
Interleukin-6-Production Is Responsible for Induction of Hepatic Synthesis of Several Chemokines as Acute-Phase Mediators in Two Animal Models: Possible Significance for Interpretation of Laboratory Changes in Severely Ill Patients



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Induction of APR and Harvesting Blood and Liver Tissue
2.2. Enzyme-Linked Immunosorbent Assay (ELISA) for IL6 and Chemokines
2.3. RNA Isolation and Real-Time-PCR
2.4. Protein Isolation and Western Blot Analysis
2.5. Immunohistochemical Study
3. Results
3.1. Measurement of Serum Cytokine and Chemokines Level
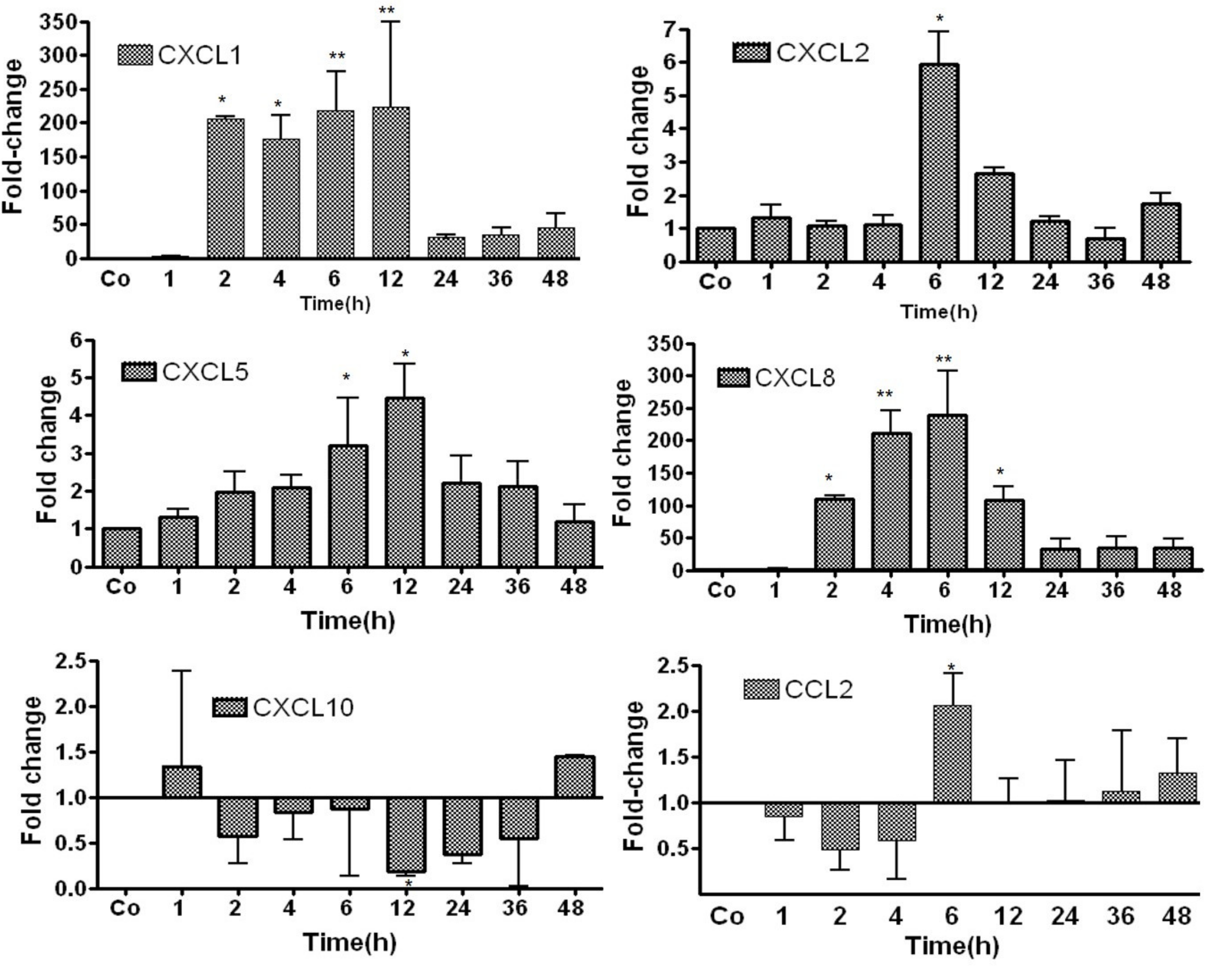
3.2. Study of Gene Expression of Chemokines in TO-Treated Rat Liver by RT-PCR
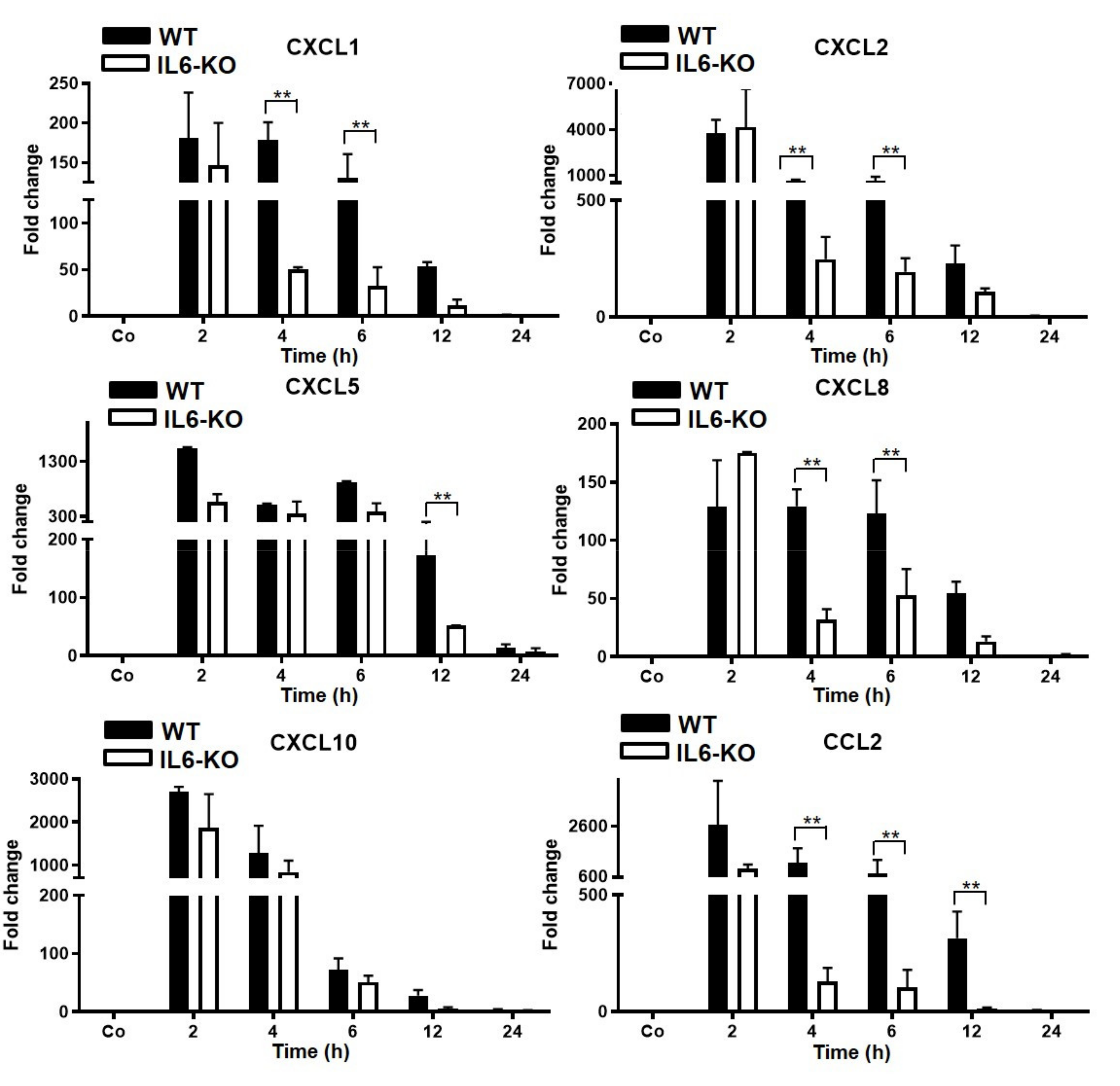
3.3. Changes in Gene Expression Levels of Chemokine in Wild-Type (WT) and IL6-Knock-Out (KO) in TO-Injected Mice
- A.
