
The VIP/VPAC1R Pathway Regulates Energy and Glucose Homeostasis by Modulating GLP-1, Glucagon, Leptin and PYY Levels in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Experimental Design
2.3. Body Phenotype Analysis
2.4. Determination of Food Intake Behavior
2.5. Assessment of Indirect Calorimetry Using the Promethion Metabolic System
2.6. Plasma Samples and Measurement of Metabolic Hormone Levels
2.7. Insulin Tolerance and Glucose Tolerance Test
2.8. Data Analysis and Trial Exclusion
3. Results
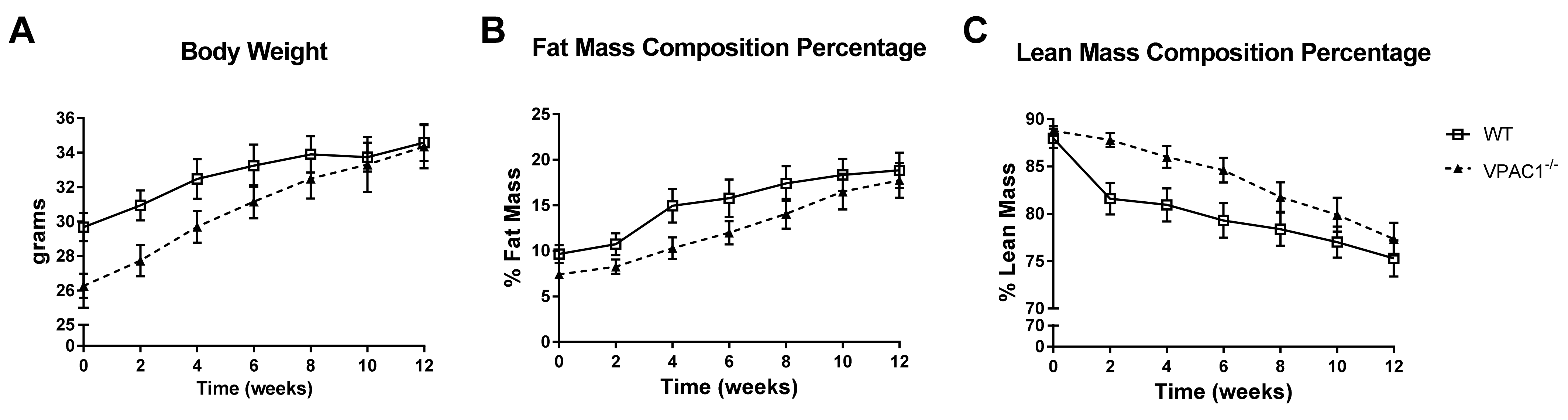
3.1. Body Phenotype Long-Term Study in VPAC1−/− Mice
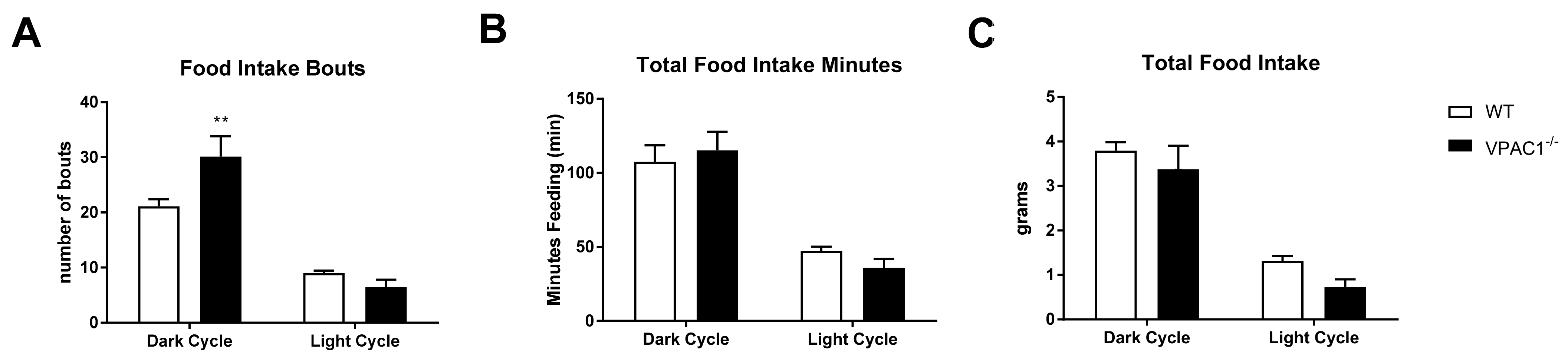
3.2. Feeding Behavior and Food Intake in VPAC1−/− Mice
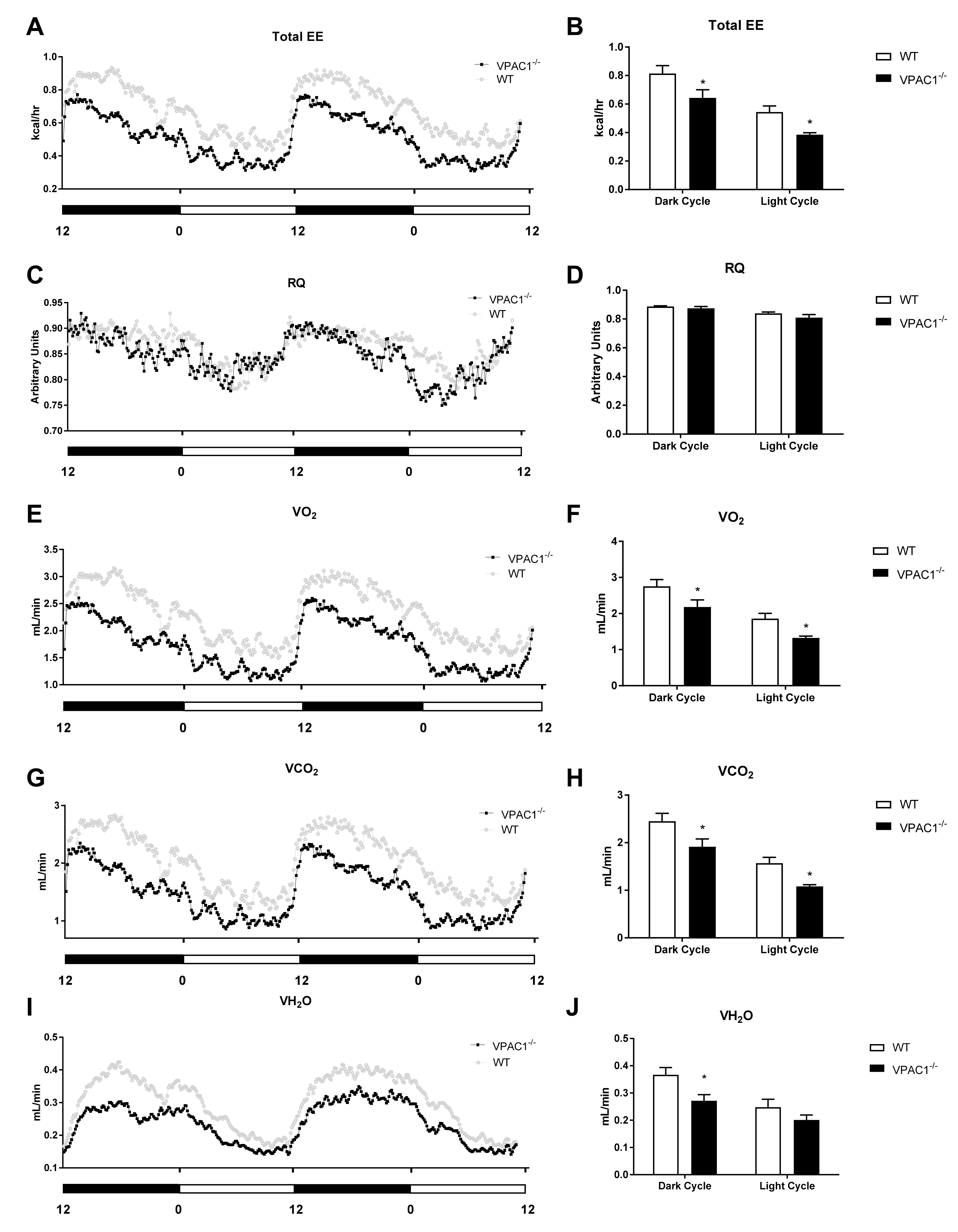
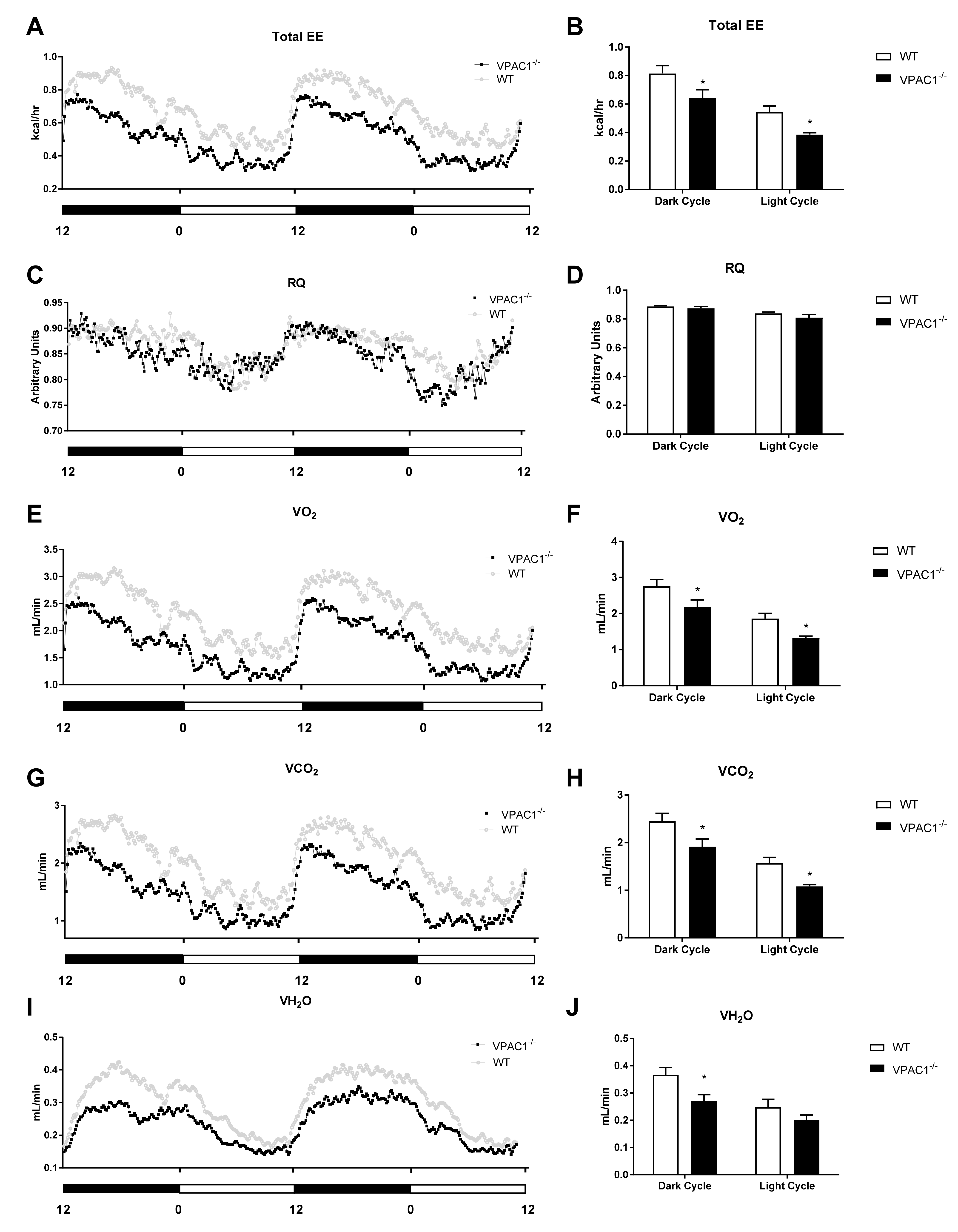
3.3. Calorimetric Parameters Alteration in VPAC1−/− Mice
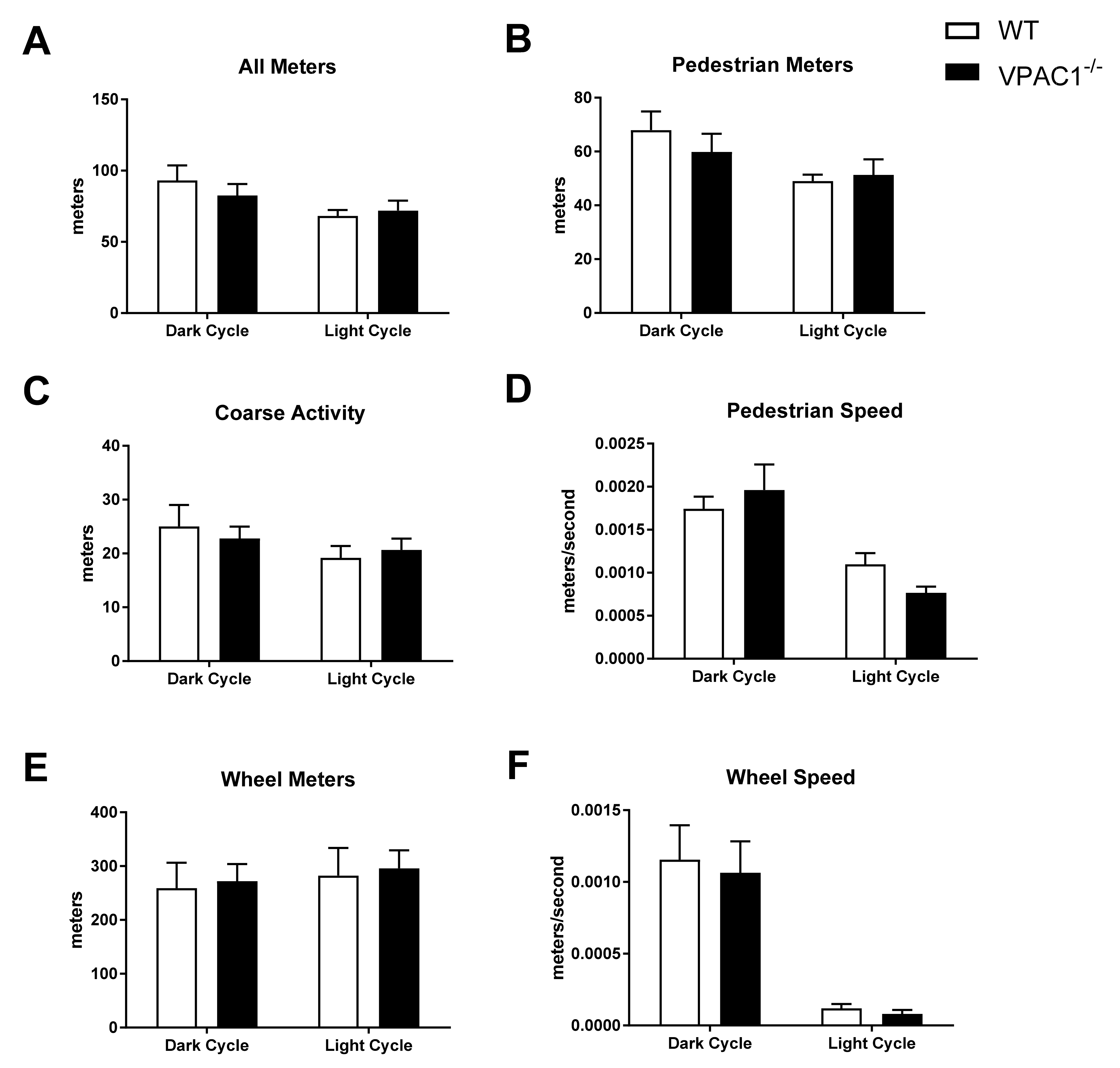
3.4. Physical Activity Values in VPAC1−/− Mice
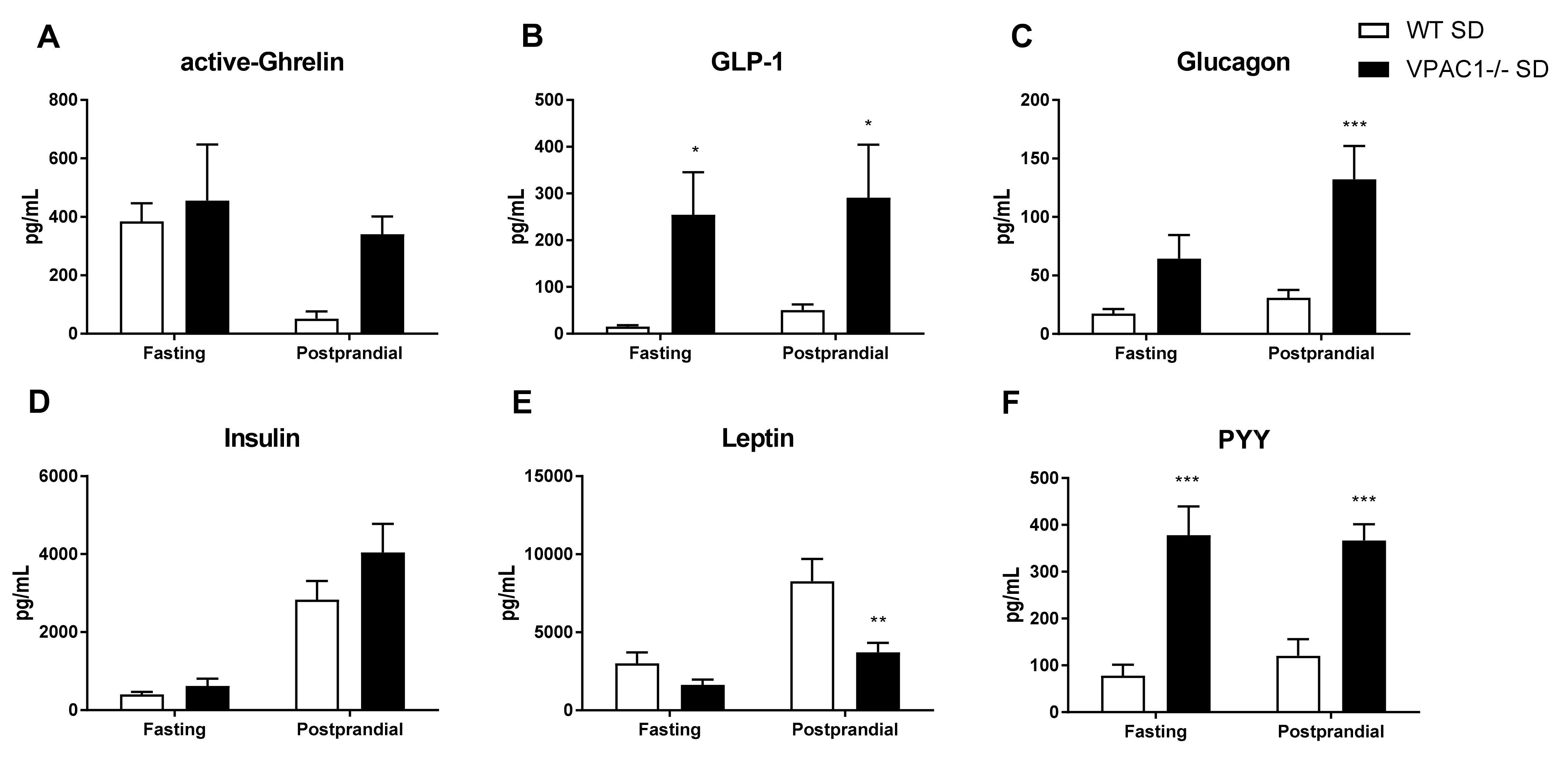
3.5. Circulating Levels of Metabolic Hormones Are Dysregulated in VPAC1−/− Mice
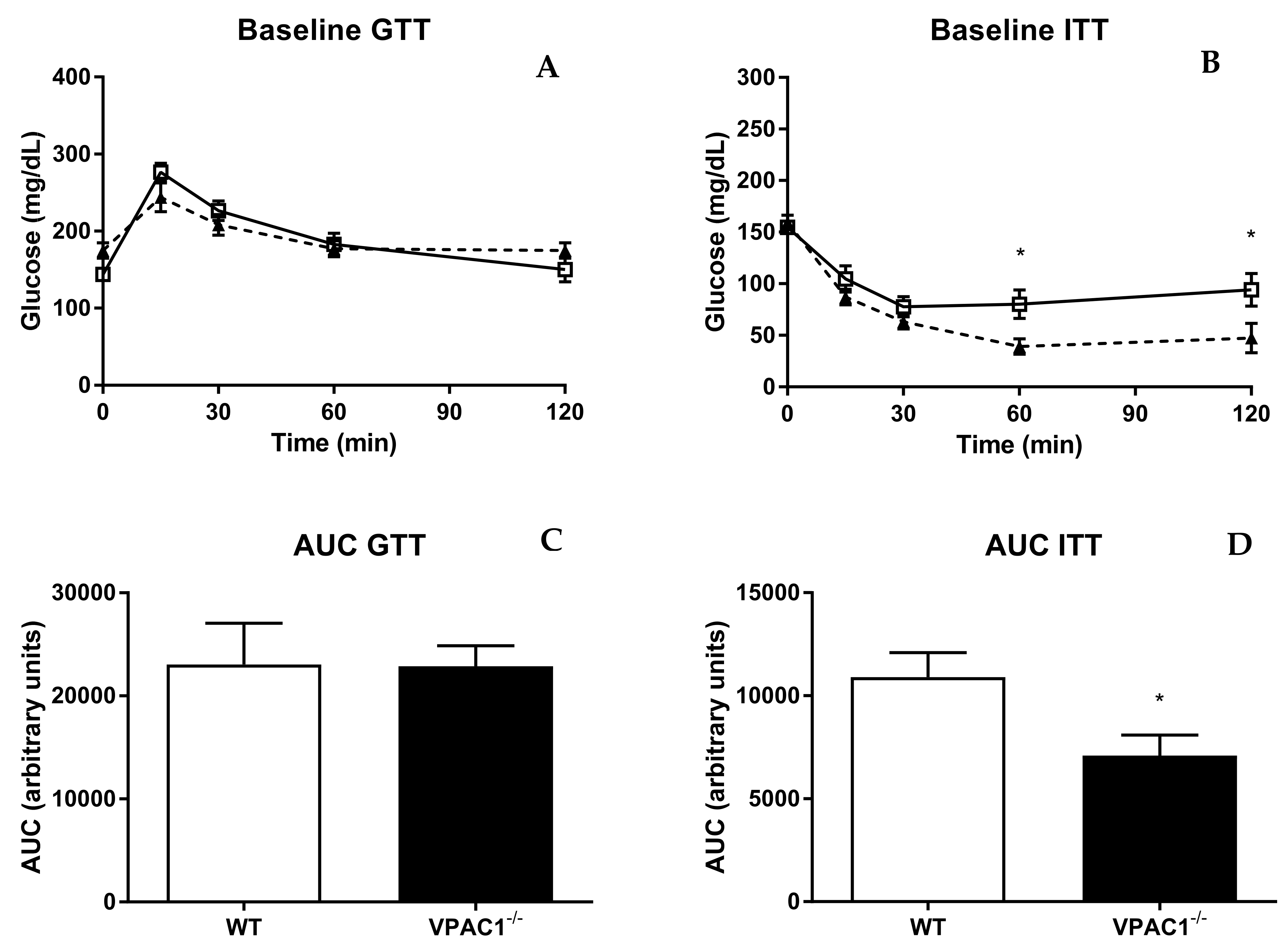
3.6. VPAC1−/− Mice Have Altered Insulin Sensitivity and Tolerance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CNS | central nervous system |
| GI | gastrointestinal |
| GLP-1 | glucagon-like peptide 1 |
| IGF | insulin-like growth factor |
| PAC1-R | pituitary adenylate cyclase activating polypeptide type 1 receptor |
| PACAP | pituitary adenylate cyclase activating polypeptide |
| PCR | polymerase chain reaction |
| PYY | peptide YY |
| SEM | standard error of the mean |
| VIP | vasoactive intestinal polypeptide |
| VPAC1R | vasoactive intestinal polypeptide receptor 1 |
| VPAC2R | vasoactive intestinal polypeptide receptor 2 |
| BW | body weight |
| qPCR | quantitative polymerase chain reaction |
| T2D | type 2 diabetes |
| TEE | Total Energy Expenditure |
| VCO2 | carbon dioxide production |
| VO2 | rate of oxygen consumption |
| VH2O | water vapor |
| RQ | respiratory quotient |
| GPCRs | G-protein coupled receptors |
| WT | wild type |
| GTT | glucose tolerance test |
| ITT | insulin tolerance test |
References
- Alhabeeb, H.; AlFaiz, A.; Kutbi, E.; AlShahrani, D.; Alsuhail, A.; AlRajhi, S.; Alotaibi, N.; Alotaibi, K.; AlAmri, S.; Alghamdi, S.; et al. Gut Hormones in Health and Obesity: The Upcoming Role of Short Chain Fatty Acids. Nutrients 2021, 13, 481. [Google Scholar] [CrossRef] [PubMed]
