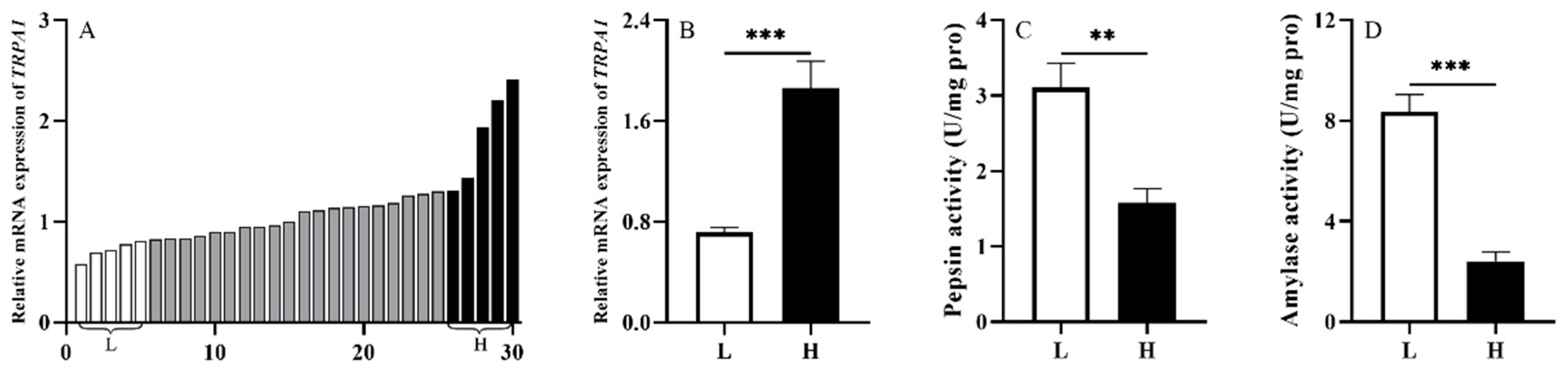
3.1. Experiment 1: TRPA1 Is Involved in the Food Digestion of S. intermedius
3.2. Experiment 2: An Interregulatory Relationship Exists between TRPA1 and 5-HT in the Gut of S. intermedius
3.3. Experiment 3: Digestive Enzyme Activities Are Negatively Correlated with TRPA1 and Concentration of 5-HT in the Gut of S. intermedius
3.4. Experiment 4: Gut Emptying Is Positively Correlated with TRPA1 and Concentration of 5-HT in the Gut of S. intermedius
4. Discussion
4.1. TRPA1 Is Involved in the Digestion of S. intermedius
4.2. An Inter-Regulatory Relationship Exists between TRPA1 and 5-HT in the Gut of S. intermedius
4.3. Digestive Enzyme Activities Are Negatively Correlated with the TRPA1 and Concentration of 5-HT in the Gut of S. intermedius
4.4. Gut Emptying Is Positively Correlated with the TRPA1 and Concentration of 5-HT in the Gut of S. intermedius
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pearse, J. Ecological role of purple sea urchins. Science 2006, 314, 940–941. [Google Scholar] [CrossRef] [PubMed]
- Yorke, C.E.; Page, H.M.; Miller, R.J. Sea urchins mediate the availability of kelp detritus to benthic consumers. Proc. R. Soc. B 2019, 286, 20190846. [Google Scholar] [CrossRef] [PubMed]
- Agatsuma, Y.; Takagi, S.; Inomata, E.; Aoki, M.N. Process of deterioration of a kelp (Ecklonia bicyclis Kjellman) bed as a result of grazing by the sea urchin Mesocentrotus nudus (Agassiz) in shizugawa bay in northeastern Honshu, Japan. J. Appl. Phycol. 2018, 31, 599–605. [Google Scholar] [CrossRef]
- Lawrence, J.M. On the relationships between marine plants and sea urchins. Oceanogr. Mar. Biol. 1975, 13, 213–286. [Google Scholar]
- Lawrence, J.M.; Lawrence, A.L.; Watts, S.A. Feeding, digestion and digestibility in sea urchins. In Sea Urchins: Biology and Ecology, 3rd ed.; Lawrence, J.M., Ed.; Academic Press: San Diego, CA, USA, 2013; pp. 135–154. [Google Scholar] [CrossRef]
- Lawrence, J.M.; Lawrence, A.L.; Watts, S.A. Ingestion, digestion, and digestibility of regular sea urchins. In Sea Urchins: Biology and Ecology, 4th ed.; Lawrence, J.M., Ed.; Academic Press: San Diego, CA, USA, 2020; pp. 165–190. [Google Scholar] [CrossRef]
- Klinger, T.S.; Mcclintock, J.B.; Watts, S.A. Activities of digestive enzymes of polar and subtropical echinoderms. Polar. Biol. 1997, 18, 154–157. [Google Scholar] [CrossRef]
- Larson, B.R.; Vadas, R.L.; Keser, M. Feeding and nutritional ecology of the sea urchin Strongylocentrotus drobachiensis in Maine, USA. Mar. Biol. 1980, 59, 49–62. [Google Scholar] [CrossRef]
- Mamelona, J.; Pelletier, E. Green urchin as a significant source of fecal particulate organic matter within nearshore benthic ecosystems. J. Exp. Mar. Biol. Ecol. 2005, 314, 163–174. [Google Scholar] [CrossRef]
- Peduzzi, P.; Herndl, G. Role of bacteria in decomposition of fecal pellets egested by the epiphyte-grazing gastropod Gibbula umbilicaris. Mar. Biol. 1986, 92, 417–424. [Google Scholar] [CrossRef]
- Yoon, W.D.; Marty, J.C.; Sylvain, D.; Nival, P. Degradation of faecal pellets in Pegea confoederata (Salpidae, Thaliacea) and its implication in the vertical flux of organic matter. J. Exp. Mar. Biol. Ecol. 1996, 203, 147–177. [Google Scholar] [CrossRef]
- Vadas, R. Preferential feeding: An optimization strategy in sea urchins. Ecol. Monogr. 1977, 47, 337–371. [Google Scholar] [CrossRef]