- Chemokine-genes are constitutively expressed in liver tissue at different magnitudes;
- B.
- Changes in the expression of various chemokines in the liver can be divided into three groups:
- (a)
- IL6-strongly upregulated (up to 150-fold) chemokines: CXCL1 and CXCL8 (independent of the constitutive amount of their mRNA);
- (b)
- IL6-moderately upregulated chemokines: CXCL5 and CXCL2;
- (c)
- IL6-unaffected chemokines: CXCL10 and CCL2.
3.4. Changes in Hepatic Gene Expression Level of Chemokines in WT and IL6-KO LPS-Injected Mice (Intraperitoneal)
3.5. Detection and Change in Chemokines and STAT-3-Protein Levels in Liver Tissue of WT and IL6-KO Mice after Intramuscular TO or Intraperitoneal LPS Administration
3.6. Immunohistochemical Detection of Neutrophil Granulocytes in the Rat Liver after Intramuscular TO-Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramadori, G.; Christ, B. Cytokines and the Hepatic Acute-Phase Response. Semin. Liver Dis. 1999, 19, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, F.; Patzelt, J.; Langer, H.F. The Platelet Response to Tissue Injury. Front. Med. 2018, 5, 317. [Google Scholar] [CrossRef]
- Ramadori, G.; Moriconi, F.; Malik, I.; Dudas, J. Physiology and pathophysiology of liver inflammation, damage and repair. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2008, 59, 107–117. [Google Scholar]
- Moshage, H. Cytokines and the hepatic acute phase response. J. Pathol. 1997, 181, 257–266. [Google Scholar] [CrossRef]
- Gruys, E.; Toussaint, M.J.M.; Niewold, T.A.; Koopmans, S.J. Acute phase reaction and acute phase proteins. J. Zhejiang Univ. Sci. B 2005, 6, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Whicher, J.T.; Chambers, R.E.; Higginson, J.; Nashef, L.; Higgins, P.G. Acute phase response of serum amyloid A protein and C reactive protein to the common cold and influenza. J. Clin. Pathol. 1985, 38, 312–316. [Google Scholar] [CrossRef]
- Nemeth, E.; Valore, E.V.; Territo, M.; Schiller, G.; Lichtenstein, A.; Ganz, T. Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003, 101, 2461–2463. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, G.; Sial, G.Z.K.; Ramadori, P.; Dudas, J.; Batusic, D.S.; Ramadori, G. Changes of hepatic lactoferrin gene expression in two mouse models of the acute phase reaction. Int. J. Biochem. Cell Biol. 2011, 43, 1822–1832. [Google Scholar] [CrossRef]
- Sultan, S.; Pascucci, M.; Ahmad, S.; Malik, I.A.; Bianchi, A.; Ramadori, P.; Ahmad, G.; Ramadori, G. LIPOCALIN-2 Is a Major Acute-Phase Protein in a Rat and Mouse Model of Sterile Abscess. Shock 2012, 37, 191–196. [Google Scholar] [CrossRef]
- Chi, Y.; Ge, Y.; Wu, B.; Zhang, W.; Wu, T.; Wen, T.; Liu, J.; Guo, X.; Huang, C.; Jiao, Y.; et al. Serum Cytokine and Chemokine Profile in Relation to the Severity of Coronavirus Disease 2019 in China. J. Infect. Dis. 2020, 222, 746–754. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kox, M.; Waalders, N.J.B.; Kooistra, E.J.; Gerretsen, J.; Pickkers, P. Cytokine Levels in Critically Ill Patients with COVID-19 and Other Conditions. JAMA 2020, 324, 1565–1567. [Google Scholar] [CrossRef]
