
The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Data Sources and Search Strategy
2.2. Elegibility Criteria
2.3. Data Extraction and Synthesis
2.4. Data Items
2.5. Risk of Bias within Studies
3. Results
3.1. Identification of Studies
3.2. Study Characteristics
3.3. Outcome Measures of the Included Studies
4. Discussion
4.1. Novelty of This Review
4.2. Different Types of PA/PE and Their Influence on GM
4.3. Gut Microbiota and Different Physical Activity Levels
4.4. Age-Related Changes on GM Induced by PA and How Long They Persist
4.5. Limitations and Future Recommendations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferreiro, A.; Crook, N.; Gasparrini, A.J.; Dantas, G. Multiscale Evolutionary Dynamics of Host-Associated Microbiomes. Cell 2018, 172, 1216–1227. [Google Scholar] [PubMed] [Green Version]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The Human Microbiota in Health and Disease. Engineering 2017, 1, 71–82. [Google Scholar]
- Ussar, S.; Fujisaka, S.; Kahn, C.R. Interactions between host genetics and gut microbiome in diabetes and metabolic syndrome. Mol. Metab. 2016, 5, 795–803. [Google Scholar] [PubMed]
- Heiman, M.L.; Greenway, F.L. A healthy gastrointestinal microbiome is dependent on dietary diversity. Mol. Metab. 2016, 5, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Lim, J.Y.; Kim, B.S.; Cho, S.J.; Kim, N.Y.; Kim, O.B.; Kim, Y. Comparison of the gut microbiota profile in breast-fed and formula-fed Korean infants using pyrosequencing. Nutr. Res. Pract. 2015, 9, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, C.E.; Renz, H.; Jenmalm, M.C.; Kozyrskyj, A.L.; Allen, K.J.; Vuillermin, P.; Prescott, S.L.; in-FLAME Microbiome Interest Group. The gut microbiota and inflammatory noncommunicable diseases: Associations and potentials for gut microbiota therapies. J. Allergy Clin. Immunol. 2015, 135, 3–14. [Google Scholar]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the human microbiome. Nutr. Rev. 2012, 70 (Suppl. S1), S38–S44. [Google Scholar]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.; Sandhu, K.V.; Bastiaanssen, T.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar]
- Savage, D.C. Microbial ecology of the gastrointestinal tract. Annu. Rev. Microbiol. 1977, 31, 107–133. [Google Scholar] [CrossRef]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 42. [Google Scholar]
- Campbell, S.C.; Wisniewski, P.J.; Noji, M.; McGuinness, L.R.; Häggblom, M.M.; Lightfoot, S.A.; Joseph, L.B.; Kerkhof, L.J. The Effect of Diet and Exercise on Intestinal Integrity and Microbial Diversity in Mice. PLoS ONE 2016, 11, e0150502. [Google Scholar]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef]
- Dalton, A.; Mermier, C.; Zuhl, M. Exercise influence on the microbiome-gut-brain axis. Gut Microbes 2019, 10, 555–568. [Google Scholar]
- Schlegel, P.; Novotny, M.; Klimova, B.; Valis, M. “Muscle-Gut-Brain Axis”: Can Physical Activity Help Patients with Alzheimer’s Disease Due to Microbiome Modulation? J. Alzheimer’s Dis. 2019, 71, 861–878. [Google Scholar]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Front. Physiol. 2016, 7, 51. [Google Scholar]
- Mailing, L.J.; Allen, J.M.; Buford, T.W.; Fields, C.J.; Woods, J.A. Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc. Sport Sci. Rev. 2019, 47, 75–85. [Google Scholar]
- Heenam, S.K. Do an Altered Gut Microbiota and an Associated Leaky Gut Affect COVID-19 Severity? mBio 2021, 12, e03022-20. [Google Scholar]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the Gut Microbiota in Patients with COVID-19 or H1N1 Influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. J. Clin. Epidemiol. 2009, 62, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, H.; Tanisawa, K.; Sun, X.; Kubo, T.; Hoshino, Y.; Hosokawa, M.; Takeyama, H.; Higuchi, M. Effects of short-term endurance exercise on gut microbiota in elderly men. Physiol. Rep. 2018, 6, e13935. [Google Scholar] [CrossRef]
- Kern, T.; Blond, M.B.; Hansen, T.H.; Rosenkilde, M.; Quist, J.S.; Gram, A.S.; Ekstrøm, C.T.; Hansen, T.; Stallknecht, B. Structured exercise alters the gut microbiota in humans with overweight and obesity-A randomized controlled trial. Int. J. Obes. 2020, 44, 125–135. [Google Scholar]
- Motiani, K.K.; Collado, M.C.; Eskelinen, J.J.; Virtanen, K.A.; Löyttyniemi, E.; Salminen, S.; Nuutila, P.; Kalliokoski, K.K.; Hannukainen, J.C. Exercise Training Modulates Gut Microbiota Profile and Improves Endotoxemia. Med. Sci. Sports Exerc. 2020, 52, 94–104. [Google Scholar]
- Liu, Y.; Wang, Y.; Ni, Y.; Cheung, C.; Lam, K.; Wang, Y.; Xia, Z.; Ye, D.; Guo, J.; Tse, M.A.; et al. Gut Microbiome Fermentation Determines the Efficacy of Exercise for Diabetes Prevention. Cell Metab. 2020, 31, 77–91.e5. [Google Scholar]
