
The Effect of Feeding on Briareum violacea Growth, Survival and Larval Development under Temperature and Salinity Stress


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Coral Samples
2.2. Experimental Conditions
2.3. Feed Source
2.4. Experiment 1: Effects of Temperature, Salinity and Diet on Body Composition, Digestive Enzymes, Reproduction and Stress Response of Briareum violacea
2.4.1. Determination of Briareum violacea Growth and Survival
2.4.2. Analysis of Coral Body Composition
2.4.3. Analysis of Digestive Enzymes
2.4.4. Analysis of Zooxanthellar Density and Chlorophyll a
2.4.5. Antioxidant Enzyme Analysis
Preparation of Coral Tissue Solution
Superoxide Dismutase Detection
Catalase Detection
Protein Concentrations
2.4.6. Egg Production Calculation and Egg Size Measurement
2.5. Experiment 2: Effect of Temperature, Salinity and Diet on the Settlement and Survival of Briareum violacea Larvae
2.5.1. Experimental Conditions
2.5.2. Determination of Zooxanthellae Number and Chlorophyll a in Briareum violacea Larvae
2.6. Statistical Analysis
3. Results
3.1. Experiment 1: Evaluation of the Response of Feeding Bait to Thermohaline Stress in B. violacea
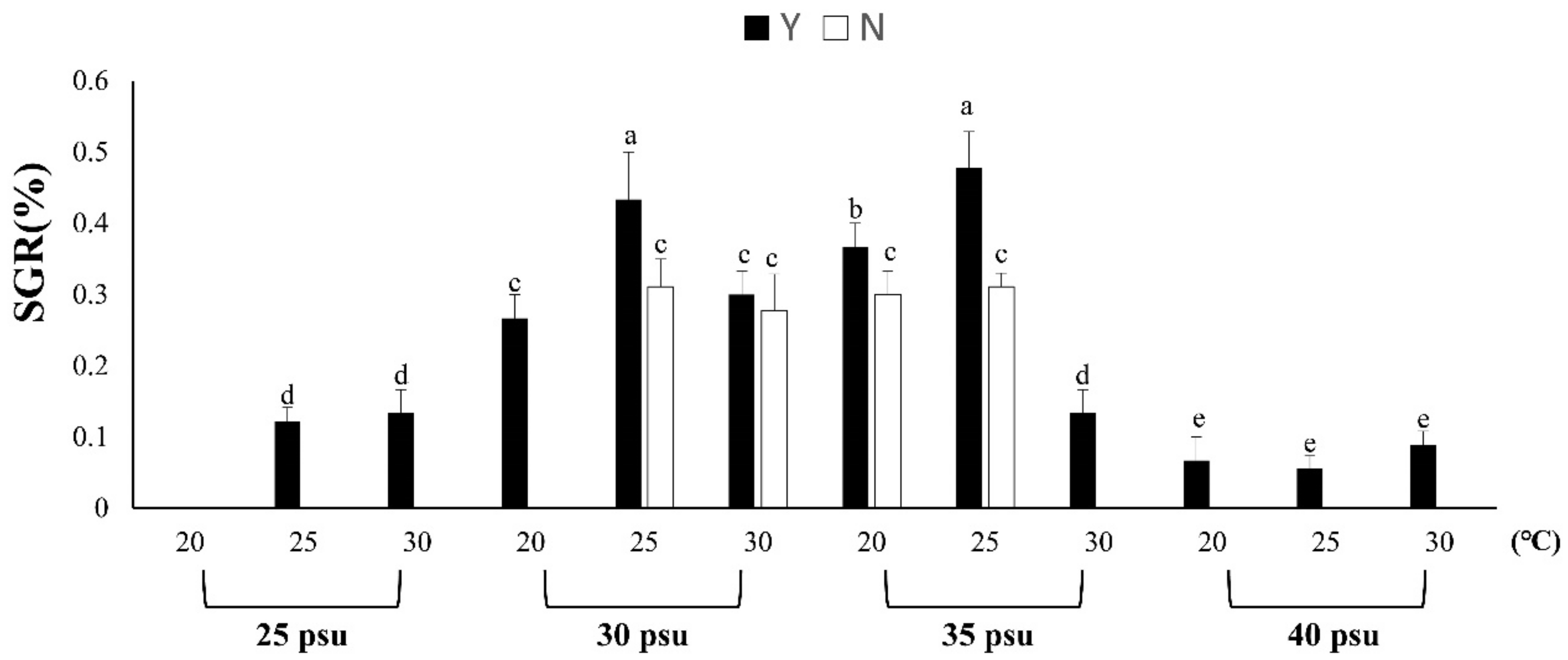