- Sauchyn, L.; Lauzon-Guay, J.; Scheibling, R. Sea urchin fecal production and accumulation in a rocky subtidal ecosystem. Aquat. Biol. 2011, 13, 215–223. [Google Scholar] [CrossRef]
- Dethier, M.N.; Hoins, G.; Kobelt, J.; Lowe, A.T.; Galloway, A.W.E.; Schram, J.B.; Raymore, M.; Duggins, D.O. Feces as food: The nutritional value of urchin feces and implications for benthic food webs. J. Exp. Mar. Biol. Ecol. 2019, 514, 95–102. [Google Scholar] [CrossRef]
- Hou, S.Q.; Zuo, R.T.; Chang, Y.Q.; Li, G.; Song, J.; Zhao, C. Effects of dietary lipid level on growth performance, digestive enzyme activity and anti-oxidative capacity after heat stress in juvenile sea urchin Strongylocentrotus intermedius. J. Dalian Fish. Univ. 2016, 31, 538–543, (In Chinese with an English abstract). [Google Scholar] [CrossRef]
- Onitsuka, T.; Niwa, K.; Unuma, T.; Umezu, Y. Dietary shifts in the juvenile sea urchin Strongylocentrotus intermedius associated with the development of digestive enzymes. Mar. Biol. 2015, 162, 869–880. [Google Scholar] [CrossRef]
- Horowitz, M.; O’Donovan, D.; Jones, K.L.; Feinle, C.; Rayner, C.K.; Samsom, M. Gastric emptying in diabetes: Clinical significance and treatment. Diabet. Med. 2002, 19, 177–194. [Google Scholar] [CrossRef]
- Mahé, S.; Roos, N.; Benamouzig, R.; Davin, L.; Luengo, C.; Gagnon, L.; Gaussergès, N.; Rautureau, J.; Tomé, D. Gastrojejunal kinetics and the digestion of [15N] ß-lactoglobulin and casein in humans: The influence of the nature and quantity of the protein. Am. J. Clin. Nutr. 1996, 63, 546–552. [Google Scholar] [CrossRef]
- Rehner, G.; Heil, M.; Auge, M.; Harzer, G.; Daniel, H. Effect of proteins on availability of zinc. I. Gastrointestinal transit time of casein and whey protein and zinc absorption in weaned rats. Z. Ernährungswiss 1985, 24, 245–255. [Google Scholar] [CrossRef]
- Lawrence, J.M.; McClintock, J. Energy acquisition and allocation by Echinoderms (Echinodermata) in polar seas: Adaptations for success? In Echinoderms through Time; David, B., Guille, A., Feral, J.P., Roux, M., Eds.; Balkema: Rotterdam, The Netherlands, 1994; pp. 39–52. [Google Scholar]
- McClintock, J. Trophic biology of antarctic shallow-water echinoderms. Mar. Ecol. Prog. Ser. 1994, 111, 119–202. [Google Scholar] [CrossRef]
- Damann, N.; Voets, T.; Nilius, B. TRPs in our senses. Curr. Biol. 2008, 18, 880–889. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef]
- Pedersen, S.F.; Owsianik, G.; Nilius, B. TRP channels: An overview. Cell Calcium 2005, 38, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. Trpa1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Y.; Yu, M.G.; Yu, S.Y. Trp channel functions in the gastrointestinal tract. Semin. Immunopathol. 2015, 38, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.Y.; Yu, Y.S.; Yang, M.F.; Shi, D.T.; Li, Z.Q.; Chi, X.M.; Chang, Y.Q.; Wang, Q.Z.; Zhao, C. TRPA1 expression provides new insights into thermal perception by the sea urchin Strongylocentrotus intermedius. J. Mar. Biol. Assoc. UK 2019, 99, 1825–1829. [Google Scholar] [CrossRef]
- Grundy, D. 5-HT system in the gut: Roles in the regulation of visceral sensitivity and motor functions. Eur. Rev. Med. Pharmacol. 2008, 12, 63–67. [Google Scholar] [CrossRef]
- Li, T. Effect of 5-HT on the Intestinal Motility of Eriocheir sinensis. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2016. CNKI:CDMD:2.1016.912281. [Google Scholar]