- Quiroga, R.; Nistal, E.; Estébanez, B.; Porras, D.; Juárez-Fernández, M.; Martínez-Flórez, S.; García-Mediavilla, M.V.; de Paz, J.A.; González-Gallego, J.; Sánchez-Campos, S.; et al. Exercise training modulates the gut microbiota profile and impairs inflammatory signalling pathways in obese children. Exp. Mol. Med. 2020, 52, 1048–1061. [Google Scholar] [CrossRef]
- Resende, A.S.; Leite, G.; Lancha Junior, A.H. Changes in the Gut Bacteria Composition of Healthy Men with the Same Nutritional Profile Undergoing 10-Week Aerobic Exercise Training: A Randomized Controlled Trial. Nutrients 2021, 13, 2839. [Google Scholar] [CrossRef]
- Zhong, F.; Wen, X.; Yang, M.; Lai, H.Y.; Momma, H.; Cheng, L.; Sun, X.; Nagatomi, R.; Huang, C. Effect of an 8-week Exercise Training on Gut Microbiota in Physically Inactive Older Women. Int. J. Sports Med. 2021, 42, 610–623. [Google Scholar]
- Moitinho-Silva, L.; Wegener, M.; May, S.; Schrinner, F.; Akhtar, A.; Boysen, T.J.; Schaeffer, E.; Hansen, C.; Schmidt, T.; Rühlemann, M.C.; et al. Short-term physical exercise impacts on the human holobiont obtained by a randomised intervention study. BMC Microbiol. 2021, 21, 162. [Google Scholar] [CrossRef]
- Munukka, E.; Ahtiainen, J.P.; Puigbó, P.; Jalkanen, S.; Pahkala, K.; Keskitalo, A.; Kujala, U.M.; Pietilä, S.; Hollmén, M.; Elo, L.; et al. Six-Week Endurance Exercise Alters Gut Metagenome That Is not Reflected in Systemic Metabolism in Over-weight Women. Front. Microbiol. 2018, 9, 2323. [Google Scholar] [CrossRef]
- Morita, E.; Yokoyama, H.; Imai, D.; Takeda, R.; Ota, A.; Kawai, E.; Hisada, T.; Emoto, M.; Suzuki, Y.; Okazaki, K. Aerobic Exercise Training with Brisk Walking Increases Intestinal Bacteroides in Healthy Elderly Women. Nutrients 2019, 11, 868. [Google Scholar] [CrossRef] [Green Version]
- Rettedal, E.A.; Cree, J.; Adams, S.E.; MacRae, C.; Skidmore, P.; Cameron-Smith, D.; Gant, N.; Blenkiron, C.; Merry, T.L. Short-term high-intensity interval training exercise does not affect gut bacterial community diversity or composition of lean and overweight men. Exp. Physiol. 2020, 105, 1268–1279. [Google Scholar] [CrossRef]
- Bycura, D.; Santos, A.C.; Shiffer, A.; Kyman, S.; Winfree, K.; Sutliffe, J.; Pearson, T.; Sonderegger, D.; Cope, E.; Caporaso, J.G. Impact of Different Exercise Modalities on the Human Gut Microbiome. Sports 2021, 9, 14. [Google Scholar]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar]
- Mörkl, S.; Lackner, S.; Müller, W.; Gorkiewicz, G.; Kashofer, K.; Oberascher, A.; Painold, A.; Holl, A.; Holzer, P.; Meinitzer, A.; et al. Gut microbiota and body composition in anorexia nervosa inpatients in comparison to athletes, overweight, obese, and normal weight controls. Int. J. Eat. Disord. 2017, 50, 1421–1431. [Google Scholar]
- Yang, Y.; Shi, Y.; Wiklund, P.; Tan, X.; Wu, N.; Zhang, X.; Tikkanen, O.; Zhang, C.; Munukka, E.; Cheng, S. The Association between Cardiorespiratory Fitness and Gut Microbiota Composition in Premenopausal Women. Nutrients 2017, 9, 792. [Google Scholar] [CrossRef]
- Petersen, L.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2018, 67, 625–633. [Google Scholar]
- Durk, R.P.; Castillo, E.; Márquez-Magaña, L.; Grosicki, G.J.; Bolter, N.D.; Lee, C.M.; Bagley, J.R. Gut Microbiota Composition Is Related to Cardiorespiratory Fitness in Healthy Young Adults. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Langsetmo, L.; Johnson, A.; Demmer, R.T.; Fino, N.; Orwoll, E.S.; Ensrud, K.E.; Hoffman, A.R.; Cauley, J.A.; Shmagel, A.; Meyer, K.; et al. The Association between Objectively Measured Physical Activity and the Gut Microbiome among Older Community Dwelling Men. J. Nutr. Health Aging 2019, 23, 538–546. [Google Scholar] [PubMed]
- O’Donovan, C.M.; Madigan, S.M.; Garcia-Perez, I.; Rankin, A.; O’ Sullivan, O.; Cotter, P.D. Distinct microbiome composition and metabolome exists across subgroups of elite Irish athletes. J. Sci. Med. Sport 2020, 23, 63–68. [Google Scholar] [PubMed] [Green Version]
- Castellanos, N.; Diez, G.G.; Antúnez-Almagro, C.; Bailén, M.; Bressa, C.; González Soltero, R.; Pérez, M.; Larrosa, M. A Critical Mutualism—Competition Interplay Underlies the Loss of Microbial Diversity in Sedentary Lifestyle. Front. Microbiol. 2020, 10, 3142. [Google Scholar] [CrossRef]
- Tabone, M.; Bressa, C.; García-Merino, J.A.; Moreno-Pérez, D.; Van, E.C.; Castelli, F.A.; Fenaille, F.; Larrosa, M. The effect of acute moderate-intensity exercise on the serum and fecal metabolomes and the gut microbiota of cross-country endurance athletes. Sci. Rep. 2021, 11, 3558. [Google Scholar]
- Aya, V.; Flórez, A.; Perez, L.; Ramírez, J.D. Association between physical activity and changes in intestinal microbiota composition: A systematic review. PLoS ONE 2021, 16, e0247039. [Google Scholar]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Scheepers, L.E.; Penders, J.; Mbakwa, C.A.; Thijs, C.; Mommers, M.; Arts, I.C. The intestinal microbiota composition and weight development in children: The KOALA Birth Cohort Study. Int. J. Obes. 2015, 39, 16–25. [Google Scholar]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar]