- Marić, G.; Gazibara, T.; Zaletel, I.; Borović, M.L.; Tomanović, N.; Cirić, M.; Puškaš, N. The Role of Gut Hormones in Appetite Regulation (Review). Acta Physiol. Hung. 2014, 101, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Vu, J.P.; Goyal, D.; Luong, L.; Oh, S.; Sandhu, R.; Norris, J.; Parsons, W.; Pisegna, J.R.; Germano, P.M. PACAP intraperitoneal treatment suppresses appetite and food intake via PAC1 receptor in mice by inhibiting ghrelin and increasing GLP-1 and leptin. Am. J. Physiol. Liver Physiol. 2015, 309, G816–G825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sureshkumar, K.; Saenz, A.; Ahmad, S.M.; Lutfy, K. The PACAP/PAC1 Receptor System and Feeding. Brain Sci. 2021, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Sanford, D.; Luong, L.; Gabalski, A.; Oh, S.; Vu, J.P.; Pisegna, J.R.; Germano, P.M. An Intraperitoneal Treatment with Calcitonin Gene-Related Peptide (CGRP) Regulates Appetite, Energy Intake/Expenditure, and Metabolism. J. Mol. Neurosci. 2019, 67, 28–37. [Google Scholar] [CrossRef]
- Moody, T.W.; Ito, T.; Osefo, N.; Jensen, R.T. VIP and PACAP: Recent insights into their functions/roles in physiology and disease from molecular and genetic studies. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, M.; Akiba, Y.; Kaunitz, J.D. Recent advances in VIP physiology and pathophysiology: Focus on the gastrointestinal system. F1000Research 2019, 8, 1629. [Google Scholar] [CrossRef] [Green Version]
- Bains, M.; Laney, C.; Wolfe, A.E.; Orr, M.; Waschek, J.A.; Ericsson, A.C.; Glenn, P.D. VIP is associated with altered gut microbiota communities in male and female C57Bl/6 mice. Front. Microbiol. 2019, 10, 2689. [Google Scholar] [CrossRef] [Green Version]
- Talbot, J.; Hahn, P.; Kroehling, L.; Nguyen, H.; Li, D.; Littman, D.R. Feeding-dependent VIP neuron-ILC3 circuit regulates the intestinal barrier. Nature 2020, 579, 575–580. [Google Scholar] [CrossRef]
- Jia, T.; Zhao, N. Expression and function of vasoactive intestinal peptide receptors in human lower esophageal sphincter. Ann. Palliat. Med. 2021, 10, 3067–3077. [Google Scholar] [CrossRef]