- Jafarzadeh, A.; Chauhan, P.; Saha, B.; Jafarzadeh, S.; Nemati, M. Contribution of monocytes and macrophages to the local tissue inflammation and cytokine storm in COVID-19: Lessons from SARS and MERS, and potential therapeutic interventions. Life Sci. 2020, 257, 118102. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, M.; Chen, X.; Montaner, L.J. Cytokine storm and leukocyte changes in mild versus severe SARS-CoV-2 infection: Review of 3939 COVID-19 patients in China and emerging pathogenesis and therapy concepts. J. Leukoc. Biol. 2020, 108, 17–41. [Google Scholar] [CrossRef]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Hue, S.; Beldi-Ferchiou, A.; Bendib, I.; Surenaud, M.; Fourati, S.; Frapard, T.; Rivoal, S.; Razazi, K.; Carteaux, G.; Delfau-Larue, M.-H.; et al. Uncontrolled Innate and Impaired Adaptive Immune Responses in Patients with COVID-19 Acute Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 2020, 202, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. Chemokines: A New Classification System and Their Role in Immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef]
- Bone-Larson, C.L.; Simpson, K.J.; Colletti, L.M.; Lukacs, N.W.; Chen, S.-C.; Lira, S.; Kunkel, S.L.; Hogaboam, C.M. The role of chemokines in the immunopathology of the liver. Immunol. Rev. 2000, 177, 8–20. [Google Scholar] [CrossRef]
- Karlmark, K.R.; Wasmuth, H.E.; Trautwein, C.; Tacke, F. Chemokine-directed immune cell infiltration in acute and chronic liver disease. Expert Rev. Gastroenterol. Hepatol. 2008, 2, 233–242. [Google Scholar] [CrossRef]
- Malik, I.A.; Moriconi, F.; Sheikh, N.; Naz, N.; Khan, S.; Dudas, J.; Mansuroglu, T.; Hess, C.F.; Rave-Fränk, M.; Christiansen, H.; et al. Single-Dose Gamma-Irradiation Induces Up-Regulation of Chemokine Gene Expression and Recruitment of Granulocytes into the Portal Area but Not into Other Regions of Rat Hepatic Tissue. Am. J. Pathol. 2010, 176, 1801–1815. [Google Scholar] [CrossRef]
- Amanzada, A.; Moriconi, F.; Mansuroglu, T.; Cameron, S.; Ramadori, G.; Malik, I.A. Induction of chemokines and cytokines before neutrophils and macrophage recruitment in different regions of rat liver after TAA administration. Lab. Investig. 2013, 94, 235–247. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, K.; Henderson, R.B.; Laschinger, M.; Hogg, N. Neutrophil Chemokines KC and Macrophage-Inflammatory Protein-2 Are Newly Synthesized by Tissue Macrophages Using Distinct TLR Signaling Pathways. J. Immunol. 2008, 180, 4308–4315. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, N.; Tron, K.; Dudas, J.; Ramadori, G. Cytokine-induced neutrophil chemoattractant-1 is released by the noninjured liver in a rat acute-phase model. Lab. Investig. 2006, 86, 800–814. [Google Scholar] [CrossRef]
- Ramadori, P.; Ahmad, G.; Ramadori, G. Cellular and molecular mechanisms regulating the hepatic erythropoietin expression during acute-phase response: A role for IL-6. Lab. Investig. 2010, 90, 1306–1324. [Google Scholar] [CrossRef] [PubMed]