- Harris, M.N.; Shafinaz, M.N.M.; Yaakop, S.; Wahida, O.N. Distribution of serotonin (5-HT) and dopamine (DA) on digestive tract of red palm weevil larva, Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae). Serangga 2016, 21, 39–50. [Google Scholar]
- Doihara, H.; Nozawa, K.; Kawabata-Shoda, E.; Kojima, R.; Yokoyama, T.; Ito, H. Trpa1 agonists delay gastric emptying in rats through serotonergic pathways. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 380, 353–357. [Google Scholar] [CrossRef]
- Carroll, N.K.; Scully, T.K.; Brown, K.M. Serotonin signaling initiates gastrulation in the sea urchin. Dev. Biol. 2010, 344, 451. [Google Scholar] [CrossRef][Green Version]
- Eid, S.R.; Crown, E.D.; Moore, E.L.; Liang, H.A.; Urban, M.O. Hc-030031, a trpa1 selective antagonist, attenuates inflammatory- and neuropathy-induced mechanical hypersensitivity. Mol. Pain 2008, 4, 48. [Google Scholar] [CrossRef]
- Han, L.S.; Ding, J.; Wang, H.; Zuo, R.T.; Quan, Z.J.; Fan, Z.H.; Liu, Q.D.; Chang, Y.Q. Molecular characterization and expression of SiFad1 in sea urchin (Strongylocentrotus intermedius). Gene 2019, 705, 133–141. [Google Scholar] [CrossRef]
- Zhou, Z.C.; Bao, Z.M.; Dong, Y.; Wang, L.M.; He, C.B.; Liu, W.D. MYP gene expressions at transcription level in different stages of gonads of sea urchin Strongylocentrotus intermedius and hybrids. Hereditas 2008, 30, 1453–1458. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Han, Y.; Ren, T.; Jiang, Z.; Wang, F.; Liao, M.; Wang, J. Effects of dietary lysine levels on growth, intestinal digestive enzymes, and coelomic fluid nonspecific immune enzymes of sea cucumber, Apostichopus japonicus, juveniles. J. World Aquac. Soc. 2016, 48, 290–302. [Google Scholar] [CrossRef]
- Chang, Y.Q.; Ding, J.; Song, J.; Yang, W. Biology and Aquaculture of Sea Cucumbers and Sea Urchins; China Ocean Press: Beijing, China, 2004. (In Chinese) [Google Scholar]
- Brothers, C.J.; Pol, W.J.V.D.; Morrow, C.D.; Hakim, J.A.; Koo, H.; McClintock, J.B. Ocean warming alters predicted microbiome functionality in a common sea urchin. Proc. R. Soc. B 2018, 285, 20180340. [Google Scholar] [CrossRef]
- Fothergill, L.J.; Callaghan, B.; Rivera, L.R.; Lieu, T.M.; Poole, D.P.; Cho, H.J.; Bravo, D.M.; Furness, J.B. Effects of food components that activate trpa1 receptors on mucosal ion transport in the mouse intestine. Nutrients 2016, 8, 623. [Google Scholar] [CrossRef]
- Sauchyn, L.; Scheibling, R. Degradation of sea urchin feces in a rocky subtidal ecosystem: Implications for nutrient cycling and energy flow. Aquat. Biol. 2009, 6, 99–108. [Google Scholar] [CrossRef]
- Ohara, K.; Fukuda, T.; Ishida, Y.; Takahashi, C.; Ohya, R.; Katayama, M.; Uchida, K.; Tominaga, M.; Nagai, K. β-Eudesmol, an oxygenized sesquiterpene, stimulates appetite via TRPA1 and the autonomic nervous system. Sci. Rep. 2017, 7, 15785. [Google Scholar] [CrossRef]
- Shen, J.; Niijima, A.; Tanida, M.; Horii, Y.; Maeda, K.; Nagai, K. Olfactory stimulation with scent of lavender oil affects autonomic nerves, lipolysis and appetite in rats. Neurosci. Lett. 2005, 383, 188–193. [Google Scholar] [CrossRef]
- Coleman, N.S.; Foley, S.; Dunlop, S.P.; Wheatcroft, J.; Blackshaw, E.; Perkins, A.C.; Singh, G.; Marsden, C.A.; Holmes, G.K.; Spiller, R.C. Abnormalities of serotonin metabolism and their relation to symptoms in untreated celiac disease. Clin. Gastroenterol. Hepatol. 2006, 4, 874–881. [Google Scholar] [CrossRef]
- Kim, M.J.; Son, H.J.; Song, S.H.; Jung, M.; Kim, Y.; Rhyu, M.R. The TRPA1 agonist, methyl syringate suppresses food intake and gastric emptying. PLoS ONE 2013, 8, e71603. [Google Scholar] [CrossRef]