- Conlin, V.S.; Wu, X.; Nguyen, C.; Dai, C.; Vallance, B.A.; Buchan, A.M.J.; Boyer, L.; Jacobson, K. Vasoactive intestinal peptide ameliorates intestinal barrier disruption associated with Citrobacter rodentium-induced colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G735–G750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Conlin, V.S.; Morampudi, V.; Ryz, N.R.; Nasser, Y.; Bhinder, G.; Bergstrom, K.S.; Yu, H.B.; Waterhouse, C.C.M.; Buchan, A.; et al. Vasoactive Intestinal Polypeptide Promotes Intestinal Barrier Homeostasis and Protection Against Colitis in Mice. PLoS ONE 2015, 10, e0125225. [Google Scholar]
- Voice, J.K.; Dorsam, G.; Chan, R.C.; Grinninger, C.; Kong, Y.; Goetzl, E.J. Immunoeffector and immunoregulatory activities of VIP. Regul. Pept. 2002, 109, 199–208. [Google Scholar] [CrossRef]
- Delgado, M.; Ganea, D. Vasoactive intestinal peptide: A neuropeptide with pleiotropic immune functions. Amino Acids 2013, 45, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Seillet, C.; Luong, K.; Tellier, J.; Jacquelot, N.; Shen, R.D.; Hickey, P.; Wimmer, V.C.; Whitehead, L.; Rogers, K.L.; Smyth, G.K.; et al. The neuropeptide VIP confers anticipatory mucosal immunity by regulating ILC3 activity. Nat. Immunol. 2020, 21, 168–177. [Google Scholar] [CrossRef]
- Vu, J.P.; Larauche, M.; Flores, M.; Luong, L.; Norris, J.; Oh, S.; Liang, L.J.; Waschek, J.; Pisegna, J.R.; Germano, P.M. Regulation of Appetite, Body Composition, and Metabolic Hormones by Vasoactive Intestinal Polypeptide (VIP). J. Mol. Neurosci. 2015, 56, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Harmar, A.J.; Fahrenkrug, J.; Gozes, I.; Laburthe, M.; May, V.; Pisegna, J.R.; Vaudry, D.; Vaudry, H.; Waschek, J.A.; Said, S.I. Pharmacology and functions of receptors for vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide: IUPHAR review 1. Br. J. Pharmacol. 2012, 166, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Couvineau, A.; Ceraudo, E.; Tan, Y.V.; Nicole, P.; Laburthe, M. The VPAC1 receptor: Structure and function of a class B GPCR prototype. Front. Endocrinol. 2012, 3, 139. [Google Scholar] [CrossRef] [Green Version]
- Ceraudo, E.; Hierso, R.; Tan, Y.V.; Murail, S.; Rouyer-Fessard, C.; Nicole, P.; Robert, J.C.; Jamin, N.; Neumann, J.M.; Robberecht, P.; et al. Spatial proximity between the VPAC1 receptor and the amino terminus of agonist and antagonist peptides reveals distinct sites of interaction. FASEB J. 2012, 26, 2060–2071. [Google Scholar] [CrossRef]