- Malik, G.; Wilting, J.; Hess, C.F.; Ramadori, G.; Malik, I.A. PECAM -1 modulates liver damage induced by synergistic effects of TNF-α and irradiation. J. Cell. Mol. Med. 2019, 23, 3336–3344. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.; Colaço, H.G.; Neves-Costa, A.; Seixas, E.; Velho, T.R.; Pedroso, D.; Barros, A.; Martins, R.; Carvalho, N.; Payen, D.; et al. CXCL5-mediated recruitment of neutrophils into the peritoneal cavity of Gdf15-deficient mice protects against abdominal sepsis. Proc. Natl. Acad. Sci. USA 2020, 117, 12281–12287. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G. Hypoalbuminemia: An Underestimated, Vital Characteristic of Hospitalized COVID-19 Positive Patients? Hepatoma Res. 2020, 6, 28. [Google Scholar] [CrossRef]
- Baruch, Y.; Neubauer, K.; Ritzel, A.; Wilfling, T.; Lorf, T.; Ramadori, G. Von Willebrand gene expression in damaged human liver. Hepatogastroenterology 2004, 51, 684–688. [Google Scholar]
- Hummel, M.; Czerlinski, S.; Friedel, N.; Liebenthal, C.; Hasper, D.; von Baehr, R.; Hetzer, R.; Volk, H.-D. Interleukin-6 and interleukin-8 concentrations as predictors of outcome in ventricular assist device patients before heart transplantation. Crit. Care Med. 1994, 22, 448–454. [Google Scholar] [CrossRef]
- Patzwahl, R.; Meier, V.; Ramadori, G.; Mihm, S. Enhanced Expression of Interferon-Regulated Genes in the Liver of Patients with Chronic Hepatitis C Virus Infection: Detection by Suppression-Subtractive Hybridization. J. Virol. 2001, 75, 1332–1338. [Google Scholar] [CrossRef]
- Mihm, S.; Schweyer, S.; Ramadori, G. Expression of the chemokine IP-10 correlates with the accumulation of hepatic IFN-? and IL-18 mRNA in chronic hepatitis C but not in hepatitis B. J. Med Virol. 2003, 70, 562–570. [Google Scholar] [CrossRef]
- Schett, G.; Neurath, M.F. Resolution of chronic inflammatory disease: Universal and tissue-specific concepts. Nat. Commun. 2018, 9, 3261. [Google Scholar] [CrossRef]
- Neubauer, K.; Lindhorst, A.; Tron, K.; Ramadori, G.; Saile, B. Decrease of PECAM-1-gene-expression induced by proinflammatory cytokines IFN-γ and IFN-α is reversed by TGF-β in sinusoidal endothelial cells and hepatic mononuclear phagocytes. BMC Physiol. 2008, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.; Biensø, R.S.; Meinertz, S.; Van Hauen, L.; Rasmussen, S.M.; Gliemann, L.; Plomgaard, P.; Pilegaard, H. Impact of training status on LPS-induced acute inflammation in humans. J. Appl. Physiol. 2015, 118, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Petrone, L.; Bondet, V.; Vanini, V.; Cuzzi, G.; Palmieri, F.; Palucci, I.; Delogu, G.; Ciccosanti, F.; Fimia, G.M.; Blauenfeldt, T.; et al. First description of agonist and antagonist IP-10 in urine of patients with active TB. Int. J. Infect. Dis. 2019, 78, 15–21. [Google Scholar] [CrossRef]
- Vazirinejad, R.; Ahmadi, Z.; Kazemi Arababadi, M.; Hassanshahi, G.; Kennedy, D. The Biological Functions, Structure and Sources of CXCL10 and Its Outstanding Part in the Pathophysiology of Multiple Sclerosis. Neuroimmunomodulation 2014, 21, 322–330. [Google Scholar] [CrossRef]
- Dufour, J.H.; Dziejman, M.; Liu, M.T.; Leung, J.H.; Lane, T.E.; Luster, A.D. IFN-γ-Inducible Protein 10 (IP-10; CXCL10)-Deficient Mice Reveal a Role for IP-10 in Effector T Cell Generation and Trafficking. J. Immunol. 2002, 168, 3195–3204. [Google Scholar] [CrossRef]