- Samuel, B.S.; Gordon, J.I. A humanized gnotobiotic mouse model of host-archaeal-bacterial mutualism. Proc. Natl. Acad. Sci. USA 2006, 103, 10011–10016. [Google Scholar] [CrossRef] [Green Version]
- Samuel, B.S.; Hansen, E.E.; Manchester, J.K.; Coutinho, P.M.; Henrissat, B.; Fulton, R.; Latreille, P.; Kim, K.; Wilson, R.K.; Gordon, J.I. Genomic and metabolic adaptations of Methanobrevibacter smithii to the human gut. Proc. Natl. Acad. Sci. USA 2007, 104, 10643–10648. [Google Scholar] [CrossRef] [Green Version]
- Merino, E.; Jensen, R.A.; Yanofsky, C. Evolution of bacterial trp operons and their regulation. Curr. Opin. Microbiol. 2008, 11, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, F.; Pasolli, E.; Ercolini, D. Newly Explored Faecalibacterium Diversity Is Connected to Age, Lifestyle, Geography, and Disease. Curr. Biol. 2020, 30, 4932–4943. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Recommendations on Physical Activity for Health; WHO Library Cataloguing-in-Publication Data: Geneva, Switzerland, 2010; ISBN 9789241599979. [Google Scholar]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.C.; LePard, K.J.; Kwak, J.W.; Stancukas, M.C.; Laskowski, S.; Dougherty, J.; Moulton, L.; Glawe, A.; Wang, Y.; Leone, V.; et al. Exercise prevents weight gain and alters the gut microbiota in a mouse model of high fat diet-induced obesity. PLoS ONE 2014, 9, e92193. [Google Scholar]
- Allen, J.M.; Berg Miller, M.E.; Pence, B.D.; Whitlock, K.; Nehra, V.; Gaskins, H.R.; White, B.A.; Fryer, J.D.; Woods, J.A. Voluntary and forced exercise differentially alters the gut microbiome in C57BL/6J mice. J. Appl. Physiol. 2015, 118, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Mahizir, D.; Briffa, J.F.; Wood, J.L.; Anevska, K.; Hill-Yardin, E.L.; Jefferies, A.J.; Gravina, S.; Mazzarino, G.; Franks, A.E.; Moritz, K.M.; et al. Exercise improves metabolic function and alters the microbiome in rats with gestational diabetes. FASEB J. 2020, 34, 1728–1744. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar]
- Shaw, K.A.; Bertha, M.; Hofmekler, T.; Chopra, P.; Vatanen, T.; Srivatsa, A.; Prince, J.; Kumar, A.; Sauer, C.; Zwick, M.E.; et al. Dysbiosis, inflammation, and response to treatment: A longitudinal study of pediatric subjects with newly diagnosed inflammatory bowel disease. Genome Med. 2016, 8, 75. [Google Scholar]
- Myhrstad, M.; Tunsjø, H.; Charnock, C.; Telle-Hansen, V.H. Dietary Fiber, Gut Microbiota, and Metabolic Regulation-Current Status in Human Randomized Trials. Nutrients 2020, 12, 859. [Google Scholar] [CrossRef] [Green Version]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling Interactions in the Microbiome: A Network Perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar]
- Jangi, S.; Gandhi, R.; Cox, L.M.; Li, N.; von Glehn, F.; Yan, R.; Patel, B.; Mazzola, M.A.; Liu, S.; Glanz, B.L.; et al. Alterations of the human gut microbiome in multiple sclerosis. Nat. Commun. 2016, 7, 12015. [Google Scholar]
- Kaakoush, N.O. Sutterella Species, IgA-degrading Bacteria in Ulcerative Colitis. Trends Microbiol. 2020, 28, 519–522. [Google Scholar] [CrossRef]
- Gaike, A.H.; Paul, D.; Bhute, S.; Dhotre, D.P.; Pande, P.; Upadhyaya, S.; Reddy, Y.; Sampath, R.; Ghosh, D.; Chandraprabha, D.; et al. The Gut Microbial Diversity of Newly Diagnosed Diabetics but Not of Prediabetics Is Significantly Different from That of Healthy Nondiabetics. mSystems 2020, 5, e00578-19. [Google Scholar] [CrossRef] [Green Version]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [PubMed] [Green Version]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Gomes Neto, J.C.; Boekschoten, M.V.; Schwab, C.; Urich, T.; et al. Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum schaedleri, a Core Member of the Murine Gut Microbiota. mSystems 2017, 2, e00171-16. [Google Scholar] [PubMed] [Green Version]
- Getachew, B.; Aubee, J.I.; Schottenfeld, R.S.; Csoka, A.B.; Thompson, K.M.; Tizabi, Y. Ketamine interactions with gut-microbiota in rats: Relevance to its antidepressant and anti-inflammatory properties. BMC Microbiol. 2018, 18, 222. [Google Scholar]
- Miquel, S.; Martín, R.; Bridonneau, C.; Robert, V.; Sokol, H.; Bermúdez-Humarán, L.G.; Thomas, M.; Langella, P. Ecology and metabolism of the beneficial intestinal commensal bacterium Faecalibacterium prausnitzii. Gut Microbes 2014, 5, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Meehan, C.J.; Beiko, R.G. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar]