- Langer, I.; Jeandriens, J.; Couvineau, A.; Sanmukh, S.; Latek, D. Signal Transduction by VIP and PACAP Receptors. Biomedicines 2022, 10, 406. [Google Scholar] [CrossRef]
- Jayawardena, D.; Guzman, G.; Gill, R.K.; Alrefai, W.A.; Onyuksel, H.; Dudeja, P.K. Expression and localization of VPAC1, the major receptor of vasoactive intestinal peptide along the length of the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G16–G25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, S.R.; Bryant, M.G. Proceedings: The distribution of vasoactive intestinal peptide (VIP) in the primate gastrointestinal tract and characterization of VIP from human tumors. Gut 1973, 14, 823. [Google Scholar] [PubMed]
- Ishihara, T.; Shigemoto, R.; Mori, K.; Takahashi, K.; Nagata, S. Functional expression and tissue distribution of a novel receptor for vasoactive intestinal polypeptide. Neuron 1992, 8, 811–819. [Google Scholar] [CrossRef]
- Wei, Y.; Mojsov, S. Tissue specific expression of different human receptor types for pituitary adenylate cyclase activating polypeptide and vasoactive intestinal polypeptide: Implications for their role in human physiology. J. Neuroendocrinol. 1996, 8, 811–817. [Google Scholar] [CrossRef]
- Usdin, T.B.; Bonner, T.I.; Mezey, E. Two receptors for vasoactive intestinal polypeptide with similar specificity and complementary distributions. Endocrinology 1994, 135, 2662–2680. [Google Scholar] [CrossRef]
- Reubi, J.C. In vitro evaluation of VIP/PACAP receptors in healthy and diseased human tissues. Clinical implications. Ann. N. Y. Acad. Sci. 2000, 921, 1–25. [Google Scholar] [CrossRef]
- Fung, C.; Unterweger, P.; Parry, L.J.; Bornstein, J.C.; Foong, J.P. VPAC1 receptors regulate intestinal secretion and muscle contractility by activating cholinergic neurons in guinea pig jejunum. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G748–G758. [Google Scholar] [CrossRef]
- Fabricius, D.; Karacay, B.; Shutt, D.; Leverich, W.; Schafer, B.; Takle, E.; Thedens, D.; Khanna, G.; Raikwar, S.; Yang, B.; et al. Characterization of intestinal and pancreatic dysfunction in VPAC1-null mutant mouse. Pancreas 2011, 40, 861–871. [Google Scholar] [CrossRef]