- Holzer, P. Transient receptor potential (TRP) channels as drug targets for diseases of the digestive system. Pharmacol. Ther. 2011, 131, 142–170. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.; Critchley, D.; Dourish, C.T.; Cooper, S.J. Comparisons between the effects of 5-HT and dl-fenfluramine on food intake and gastric emptying in the rat. Pharmacol. Biochem. Behav. 1995, 50, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M.; Mckinzie, S.; Fox, J.; Foxxorenstein, A.; Burton, D.; Thomforde, G.; Baxter, K.; Zinsmeister, A. Effect of renzapride on transit in constipation-predominant irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2004, 2, 895–904. [Google Scholar] [CrossRef]
- Degen, L.; Petrig, C.; Studer, D.; Schroller, S.; Beglinger, C. Effect of tegaserod on gut transit in male and female subjects. Neurogastroent. Motil. 2006, 17, 821–826. [Google Scholar] [CrossRef]
- Klinger, T.S.; Hsieh, H.L.; Pangallo, R.A.; Chen, C.P.; Lawrence, J.M. The Effect of Temperature on Feeding, Digestion, and Absorption of Lytechinus Variegatus (Lamarck) (Echinodermata, Echinoidea). Physiol. Zool. 1986, 59, 332–336. [Google Scholar] [CrossRef]
- McBride, S.C.; Pinnix, W.D.; Lawrence, J.M.; Lawrence, A.L.; Mulligan, T.J. The effects of temperature on production of gonads by the sea urchin Strongylocentrotus franciscanus fed natural and prepared diets. J. World Aquacult. Soc. 1997, 28, 357–365. [Google Scholar] [CrossRef]
- Siikavuopio, S.I.; Christainsen, J.S.; Dale, T. Effects of temperature and season on gonad growth and feed intake in the green sea urchin (Strongylocentrotus droebachiensis). Aquaculture 2006, 255, 389–394. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) | Application |
|---|---|---|
| TRPA1-F | GCCACCGCAGTCGTGTGTG | qPCR |
| TRPA1-R | TGGGCGTGGTCCGATAGTTAGTCTC | qPCR |
| 18S rRNA-F | GTTCGAAGGCGATCAGATAC | Reference gene |
| 18S rRNA-R | CTGTCAATCCTCACTGTGTC | Reference gene |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, J.; Wang, H.; Li, Z.; Sun, J.; Ding, P.; Chi, X.; Yang, M.; Chang, Y.; Zhao, C. Digestive Enzyme Activities and Gut Emptying Are Correlated with the Reciprocal Regulation of TRPA1 Ion Channel and Serotonin in the Gut of the Sea Urchin Strongylocentrotus intermedius. Biology 2022, 11, 503. https://doi.org/10.3390/biology11040503
Ding J, Wang H, Li Z, Sun J, Ding P, Chi X, Yang M, Chang Y, Zhao C. Digestive Enzyme Activities and Gut Emptying Are Correlated with the Reciprocal Regulation of TRPA1 Ion Channel and Serotonin in the Gut of the Sea Urchin Strongylocentrotus intermedius. Biology. 2022; 11(4):503. https://doi.org/10.3390/biology11040503
Chicago/Turabian StyleDing, Jingyun, Huiyan Wang, Zequn Li, Jiangnan Sun, Peng Ding, Xiaomei Chi, Mingfang Yang, Yaqing Chang, and Chong Zhao. 2022. "Digestive Enzyme Activities and Gut Emptying Are Correlated with the Reciprocal Regulation of TRPA1 Ion Channel and Serotonin in the Gut of the Sea Urchin Strongylocentrotus intermedius" Biology 11, no. 4: 503. https://doi.org/10.3390/biology11040503
APA StyleDing, J., Wang, H., Li, Z., Sun, J., Ding, P., Chi, X., Yang, M., Chang, Y., & Zhao, C. (2022). Digestive Enzyme Activities and Gut Emptying Are Correlated with the Reciprocal Regulation of TRPA1 Ion Channel and Serotonin in the Gut of the Sea Urchin Strongylocentrotus intermedius. Biology, 11(4), 503. https://doi.org/10.3390/biology11040503