- Säemann, M.D.; Böhmig, G.A.; Osterreicher, C.H.; Burtscher, H.; Parolini, O.; Diakos, C.; Stöckl, J.; Hörl, W.H.; Zlabinger, G.J. Anti-inflammatory effects of sodium butyrate on human monocytes: Potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. 2000, 14, 2380–2382. [Google Scholar] [CrossRef]
- Morishima, S.; Aoi, W.; Kawamura, A.; Kawase, T.; Takagi, T.; Naito, Y.; Tsukahara, T.; Inoue, R. Intensive, prolonged exercise seemingly causes gut dysbiosis in female endurance runners. J. Clin. Biochem. Nutr. 2021, 68, 253–258. [Google Scholar] [CrossRef]
- Roberts, J.D.; Suckling, C.A.; Peedle, G.Y.; Murphy, J.A.; Dawkins, T.G.; Roberts, M.G. An Exploratory Investigation of Endotoxin Levels in Novice Long Distance Triathletes, and the Effects of a Multi-Strain Probiotic/Prebiotic, Antioxidant Intervention. Nutrients 2016, 8, 733. [Google Scholar] [CrossRef] [Green Version]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3kinase signalling. J. Inflamm. 2012, 9, 1. [Google Scholar]
- Yücel, G.; Zhao, Z.; El-Battrawy, I.; Lan, H.; Lang, S.; Li, X.; Akin, I. Lipopolysaccharides induced inflammatory responses and electrophysiological dysfunctions in human-induced pluripotent stem cell derived cardiomyocytes. Sci. Rep. 2017, 7, 2935. [Google Scholar] [PubMed]
- Rajilić-Stojanović, M.; de Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [PubMed]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tomé, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar]
- Tuovinen, E.; Keto, J.; Nikkilä, J.; Mättö, J.; Lähteenmäki, K. Cytokine response of human mononuclear cells induced by intestinal Clostridium species. Anaerobe 2013, 19, 70–76. [Google Scholar]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Paz Soldan, M.M.; Luckey, D.H.; Marietta, E.V.; Jeraldo, P.R.; Chen, X.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci. Rep. 2016, 6, 28484. [Google Scholar]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar]
- Rosenkilde, M.; Petersen, M.B.; Gram, A.S.; Quist, J.S.; Winther, J.; Kamronn, S.D.; Milling, D.H.; Larsen, J.E.; Jespersen, A.P.; Stallknecht, B. The GO-ACTIWE randomized controlled trial—An interdisciplinary study designed to investigate the health effects of active commuting and leisure time physical activity. Contemp. Clin. Trials 2017, 53, 122–129. [Google Scholar]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar]
- Yoon, M.S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar]
- Shou, J.; Chen, P.J.; Xiao, W.H. The Effects of BCAAs on Insulin Resistance in Athletes. J. Nutr. Sci. Vitaminol. 2019, 65, 383–389. [Google Scholar]
- Bosco, N.; Noti, M. The aging gut microbiome and its impact on host immunity. Genes Immun. 2021, 22, 289–303. [Google Scholar]
- Maynard, C.; Weinkove, D. The Gut Microbiota and Ageing. Sub-Cell. Biochem. 2018, 90, 351–371. [Google Scholar]
- Zapata, H.J.; Quagliarello, V.J. The microbiota and microbiome in aging: Potential implications in health and age-related diseases. J. Am. Geriatr. Soc. 2015, 63, 776–781. [Google Scholar] [CrossRef] [Green Version]
- Rondanelli, M.; Giacosa, A.; Faliva, M.A.; Perna, S.; Allieri, F.; Castellazzi, A.M. Review on microbiota and effectiveness of probiotics use in older. World J. Clin. Cases 2015, 3, 156–162. [Google Scholar] [CrossRef]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Martín-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical 69 parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Tims, S.; Derom, C.; Jonkers, D.M.; Vlietinck, R.; Saris, W.H.; Kleerebezem, M.; de Vos, W.M.; Zoetendal, E.G. Microbiota conservation and BMI signatures in adult monozygotic twins. ISME J. 2013, 7, 707–717. [Google Scholar]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar]
- American College of Sports Medicine; Chodzko-Zajko, W.J.; Proctor, D.N.; Fiatarone Singh, M.A.; Minson, C.T.; Nigg, C.R.; Salem, G.J.; Skinner, J.S. American College of Sports Medicine position stand. Exercise and physical activity for older adults. Med. Sci. Sports Exerc. 2009, 41, 1510–1530. [Google Scholar] [PubMed]
- Šoltys, K.; Lendvorský, L.; Hric, I.; Baranovičová, E.; Penesová, A.; Mikula, I.; Bohmer, M.; Budiš, J.; Vávrová, S.; Grones, J.; et al. Strenuous Physical Training, Physical Fitness, Body Composition and Bacteroides to Prevotella Ratio in the Gut of Elderly Athletes. Front. Physiol. 2021, 12, 670989. [Google Scholar] [PubMed]
- Hjorth, M.F.; Christensen, L.; Kjølbæk, L.; Larsen, L.H.; Roager, H.M.; Kiilerich, P.; Kristiansen, K.; Astrup, A. Pretreatment Prevotella-to-Bacteroides ratio and markers of glucose metabolism as prognostic markers for dietary weight loss maintenance. Eur. J. Clin. Nutr. 2020, 74, 338–347. [Google Scholar] [PubMed]
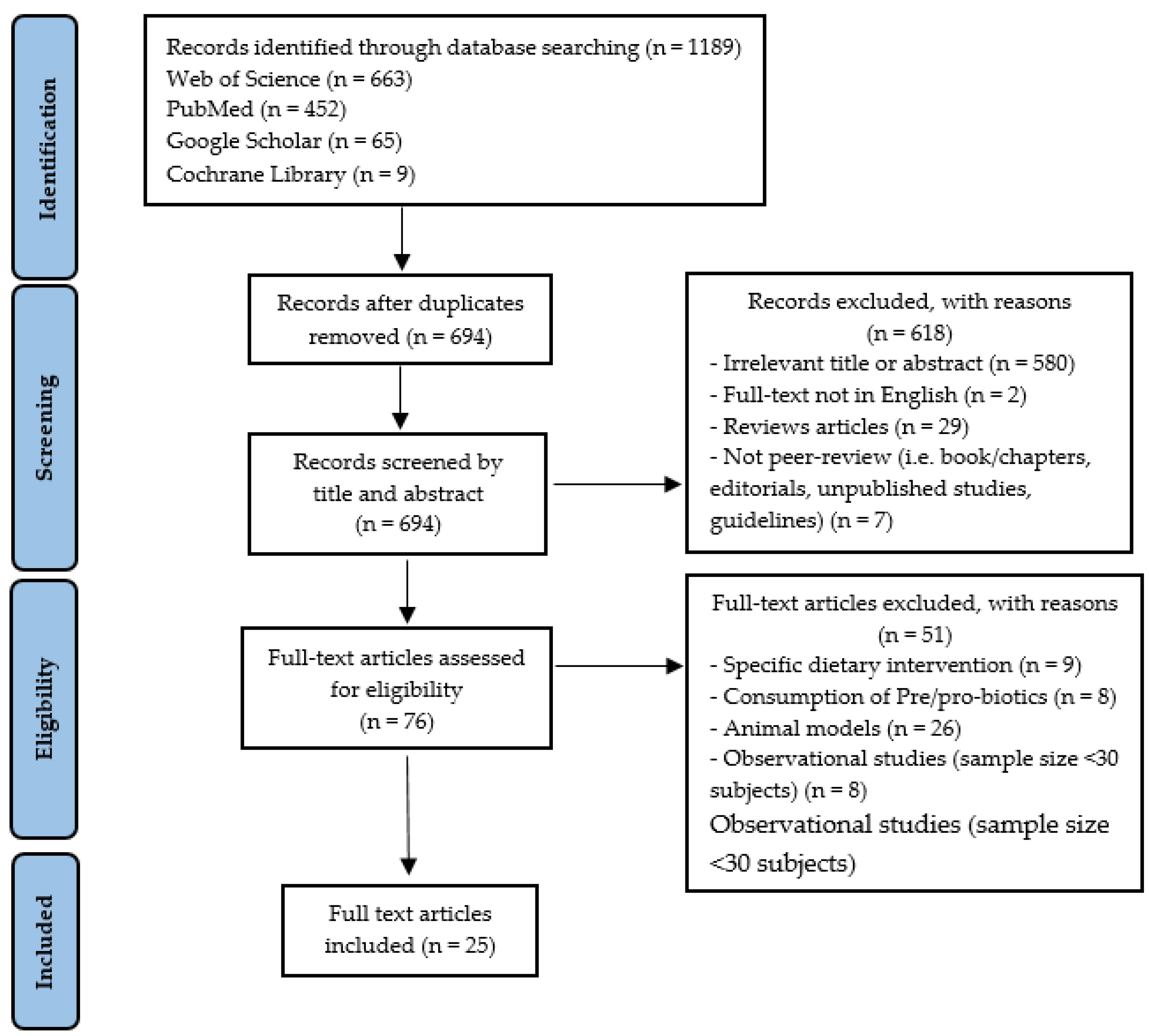

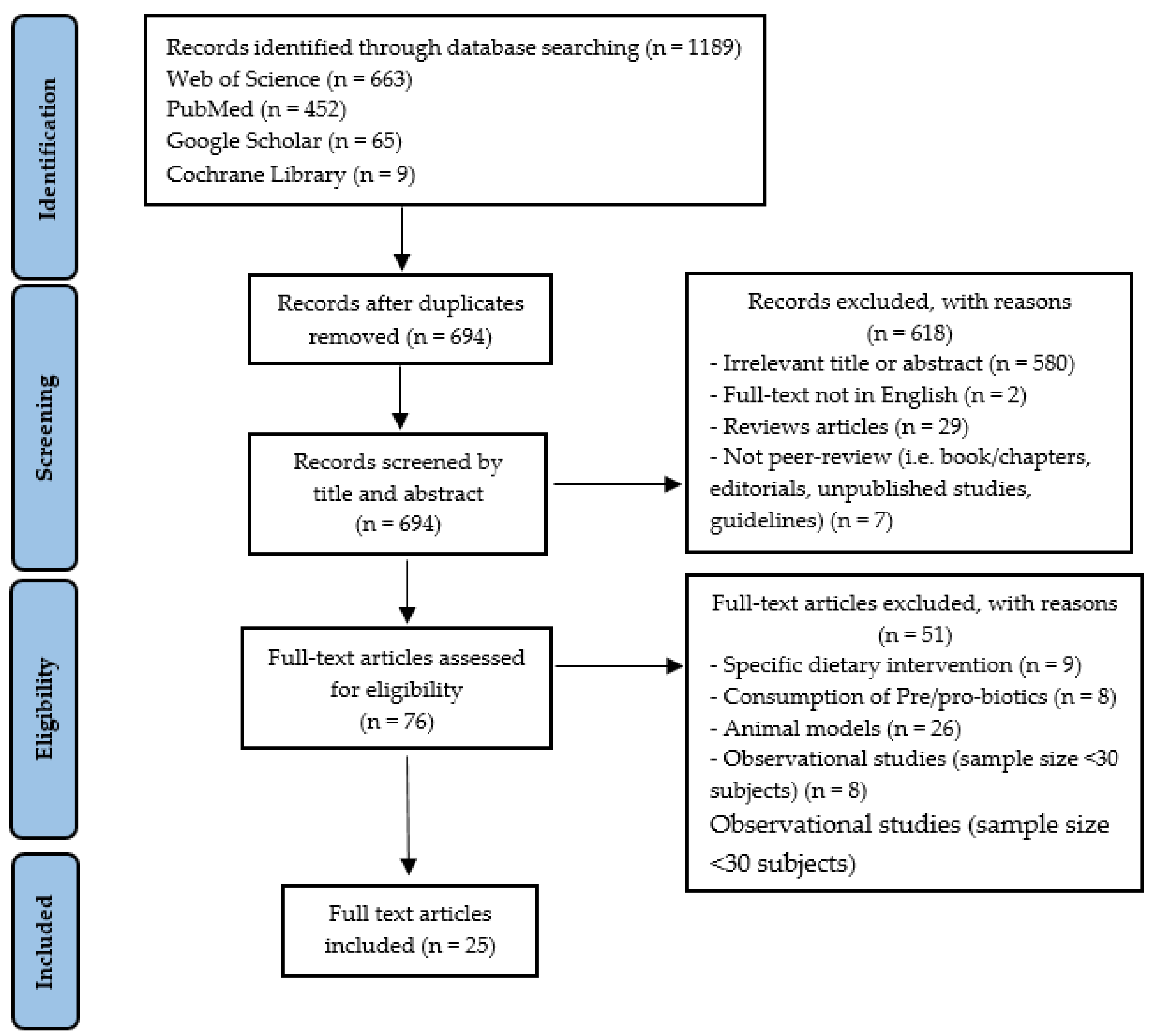{kind=link}
| Inclusion Criteria | Exclusion Criteria | |
|---|---|---|