- Vu, J.P.; Luong, L.; Parsons, W.F.; Oh, S.; Sanford, D.; Gabalski, A.; Lighton, J.R.; Pisegna, J.R.; Germano, P.M. Long-Term Intake of a High-Protein Diet Affects Body Phenotype, Metabolism, and Plasma Hormones in Mice. J. Nutr. 2017, 147, 2243–2251. [Google Scholar] [CrossRef] [Green Version]
- Lighton, J.R.B. Measuring Metabolic Rates: A Manual for Scientists, 1st ed.; Oxford University Press: New York, NY, USA, 2008. [Google Scholar]
- Weir, J.B.d.V. New methods for calculating metabolic rate with special reference to protein metabolism. J. Physiol. 1949, 109, 1–9. [Google Scholar] [CrossRef]
- Tschoep, M.; Speakman, J.R.; Arch, J.R.S.; Auwerx, J.; Brüning, J.C.; Chan, L.; Eckel, R.H.; Farese, R.V., Jr.; Galgani, J.E.; Hambly, C.; et al. A guide to analysis of mouse energy metabolism. Nat. Methods 2011, 9, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asnicar, M.A.; Köster, A.; Heiman, M.L.; Tinsley, F.; Smith, D.P.; Galbreath, E.; Fox, N.; Ma, Y.L.; Blum, W.F.; Hsiung, H.M. Vasoactive intestinal polypeptide/pituitary adenylate cyclase-activating peptide receptor 2 deficiency in mice results in growth retardation and increased basal metabolic rate. Endocrinology 2002, 143, 3994–4006. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.J.; Zhang, L.; Yi, T.H.; Xie, S.S.; Dai, Y. In vivo anti-obesity effect of the agonist for receptor VPAC1. Sheng Li Xue Bao 2008, 60, 751–758. [Google Scholar] [PubMed]
- Lijnen, H.R.; Freson, K.; Hoylaerts, M.F. Effect of VPAC1 Blockade on Adipose Tissue Formation and Composition in Mouse Models of Nutritionally Induced Obesity. J. Obes. 2010, 2010, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Akesson, L.; Ahrén, B.; Edgren, G.; Degerman, E. VPAC2-R mediates the lipolytic effects of pituitary adenylate cyclase-activating polypeptide/vasoactive intestinal polypeptide in primary rat adipocytes. Endocrinology 2005, 146, 744–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, L.D.; Evans, K.; Sander, L.D. A possible involvement of VIP in feeding-induced secretion of ACTH and corticosterone in the rat. Physiol. Behav. 1995, 58, 409–413. [Google Scholar] [CrossRef]
- Bechtold, D.A.; Brown, T.M.; Luckman, S.M.; Piggins, H.D. Metabolic rhythm abnormalities in mice lacking VIP-VPAC2 signaling. Am. J. Physiol Regul. Integr. Comp. Physiol. 2008, 294, R344–R351. [Google Scholar] [CrossRef] [Green Version]