- Neville, L.F.; Mathiak, G.; Bagasra, O. The immunobiology of interferon-gamma inducible protein 10 kD (IP-10): A novel, pleiotropic member of the C-X-C chemokine superfamily. Cytokine Growth Factor Rev. 1997, 8, 207–219. [Google Scholar] [CrossRef]
- Nabavizadeh, N.; Mitin, T.; Dawson, L.; Hong, T.S.; Thomas, C.R. Stereotactic body radiotherapy for patients with hepatocellular carcinoma and intermediate grade cirrhosis. Lancet Oncol. 2017, 18, e192. [Google Scholar] [CrossRef][Green Version]
- Pastor, L.; Casellas, A.; Rupérez, M.; Carrillo, J.; Maculuve, S.; Jairoce, C.; Paredes, R.; Blanco, J.; Naniche, D. Interferon-γ–Inducible Protein 10 (IP-10) as a Screening Tool to Optimize Human Immunodeficiency Virus RNA Monitoring in Resource-Limited Settings. Clin. Infect. Dis. 2017, 65, 1670–1675. [Google Scholar] [CrossRef] [PubMed]
- Hayney, M.S.; Henriquez, K.M.; Barnet, J.H.; Ewers, T.; Champion, H.M.; Flannery, S.; Barrett, B. Serum IFN-γ-induced protein 10 (IP-10) as a biomarker for severity of acute respiratory infection in healthy adults. J. Clin. Virol. 2017, 90, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Vaninov, N. In the eye of the COVID-19 cytokine storm. Nat. Rev. Immunol. 2020, 20, 277. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, C.; Li, J.; Yuan, J.; Wei, J.; Huang, F.; Wang, F.; Li, G.; Li, Y.; Xing, L.; et al. Plasma IP-10 and MCP-3 levels are highly associated with disease severity and predict the progression of COVID-19. J. Allergy Clin. Immunol. 2020, 146, 119–127.e4. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.-C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y. The severe COVID-19: A sepsis induced by viral infection? And its immunomodulatory therapy. Chin. J. Traumatol. 2020, 23, 190–195. [Google Scholar] [CrossRef]
- Joynt, G.M.; Wu, W.K. Understanding COVID-19: What does viral RNA load really mean? Lancet Infect. Dis. 2020, 20, 635–636. [Google Scholar] [CrossRef]
- Yu, F.; Yan, L.; Wang, N.; Yang, S.; Wang, L.; Tang, Y.; Gao, G.; Wang, S.; Ma, C.; Xie, R.; et al. Quantitative Detection and Viral Load Analysis of SARS-CoV-2 in Infected Patients. Clin. Infect. Dis. 2020, 71, 793–798. [Google Scholar] [CrossRef]
- Lescure, F.-X.; Bouadma, L.; Nguyen, D.; Parisey, M.; Wicky, P.-H.; Behillil, S.; Gaymard, A.; Bouscambert-Duchamp, M.; Donati, F.; Le Hingrat, Q.; et al. Clinical and virological data of the first cases of COVID-19 in Europe: A case series. Lancet Infect. Dis. 2020, 20, 697–706, Correction in Lancet Infect. Dis. 2020, 20, e148. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Sang, L.; Ye, F.; Ruan, S.; Zhong, B.; Song, T.; Alshukairi, A.N.; Chen, R.; Zhang, Z.; et al. Kinetics of viral load and antibody response in relation to COVID-19 severity. J. Clin. Investig. 2020, 130, 5235–5244. [Google Scholar] [CrossRef]
- Pujadas, E.; Chaudhry, F.; McBride, R.; Richter, F.; Zhao, S.; Wajnberg, A.; Nadkarni, G.; Glicksberg, B.S.; Houldsworth, J.; Cordon-Cardo, C. SARS-CoV-2 viral load predicts COVID-19 mortality. Lancet Respir. Med. 2020, 8, e70. [Google Scholar] [CrossRef]