| Population | Healthy and unhealthy subjects, no age restrictions, both sexes (from sedentary to athlete subjects). | Subjects who take or have taken (in the month before the intervention) pre/pro-biotics and/or antibiotics. |
| Intervention/Exposure | Intervention with any kind of PE protocol or PA/PE exposure. | Intervention with a specific dietary protocol. |
| Comparator | Intervention that has a control group running a different PA/PE protocol or none, a comparison subgroup, or at least a pre/post-intervention comparison. | Absence of any kind of control/comparison. |
| Outcome(s) | Measures of differences for α and β diversity, relative abundance of specific bacteria, metabolomic and metagenomic data analyzed with any kind of sequencing tool. | Lack of baseline and/or follow-up data, or absence of at least one of the measurements indicated in the inclusion criteria—outcome(s). |
| Study Design | Studies with experimental design (randomized and non-randomized trial), observational studies (sample size >30 subjects). | Observational studies (sample size < 30 subjects), case study. |
| Authors | Was the Treatment Randomly Allocated? | Was the Randomization Procedure Described and Was it Appropriate? | Was There a Description of Withdrawals and Dropout? | Was There a Clear Description of the Inclusion/Exclusion Criteria? | Were the Methods of Statistical Analysis Described? | Jadad Score (0–5) |
|---|---|---|---|---|---|---|
| Taniguchi et al. (2018) [24] | Yes | Yes | Yes | Yes | Yes | 5 |
| Kern et al. (2020) [25] | Yes | Yes | No | Yes | Yes | 4 |
| Motiani et al. (2020) [26] | Yes | Yes | Yes | Yes | Yes | 5 |
| Liu et al. (2020) [27] | Yes | Yes | No | Yes | Yes | 4 |
| Quiroga et al. (2020) [28] | Yes | Yes | Yes | Yes | Yes | 5 |
| Resende et al. (2021) [29] | Yes | Yes | Yes | Yes | Yes | 5 |
| Zhong et al. (2021) [30] | Yes | Yes | Yes | Yes | Yes | 5 |
| Moitnho-Silva et al. (2021) [31] | Yes | Yes | Yes | Yes | Yes | 5 |
| Study | Bias Due to Confounding | Bias in Selection of Participants into the Study | Bias in Classification/ Measurement of Intervention | Bias Due to Deviations from Intended Interventions | Bias Because of Missing Data | Bias in Measurement of Outcomes | Bias in Selection of the Reported Result | Overall |
|---|---|---|---|---|---|---|---|---|
| Allen et al., 2018 [14] | Low | Moderate | Moderate | Moderate | Moderate | Low | Low | Moderate |
| Munukka et al., 2018 [32] | Low | Moderate | Low | Moderate | Moderate | Moderate | Moderate | Moderate |
| Morita et al., 2019 [33] | Low | Low | Low | Moderate | Moderate | Moderate | Low | Moderate |
| Rettedal et al., 2020 [34] | Low | Low | Low | Low | Low | Low | Low | Low |
| Bycura et al., 2021 [35] | Low | Low | Low | Moderate | Moderate | Low | Low | Moderate |
| Study | Where the Criteria for Inclusion in the Sample Clearly Defined? | Where the Study Subjects and the Setting Described in Detail? | Was the Exposure Measured in a Valid and Reliable Way? | Were Objective, Standard Criteria Used for Measurement of the Condition? | Were Confounding Factors Identified? | Were Strategies to Deal with Confounding Factors Stated? | Were the Outcomes Measured in a Valid and Reliable Way? | Was Appropriate Statistical Analysis Used? | Overall Appraisal (Included/Excluded) |
|---|---|---|---|---|---|---|---|---|---|
| Clarke et al. (2014) [11] | Y | Y | Y | Y | Y | Y | Y | Y | I |
| Estaki et al. (2016) [12] | Y | Y | Y | Y | Y | Y | Y | Y | I |
| Bressa et al. (2017) [36] | Y | Y | Y | Y | Y | Y | Y | Y | I |
| Mörkl et al. (2017) [37] | Y | Y | Y | Y | Y | N | Y | Y | I |
| Yang et al. (2017) [38] | Y | Y | Y | Y | Y | N | Y | Y | I |
| Petersen et al. (2017) [39] | Y | Y | Y | Y | Y | N | Y | Y | I |
| Barton et al. (2018) [40] | Y | Y | Y | Y | N | N | Y | Y | I |
| Durk et al. (2019) [41] | Y | Y | Y | Y | N | N | Y | Y | I |
| Langsemo et al. (2019) [42] | Y | Y | Y | Y | Y | N | Y | Y | I |
| O’Donovan et al. (2020) [43] | Y | Y | Y | Y | N | N | Y | Y | I |
| Castellanos et al. (2020) [44] | Y | Y | Y | Y | N | N | Y | Y | I |
| Tabone et al. (2021) [45] | Y | Y | Y | Y | Y | Y | Y | Y | I |
| Authors | Study Design | Sample | Subjects Age (years) | Type PA/PE | Protocol/ Workload Assessment | Diet Assessment | Duration Intervention | GM Analysis System | Main Outcomes |
|---|---|---|---|---|---|---|---|---|---|
| Clarke et al., 2014 [11] | Cross-sectional | n = 86 (M) elite professional rugby players (n = 40) (BMI 29.1 ± 2.9), healthy control (n = 46) (23: BMI ≤ 25—23: BMI > 28) | Elite: 29 (±4) Control: 29 (±6) | Rugby PE: aerobic-anaerobic | / | 187-food items FFQ. Macronutrients, fiber, and supplement intake | / | 16S rRNA GA V4 region | Athletes: ↑ α-diversity, ↑ diversity Firmicutes (phylum), ↑ Prevotella, ↓ Bacteroides, ↓ LactobacillusAthletes/Low BMI: ↑ Akkermansia (genus) |