- Sheward, W.J.; Maywood, E.S.; French, K.L.; Horn, J.M.; Hastings, M.H.; Seckl, J.R.; Holmes, M.C.; Harmar, A.J. Entrainment to feeding but not to light: Circadian phenotype of VPAC2 receptor-null mice. J. Neurosci. 2007, 27, 4351–4358. [Google Scholar] [CrossRef]
- Martin, B.; Shin, Y.K.; White, C.M.; Ji, S.; Kim, W.; Carlson, O.D.; Napora, J.K.; Chadwick, W.; Chapter, M.; Waschek, J.A.; et al. Vasoactive intestinal peptide-null mice demonstrate enhanced sweet taste preference, dysglycemia, and reduced taste bud leptin receptor expression. Diabetes 2010, 59, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Sawmiller, D.R.; Henning, R.J.; Cuevas, J.; Dehaven, W.I.; Vesely, D.L. Coronary vascular effects of vasoactive intestinal peptide in the isolated perfused rat heart. Neuropeptides 2004, 38, 289–297. [Google Scholar] [CrossRef]
- Sawmiller, D.R.; Ashtari, M.; Urueta, H.; Leschinsky, M.; Henning, R.J. Mechanisms of vasoactive intestinal peptide-elicited coronary vasodilation in the isolated perfused rat heart. Neuropeptides 2006, 40, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Fahrenkrug, J.; Georg, B.; Hannibal, J.; Jørgensen, H.L. Role of light and the circadian clock in the rhythmic oscillation of intraocular pressure: Studies in VPAC2 receptor and PACAP deficient mice. Exp. Eye Res. 2018, 169, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Ono, D.; Honma, K.I.; Honma, S. Roles of Neuropeptides, VIP and AVP, in the Mammalian Central Circadian Clock. Front. Neurosci. 2021, 15, 650154. [Google Scholar] [CrossRef]
- Green, C.B.; Takahashi, J.S.; Bass, J. The meter of metabolism. Cell 2008, 134, 728–742. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.; Zak, D.E.; Sauter, T.; Schwaber, J.; Ogunnaike, B.A. Modeling the VPAC2-activated cAMP/PKA signaling pathway: From receptor to circadian clock gene induction. Biophys. J. 2006, 90, 1560–1571. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.T.; Croft, C.L.; Samuels, R.E.; Myung, J.; Takumi, T.; Piggins, H.D. Constant light enhances synchrony among circadian clock cells and promotes behavioral rhythms in VPAC2-signaling deficient mice. Sci. Rep. 2015, 5, 14044. [Google Scholar] [CrossRef] [Green Version]