- Beasley, M.B. Acute lung injury—From cannabis to COVID. Mod. Pathol. 2022, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Konopka, K.E.; Nguyen, T.; Jentzen, J.M.; Rayes, O.; Schmidt, C.J.; Wilson, A.M.; Farver, C.F.; Myers, J.L. Diffuse alveolar damage (DAD) resulting from coronavirus disease 2019 Infection is Morphologically Indistinguishable from Other Causes of DAD. Histopathology 2020, 77, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Hopfer, H.; Herzig, M.C.; Gosert, R.; Menter, T.; Hench, J.; Tzankov, A.; Hirsch, H.H.; Miller, S.E. Hunting coronavirus by transmission electron microscopy—A guide to SARS-CoV-2-associated ultrastructural pathology in COVID-19 tissues. Histopathology 2020, 78, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C. Acute Respiratory Distress Syndrome as an Organ Phenotype of Vascular Microthrombotic Disease: Based on Hemostatic Theory and Endothelial Molecular Pathogenesis. Clin. Appl. Thromb. 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G.; Sipe, J.D.; Colten, H.R. Expression and regulation of the murine serum amyloid A (SAA) gene in extrahepatic sites. J. Immunol. 1985, 135, 3645–3647. [Google Scholar]
- Salem, F.; Li, X.Z.; Hindi, J.; Casablanca, N.M.; Zhong, F.; El Jamal, S.M.; Al Rasheed, M.R.H.; Li, L.; Lee, K.; Chan, L.; et al. Activation of STAT3 signaling pathway in the kidney of COVID-19 patients. J. Nephrol. 2021, 9, 1–9. [Google Scholar] [CrossRef]
- Can, F.K.; Özkurt, Z.; Öztürk, N.; Sezen, S. Effect of IL-6, IL-8/CXCL8, IP-10/CXCL 10 levels on the severity in COVID 19 infection. Int. J. Clin. Pr. 2021, 75, e14970. [Google Scholar] [CrossRef]
- Nikolich-Zugich, J.; Knox, K.S.; Rios, C.T.; Natt, B.; Bhattacharya, D.; Fain, M.J. SARS-CoV-2 and COVID-19 in older adults: What we may expect regarding pathogenesis, immune responses, and outcomes. GeroScience 2020, 42, 505–514. [Google Scholar] [CrossRef]
- Moreno-Guerrero, S.; Ramírez-Pacheco, A.; Rocha-Ramírez, L.; Hernández-Pliego, G.; Eguía-Aguilar, P.; Escobar-Sánchez, M.; Reyes-López, A.; Juárez-Villegas, L.; Sienra-Monge, J. Association of Genetic Polymorphisms and Serum Levels of IL-6 and IL-8 with the Prognosis in Children with Neuroblastoma. Cancers 2021, 13, 529. [Google Scholar] [CrossRef]
- Lu, Y.-R.; Rao, Y.-B.; Mou, Y.-J.; Chen, Y.; Lou, H.-F.; Zhang, Y.; Zhang, D.-X.; Xie, H.-Y.; Hu, L.-W.; Fang, P. High concentrations of serum interleukin-6 and interleukin-8 in patients with bipolar disorder. Medicine 2019, 98, e14419. [Google Scholar] [CrossRef]
- Lane, D.; Matte, I.; Rancourt, C.; Piché, A. Prognostic significance of IL-6 and IL-8 ascites levels in ovarian cancer patients. BMC Cancer 2011, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Cameron, S.; Blaschke, M.; Moriconi, F.; Naz, N.; Amanzada, A.; Ramadori, G.; Malik, I.A. Differential gene expression of chemokines in KRAS and BRAF mutated colorectal cell lines: Role of cytokines. World J. Gastroenterol. 2014, 20, 2979–2994. [Google Scholar] [CrossRef] [PubMed]
- Della-Torre, E.; Criscuolo, E.; Lanzillotta, M.; Locatelli, M.; Clementi, N.; Mancini, N.; Dagna, L.; COVID-BioB Study Group. IL-1 and IL-6 inhibition affects the neutralising activity of anti-SARS-CoV-2 antibodies in patients with COVID-19. Lancet Rheumatol. 2021, 3, e829–e831. [Google Scholar] [CrossRef]