| Estaki et al., 2016 [12] | Cross-sectional | n = 39 (M/F) healthy subjects, stratified by CRF(Low; Average; High) | L: 25.5 (±3.3) A: 24.3 (±3.7) H: 26.2 (±5.5) | PE: aerobic | / | 24 h dietary recall interview. Macronutrients, fiber, saturated fat, and PUFA intake | / | 16S rRNA GA V3/V4 region | VO2peak positively associated with ↑ GM diversity; ↑ CRF = ↑ taxa producers SCFAs. No differences in α and β-diversity |
| Bressa et al. 2017 [36] | Cross-sectional | n = 40 (F) active (ACT) (n = 19) and sedentary (SED) (n = 21) subjects, defined by WHO recommendations | ACT: 30.7(±5.9) SED: 32.2(±8.7) | PE: aerobic | / | 97-food items FFQ. Macronutrients, fiber, and main food intake | / | 16S rRNA GA V3/V4 region | ACT: PA ↑ health-promoting bacteria (F.prausnitzii, R.hominis, A.muciniphila) SED: ↑ Barnesiellaceae, ↑ Turicibacter, ↓ Cropococcus No differences in α/β-diversity and at phylum level between groups. |
| Mörkl et al., 2017 [37] | Cross-sectional | n = 106 (F) Anorexia nervosa (AN) patients (n = 18), normal weight (NW) (n = 26), overweight (OW) (n = 22), obese (O) (n = 20) and athletes (AT) (n = 20) | 24.5 (±4.6) | PE: ball sports PE: aerobic-anaerobic | / | Two 24 h recalls. Macronutrients, fiber, Vit D, and magnesium intake | / | 16S rRNA GA V1/V2 region | ↓ GM α-diversity in obese and AN groups compared to athletes. |
| Yang et al., 2017 [38] | Cross-sectional | n = 71 (F) premenopausal with low (L), moderate (M), high (H) CRF | L: 40.4 (36.9–44.0) M: 39.7 (35.5–43.8) H: 30.6 (25.6–35.6) | PA: aerobic | / | 3-days food records (2 weekdays, 1 weekend day). Macronutrients and total energy intake. | / | 16S rRNA hybridization and DNA-staining | ↓ Bacteroides and ↑ Eubacterium rectale-clostridium coccoides in Low VO2max compared to High VO2max group. |
| Petersen et al., 2017 [39] | Cross-sectional | n = 33 (M/F) professional (n = 22) and amateur (n = 11) level competitive cyclists | 19–49 (Median age 33) | Cycling PE: aerobic-anaerobic | / | Food questionnaire. Macronutrients and alcohol intake. | / | Metagenomic whole-genome shotgun sequencing and RNA sequencing | No significant correlations between taxonomic cluster and professional or amateur level. ↑ Prevotella relative abundance in cyclists training >11 h/week |
| Barton et al., 2018 [40] | Cross-sectional | n = 86 (M) elite professional athletes (n = 40), healthy control (n = 46) (22: BMI ≤ 25.2—24: BMI ≥ 26.5) | Elite: 29 (±4) Control: 29 (±6) | Rugby PE: aerobic-anaerobic | / | 187-food items FFQ. Macronutrients and total energy intake. | / | Genome shotgun sequencing, fecal metabolomics | ↑ Pathways (↑ AA biosynthesis, ↑ carbohydrate metabolism) and ↑ fecal metabolites (microbial produced SCFAs) in athletes |
| Allen et al., 2018 [14] | Longitudinal design | n = 32 (M/F) previously sedentary subjects, lean (n = 18) and obese (n = 14) | Lean: 25.1 (±6.52) Obese: 31.14 (±8.57) | PE: aerobic | 30′ to 60′ 3×wk moderate-to-vigorous intensity (60–75% HRR) exercises | 7-days dietary records, 3-days food menu before each fecal collection. Macronutrient, micronutrient, and total energy intake | 6 weeks | 16S rRNA GA V4 region | No β-diversity differences among groups. ↑ SCFAs producing taxa related to BMI (Faecalibacterium: ↑ lean ↓ obese, Bacteroides: ↓ lean ↑ obese). Changes largely reversed after 6wk of inactivity. |
| Munukka et al., 2018 [32] | Non-randomized trial | n = 17 (F) sedentary subjects BMI > 27.5 kg/m2 | 36.8 (±3.9) | PE: endurance | 40′ to 60′ 3×wk exercises, low to moderate intensity | 3-days food records (2 weekdays and 1 weekend day). Macronutrients, fiber, and total energy intake | 6 weeks | 16S rRNA GA V4 region and metagenomics. | ↑ Akkermansia and ↓ Proteobacteria (exercise-responsive taxa). Changes in GM do not affect systemic metabolites. No differences in α-diversity, slight ↑ β-diversity |
| Taniguchi et al., 2018 [24] | Randomized crossover trial | n = 33 (M) elderly Japanese subjects | 62–76 | PE: endurance | 3xwk ce, 30′ (wk 1/2)—45′ (wk 3/5), with incremental intensity | Self-administered FFQ, semi-weighted 16-days dietary records. Macronutrients and total energy intake. | 5 weeks | 16S rRNA GA V3/V4 region | No differences in α and β-diversity. ↓ C.difficile, ↑ Oscillospira. Minor changes in GM associated with cardiometabolic risk factors. |
| Durk et al., 2019 [38] | Cross-sectional | n = 37 (M/F) healthy subjects | 25.7 (±2.2) | PE: aerobic | / | Instructed to follow their normal diet for 7-days and MyFitnessPal app tracking.Macronutrients, fiber, coffee, alcohol, and total energy intake. | / | 16S rRNA GA | VO2max positively associated to ↑ Firmicutes:Bacteroidetes ratio. No differences in α and β-diversity. |
| Langsetmo et al., 2019 [42] | Cross-sectional | n = 373 (M) community-dwelling older subjects | 84.0 (3.9) | PA: aerobic | / | Not controlled or recorded | / | 16S rRNA GA V4 region | PA not associated with α-diversity, slight association with β-diversity. ↑ Cetobacterium and ↓ Coprobacillus, Adlercreutzia, Eryspelotrichaceae CC-115 in higher step counts subjects. |