- Ajpru, S.; McArthur, A.J.; Piggins, H.D.; Sugden, D. Identification of PAC1 receptor isoform mRNAs by real-time PCR in rat suprachiasmatic nucleus. Mol. Brain Res. 2002, 105, 29–37. [Google Scholar] [CrossRef]
- Date, Y.; Kojima, M.; Hosoda, H.; Sawaguchi, A.; Mondal, M.S.; Suganuma, T.; Matsukura, S.; Kangawa, K.; Nakazato, M. Ghrelin, a novel growth hormone-releasing acylated peptide, is synthesized in a distinct endocrine cell type in the gastrointestinal tracts of rats and humans. Endocrinology 2000, 141, 4255–4261. [Google Scholar] [CrossRef]
- Alamri, B.N.; Shin, K.; Chappe, V.; Anini, Y. The role of ghrelin in the regulation of glucose homeostasis. Horm. Mol. Biol. Clin. Investig. 2016, 26, 3–11. [Google Scholar] [CrossRef]
- Müller, T.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, P.; Chepurny, O.G.; Holz, G.G. Regulation of glucose homeostasis by GLP-1. Prog. Mol. Biol. Transl. Sci. 2014, 121, 23–26. [Google Scholar] [PubMed] [Green Version]
- Kulina, G.R.; Rayfield, E.J. The Role of Glucagon in the Pathophysiology and Management of Diabetes. Endocr. Pract. 2016, 22, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M.S.; Ahrén, B. Role of VIP and PACAP in islet function. Peptides 2007, 28, 1805–1813. [Google Scholar] [CrossRef]
- Yu, R.; Zhang, H.; Huang, L.; Liu, X.; Chen, J. Anti-hyperglycemic, antioxidant and anti-inflammatory effects of VIP and a VPAC1 agonist on streptozotocin-induced diabetic mice. Peptides 2011, 32, 216–222. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Claus, T.H.; Liang, Y.; Li, Y.; Yang, L.; Zhu, J.; Cruz, F.D.; Peng, X.; Chen, H.; Yung, S.L.; et al. A potent and highly selective VPAC2 agonist enhances glucose-induced insulin release and glucose disposal: A potential therapy for type 2 diabetes. Diabetes 2002, 51, 1453–1460. [Google Scholar] [CrossRef] [Green Version]
- Erendor, F.; Sahin, E.O.; Sanlioglu, A.D.; Balci, M.K.; Griffith, T.S.; Sanlioglu, S. Lentiviral gene therapy vectors encoding VIP suppressed diabetes-related inflammation and augmented pancreatic beta-cell proliferation. Gene Ther. 2021, 28, 130–141. [Google Scholar] [CrossRef]
- Klok, M.D.; Jakobsdottir, S.; Drent, M.L. The role of leptin and ghrelin in the regulation of food intake and body weight in humans: A review. Obes. Rev. 2007, 8, 21–34. [Google Scholar] [CrossRef]