- Nicola, M.; O’Neill, N.; Sohrabi, C.; Khan, M.; Agha, M.; Agha, R. Evidence based management guideline for the COVID-19 pandemic—Review article. Int. J. Surg. 2020, 77, 206–216. [Google Scholar] [CrossRef]
- Violi, F.; Ceccarelli, G.; Loffredo, L.; Alessandri, F.; Cipollone, F.; D’Ardes, D.; D’Ettorre, G.; Pignatelli, P.; Venditti, M.; Mastroianni, C.M.; et al. Albumin Supplementation Dampens Hypercoagulability in COVID-19: A Preliminary Report. Thromb. Haemost. 2020, 121, 102–105. [Google Scholar] [CrossRef]
- Neil, D.; Moran, L.; Horsfield, C.; Curtis, E.; Swann, O.; Barclay, W.; Hanley, B.; Hollinshead, M.; Roufosse, C. Ultrastructure of cell trafficking pathways and coronavirus: How to recognise the wolf amongst the sheep. J. Pathol. 2020, 252, 346–357. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | 5 → 3 Forward | 5 → 3 Reverse |
|---|---|---|
| (A) Rat primers | ||
| CXCL1/Kc | GGCAGGGATTCACTTCAAGA | GCCATCGGTGCAATCTATCT |
| CXCL2/Mip2 | ATCCAGAGCTTGACGGTGAC | AGGTACGATCCAGGCTTCCT |
| CXCL5/Lix | CTCAAGCTGCTCCTTTCTCG | GCGATCATTTTGGGGTTAAT |
| CXCL8/lL8 | CCCCCATGGTTCAGAAGATTG | TTGTCAGAAGCCAGCGTTCAC |
| CXCL10/Ip10 | CTGTCGTTCTCTGCCTCGTG | GGATCCCTTGAGTCCCACTCA |
| CCL2/Mcp1 | AGGCAGATGCAGTTAATGCCC | ACACCTGCTGCTGGTGATTCTC |
| β-actin | ACCACCATGTACCCAGGCATT | CCACACAGAGTACTTGCGCTCA |
| Ubc | CACCAAGAAGGTCAAACAGGAA | AAGACACCTCCCCATCAAACC |
| (B) Mouse primers | ||
| CXCL1/Kc | GGATTCACCTCAAGAACATCCAGAG | CACCCTTCTACTAGCACAGTGGTTG |
| CXCL2/Mip2 | CTCTCAAGGGCGGTCAAAAAGTT | TCAGACAGCGAGGCACATCAGGTA |
| CXCL5/Lix | GGTCCACAGTGCCCTACG | GCGAGTGCATTCCGCTTA |
| CXCL8/IL8 | GCTGGGATTCACCTCAAGAA | CTTTTGGACAATTTTCTGAACCA |
| CXCL10/Ip10 | AAGTGCTGCCGTCATTTTCT | GTGGCAATGATCTCAACACG |
| CCl2/Mcp1 | CCCACTCACCTGCTGCTACT | TCTGGACCCATTCCTTCTTG |
| β-actin | ATTGTTACCAACTGGGACGACATG | CGAAGTCTAGAGCAACATAGCACA |
| GAPDH | AGAACATCATCCCTGCATCC | CACATTGGGGGTAGGAACAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, I.A.; Ramadori, G. Interleukin-6-Production Is Responsible for Induction of Hepatic Synthesis of Several Chemokines as Acute-Phase Mediators in Two Animal Models: Possible Significance for Interpretation of Laboratory Changes in Severely Ill Patients. Biology 2022, 11, 470. https://doi.org/10.3390/biology11030470
Malik IA, Ramadori G. Interleukin-6-Production Is Responsible for Induction of Hepatic Synthesis of Several Chemokines as Acute-Phase Mediators in Two Animal Models: Possible Significance for Interpretation of Laboratory Changes in Severely Ill Patients. Biology. 2022; 11(3):470. https://doi.org/10.3390/biology11030470
Chicago/Turabian StyleMalik, Ihtzaz Ahmed, and Giuliano Ramadori. 2022. "Interleukin-6-Production Is Responsible for Induction of Hepatic Synthesis of Several Chemokines as Acute-Phase Mediators in Two Animal Models: Possible Significance for Interpretation of Laboratory Changes in Severely Ill Patients" Biology 11, no. 3: 470. https://doi.org/10.3390/biology11030470
APA StyleMalik, I. A., & Ramadori, G. (2022). Interleukin-6-Production Is Responsible for Induction of Hepatic Synthesis of Several Chemokines as Acute-Phase Mediators in Two Animal Models: Possible Significance for Interpretation of Laboratory Changes in Severely Ill Patients. Biology, 11(3), 470. https://doi.org/10.3390/biology11030470