| Morita et al., 2019 [33] | Non-randomized comparative trial | n = 32 (F) healthy sedentary elderly subjects, trunk muscle (TM) (n = 14) and aerobic exercise (AE) (n = 18) intervention | 70 (66–75) | PE: aerobic or anaerobic | TM: 1 h weekly resistance training AE: 1 h daily brisk walking ≥3 METs | 138-food and beverage items FFQ. Macronutrients, fiber, saturated fat and total energy intake. | 12 weeks | 16S rRNA GA | ↑ Bacteroides relative abundance only in the AE group. |
| Kern et al., 2020 [25] | Randomized controlled trial | n = 88 (M/F) overweight/obese subjects, moderate intensity (n = 31) (MOD), vigorous intensity (n = 24) (VIG), bicycling (n = 18) (BIKE), control (n = 14) (CON) | 36 (30;41) Median (25th percentile; 75th percentile) | PE: aerobic | MOD: 5×wk LTPA at 50% VO2peak VIG: 5×wk LTPA at 70% VO2peak BIKE: 5×wk active bicycle commuting to and from work (F: 9–15 km/M: 11–17 km daily), self-selected intensity | Food registrations (3 weekdays—1 weekend day), participants were asked to weigh and register intake of food and beverages. Macronutrients, fiber, and total energy itnake. | 6 months | 16S rRNA GA V4 region | β-diversity changed in all groups compared to CON, ↑ α-diversity in VIG compared to CON. Decreased heterogeneity in VIG. No genera changed significantly. |
| O’ Donovan et al., 2020 [43] | Cross-sectional | n = 37 (M/F) elite athletes from 16 different sports stratified by dynamic and/or static components | 27 (±5) | PE: different sports PE: aerobic-anaerobic | / | FFQ. Macronutrients, fiber, beverage, and total energy intake. | / | Metagenomic whole-genome shotgun sequencing and urine and fecal metabolomics | Individual variability among athletes, majority samples driven by 5 species (E. rectale, P. necessaries, F. prausnitzii, B. vulgatus, G. massiliensis). High dynamic component: most compositionally distinct GM. High dynamic+static components: most functionally distinct GM. |
| Motiani et al., 2020 [26] | Randomized controlled trial | n = 26 (M/F) obese sedentary prediabetic/T2D, sprint interval training (n = 13) (SIT), moderate-intensity continuous training (n = 13) (MICT) | 49 (±4) | PE: aerobic | SIT: 3×wk HIIT 30″ exercise bouts (4-6) cycling (wingate protocol) 4′ recovery between bouts MICT: 40′–60′ 3×wk moderate intensity (60% VO2peak) cycling | Not controlled or recorded. Instructed to maintain their dietary habit. | 2 weeks | 16S rRNA GA V3/V4 region | ↑ Bacteroidetes ↓ Firmicutes:Bacteroidetes ratio, ↓ Clostridium and Blautia genus. |
| Catellanos et al., 2020 [44] | Cross-sectional | n = 109 (M/F) healthy subjects, active (n = 64) (ACT) and sedentary (n = 45) (SED), described by WHO recommendations | ACT: 32.17 (±7.40) SED: 33.69 (±7.96) | PE: aerobic | / | 93-food items FFQ. Macronutrients, fiber, ethanol, and total energy intake. | / | 16S rRNA GA V3/V4 region | GM network of active people has higher efficiency and transmissibility rate. Key bacteria reorganization from ACT to SED: Roseburia faecis, unclassified roseburia spp. Key bacteria reorganization from SED to ACT: unclassified Sutterella spp. |
| Liu et al., 2020 [27] | Randomized controlled trial | n = 39 (M) medication naïve overweight/obese pre-diabetic subjects | Responders: 43.29 (±3.27) Non-responders:(36.00 ± 4.55) | PE: aerobic and anaerobic | 70′ 3×wk high intensity combined aerobic and resistance interval training, 80–95% HRmax | FFQ. Macronutrients, fiber, and total energy intake. | 12 weeks | Metagenomic whole genome shotgun sequencing and fecal metabolomics | Exercises-induced alterations in the GM correlated with improvement in glucose homeostasis and insulin sensitivity. GM responders: ↑ biosynthesis SCFAs, ↑ BCAA catabolism, ↓ Bacteroides, ↑ Streptococcus mitisGM non-responders: ↑ production of detrimental compounds. No differences in α and β-diversity. Functional capacity of GM can be altered without major shifts in its community structure. |
| Quiroga et al., 2020 [28] | Randomized controlled trial | n = 39 obese pediatric children (n = 25) and healthy control (n = 14) | 7–12 | PE: Endurance plus strength | 2×wk combined endurance (sprint of 30” max cadence at 3′30″, 4′30″, 5′30″, and 6′30″) and strength training (30–70% 1RM) | Nutritional advice for a healthy and balanced diet. | 12 weeks | 16S rRNA GA V3/V4 region | ↓ Proteobacteria phylum and Gammaproteobacteria class, ↑ Blautia, Dialister and Roseburia genera lead to a GM profile like that of healthy children. |