- Greenwood, H.C.; Bloom, S.R.; Murphy, K.G. Peptides and their potential role in the treatment of diabetes and obesity. Rev. Diabet. Stud. 2011, 8, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Woods, S.C.; Seeley, R.J.; Porte, D., Jr.; Schwartz, M.W. Signals that regulate food intake and energy homeostasis. Science 1998, 280, 1378–1383. [Google Scholar] [CrossRef] [Green Version]
- McGowan, B.M.; Bloom, S.R. Peptide YY and appetite control. Curr. Opin. Pharmacol. 2004, 4, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, G.H.; Goldenring, J.R.; Savoca, P.E.; Kranz, H.K.; Adrian, T.E.; Bilchik, A.J.; Modlin, I.M. Cyclic AMP-mediated release of peptide YY (PYY) from the isolated perfused rabbit distal colon. Regul. Pept. 1993, 47, 117–126. [Google Scholar] [CrossRef]
- Tamakawa, H.; Miyata, A.; Satoh, K.; Miyake, Y.; Matsuo, H.; Arimura, A.; Kangawa, K. The augmentation of pituitary adenylate cyclase-activating polypeptide (PACAP) in streptozotocin-induced diabetic rats. Peptides 1998, 19, 1497–1502. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanford, D.; Luong, L.; Vu, J.P.; Oh, S.; Gabalski, A.; Lewis, M.; Pisegna, J.R.; Germano, P. The VIP/VPAC1R Pathway Regulates Energy and Glucose Homeostasis by Modulating GLP-1, Glucagon, Leptin and PYY Levels in Mice. Biology 2022, 11, 431. https://doi.org/10.3390/biology11030431
Sanford D, Luong L, Vu JP, Oh S, Gabalski A, Lewis M, Pisegna JR, Germano P. The VIP/VPAC1R Pathway Regulates Energy and Glucose Homeostasis by Modulating GLP-1, Glucagon, Leptin and PYY Levels in Mice. Biology. 2022; 11(3):431. https://doi.org/10.3390/biology11030431
Chicago/Turabian StyleSanford, Daniel, Leon Luong, John P. Vu, Suwan Oh, Arielle Gabalski, Michael Lewis, Joseph R. Pisegna, and Patrizia Germano. 2022. "The VIP/VPAC1R Pathway Regulates Energy and Glucose Homeostasis by Modulating GLP-1, Glucagon, Leptin and PYY Levels in Mice" Biology 11, no. 3: 431. https://doi.org/10.3390/biology11030431
APA StyleSanford, D., Luong, L., Vu, J. P., Oh, S., Gabalski, A., Lewis, M., Pisegna, J. R., & Germano, P. (2022). The VIP/VPAC1R Pathway Regulates Energy and Glucose Homeostasis by Modulating GLP-1, Glucagon, Leptin and PYY Levels in Mice. Biology, 11(3), 431. https://doi.org/10.3390/biology11030431