| Rettedal et al., 2020 [34] | Non-randomized trial | n = 29 (M) overweight (n = 15) and lean (n = 14) subjects | Overweight: 31 (±2) Lean: 29 (±2) | PE: aerobic | 3×wk ce HIIT, 60″ cycling intervals at VO2peak workload interspersed with 75″ rest, 8 to 12 intervals | FFQ for baseline intake. Instructed to maintain normal dietary pattern. Macronutrients, fiber, saturated fat, PUFA, and total energy intake. | 3 weeks | 16S rRNA GA V3/V4 region | No differences in α and β-diversity. Significant association between the abundance of bacterial spp. (Coprococcus_3, Blautia, Lachnospiraceae_ge, Dorea) and insulin sensitivity marker in the overweight group. |
| Bycura et al., 2021 [35] | Non-randomized trial | n = 56 (M/F) healthy students, cardiorespiratory exercise (n = 28) (CRE), resistance exercise (n = 28) (RTE) | CRE: 20.54 (1.93) RTE: 21.28 (3.85) | PE: aerobic or anaerobic | CRE: 1 h, 3×wk (2-days group cycling, 1-day rotating CRE activity) 60–90% HRmax RTE: 1 h 3×wk full/lower/upper body at 70–85% 1RM | Not controlled or recorded. Instructed to maintain their typical dietary practice and report major deviations. | 8 weeks | 16S rRNA GA V4 region | CRE: initial changes to GM (wk 2,3) not sustained through or after the intervention. RTE: no changes in microbiome composition. |
| Resende et al., 2021 [29] | Randomized controlled trial | n = 22 (M) healthy previously sedentary subjects, exercise (n = 12) and control (n = 12) | Exercise: 25.58 (±5.07) Control: 25.5 (±4.66) | PE: aerobic | 50′ 3×wk ce at steady speed 60 rpm (wk 1,2), 60/65% VO2peak (wk 3–10, weekly progressiveoverload) | Wk 1: food records Wk 5: 3-day food records Wk 12: 48 h diet record before data collection. Macronutrients, fiber, cholesterol, water, and total energy intake. | 10 weeks | 16S rRNA GA V4 region | No differences in α- and β-diversity. No significant changes at phylum, class, order, family, or species level. VO2peak positively associated with α-diversity and to the relative abundance of Roseburia, Odoribacter, and Sutterella. BMI positively associated with Desulfovibrio and Faecalibacterium genera. |
| Tabone et al., 2021 [43] | Cross-sectional | n = 40 (M) endurance cross-country runners | 35.79 (±8.01) | PE: endurance | / | FFQ, 24 h dietary recall (2 weekdays, 1 weekend day). Macronutrients and total energy intake. | / | 16S rRNA GA V3/V4 region | 85 serum and 12 fecal metabolites and 6 bacterial taxa (Romboutsia, Escherichia coli TOP498, Ruminococcaceae UCG-005, Blautia, Ruminiclostridium 9 and Clostridium phoceensis) were modified. Crosstalk between GM and systemic tryptophan metabolism. |
| Zhong et al., 2021 [30] | Randomized controlled trial | n = 12 (F) previously inactive older healthy subjects, exercise (n = 6) and control (n = 6) | Exercise: 69.83 (±4.50) Control: 67.50 (±4.28) | PE: aerobic and anaerobic | 1 h 4 × wk combined aerobic and resistance exercises (progressive overload) | Not controlled or recorded | 8 weeks | 16S rRNA GA V4F/V4R region | No changes in α-diversity. ↑ Prevotella, ↑ Verrucomicrobia, ↓ Proteobacteria abundance in the exercise group. |
| Moitinho-Silva et al., 2021 [31] | Randomized controlled trial | n = 36 (M/F) healthy physical inactive subjects, endurance (n = 12) and strength exercises (n = 13) with control (n = 11). Elite athletes for comparison (n = 13) | Endurance: 31.4 (±8.3) Strength: 29.9 (±7.9) Control: 33.4 (±7.9) Elite: 30 (±9.9) | PE:aerobic or anaerobic | Endurance: 30′ (at least) 3 × wk running Strength: 30′ 3 × wk whole-body hypertrophy strength training | Food questionnaire Elite: no data. Macronutrients, fiber, and total energy intake. | 6 weeks | 16S rRNA GA V1/V2 region | No specific bacteria changes. GM change patterns largely varied among individuals of the same group. No differences in α-diversity between elite and physical inactive subjects. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cataldi, S.; Bonavolontà, V.; Poli, L.; Clemente, F.M.; De Candia, M.; Carvutto, R.; Silva, A.F.; Badicu, G.; Greco, G.; Fischetti, F. The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology 2022, 11, 479. https://doi.org/10.3390/biology11030479
Cataldi S, Bonavolontà V, Poli L, Clemente FM, De Candia M, Carvutto R, Silva AF, Badicu G, Greco G, Fischetti F. The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology. 2022; 11(3):479. https://doi.org/10.3390/biology11030479
Chicago/Turabian StyleCataldi, Stefania, Valerio Bonavolontà, Luca Poli, Filipe Manuel Clemente, Michele De Candia, Roberto Carvutto, Ana Filipa Silva, Georgian Badicu, Gianpiero Greco, and Francesco Fischetti. 2022. "The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review" Biology 11, no. 3: 479. https://doi.org/10.3390/biology11030479
APA StyleCataldi, S., Bonavolontà, V., Poli, L., Clemente, F. M., De Candia, M., Carvutto, R., Silva, A. F., Badicu, G., Greco, G., & Fischetti, F. (2022). The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology, 11(3), 479. https://doi.org/10.3390/biology11030479