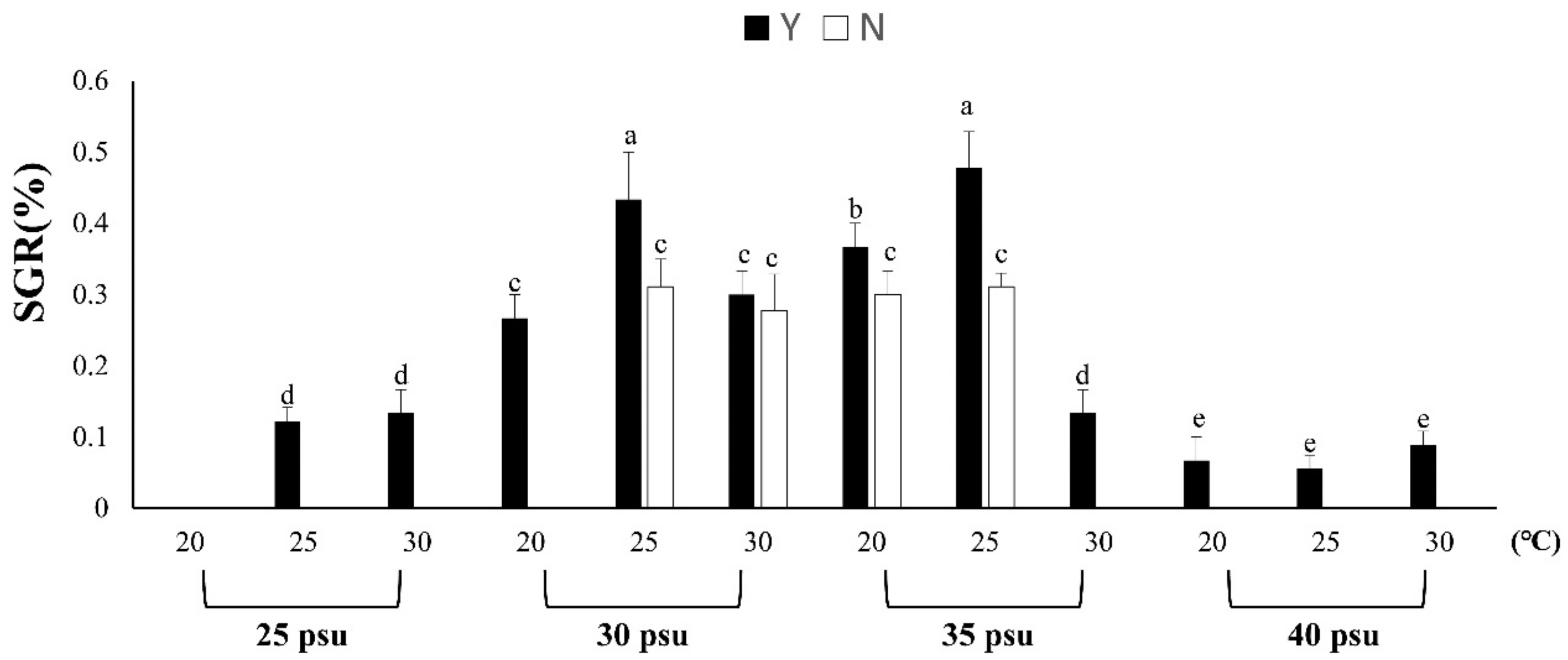
3.1.1. Effects of Temperature, Salinity and Feeding on B. violacea Growth and Survival
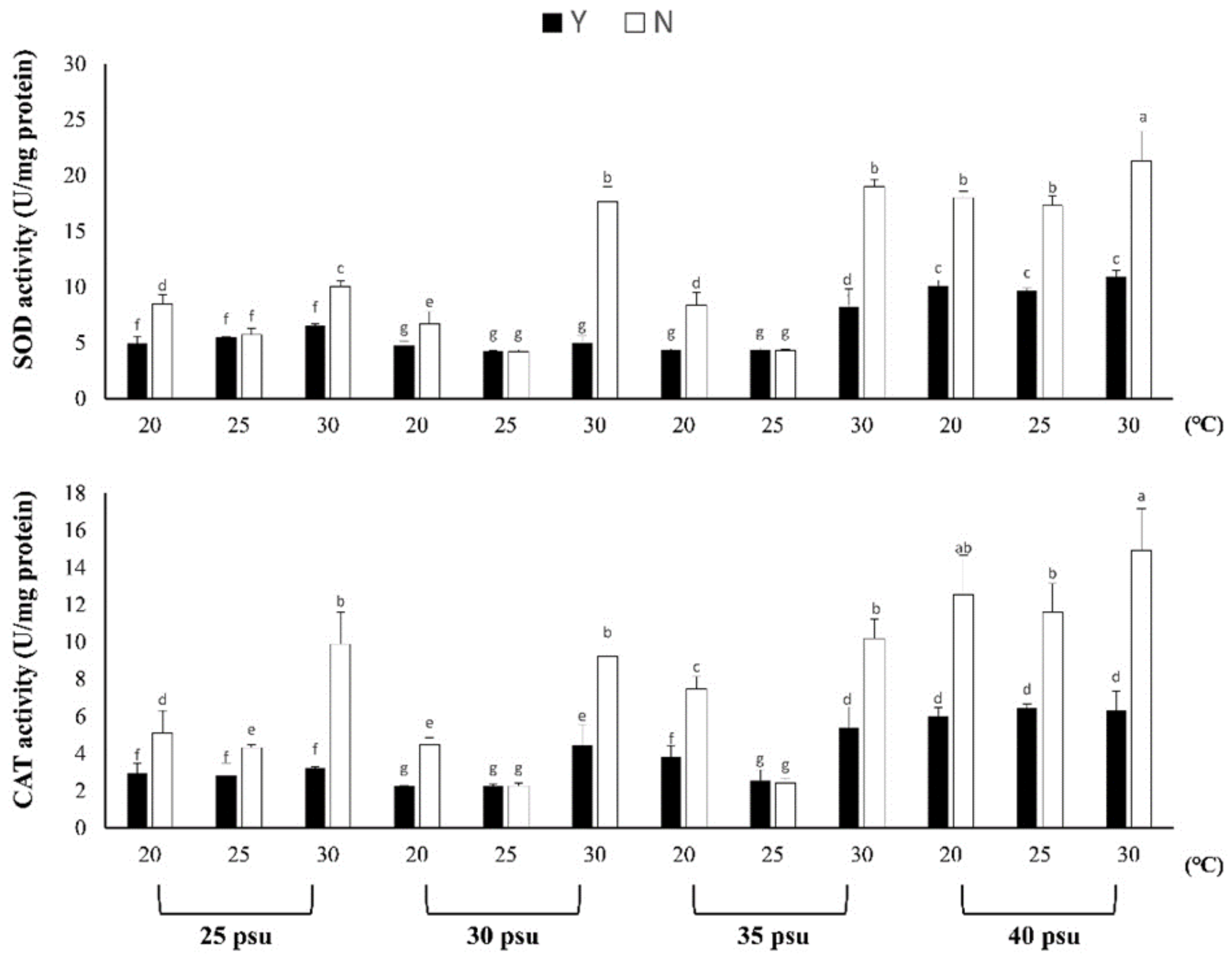
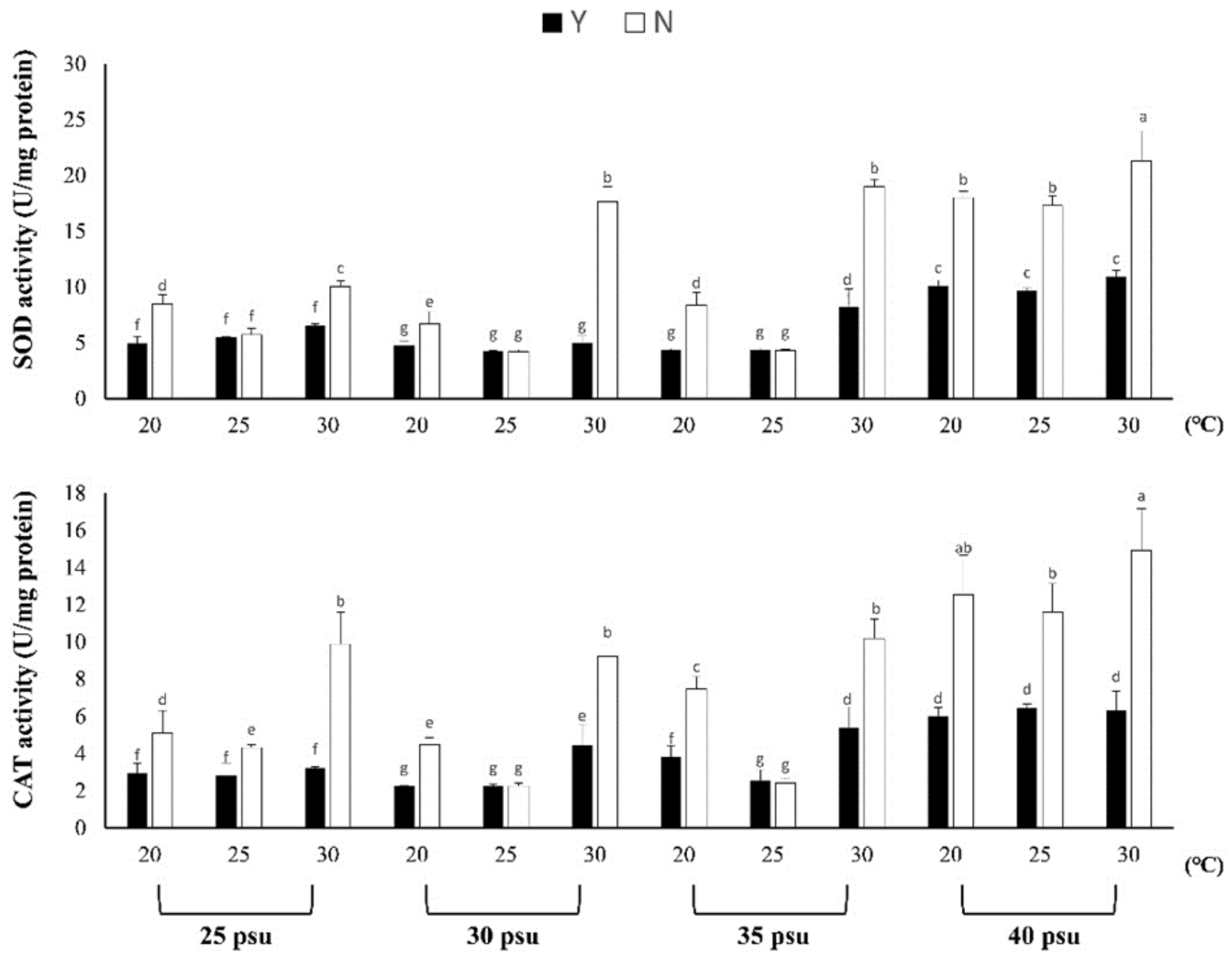
3.1.2. Evaluation of Resistance to Stress by Feeding
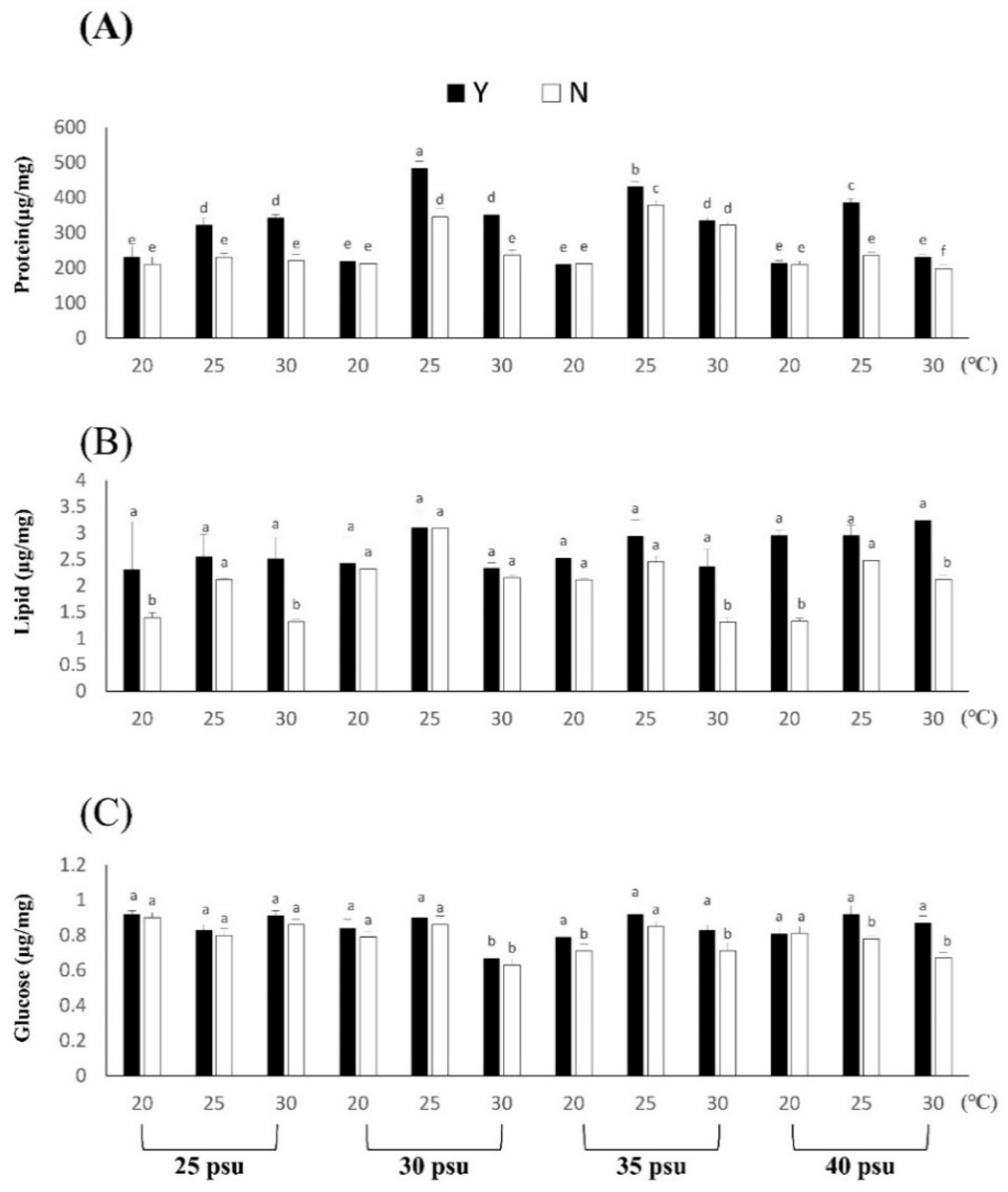
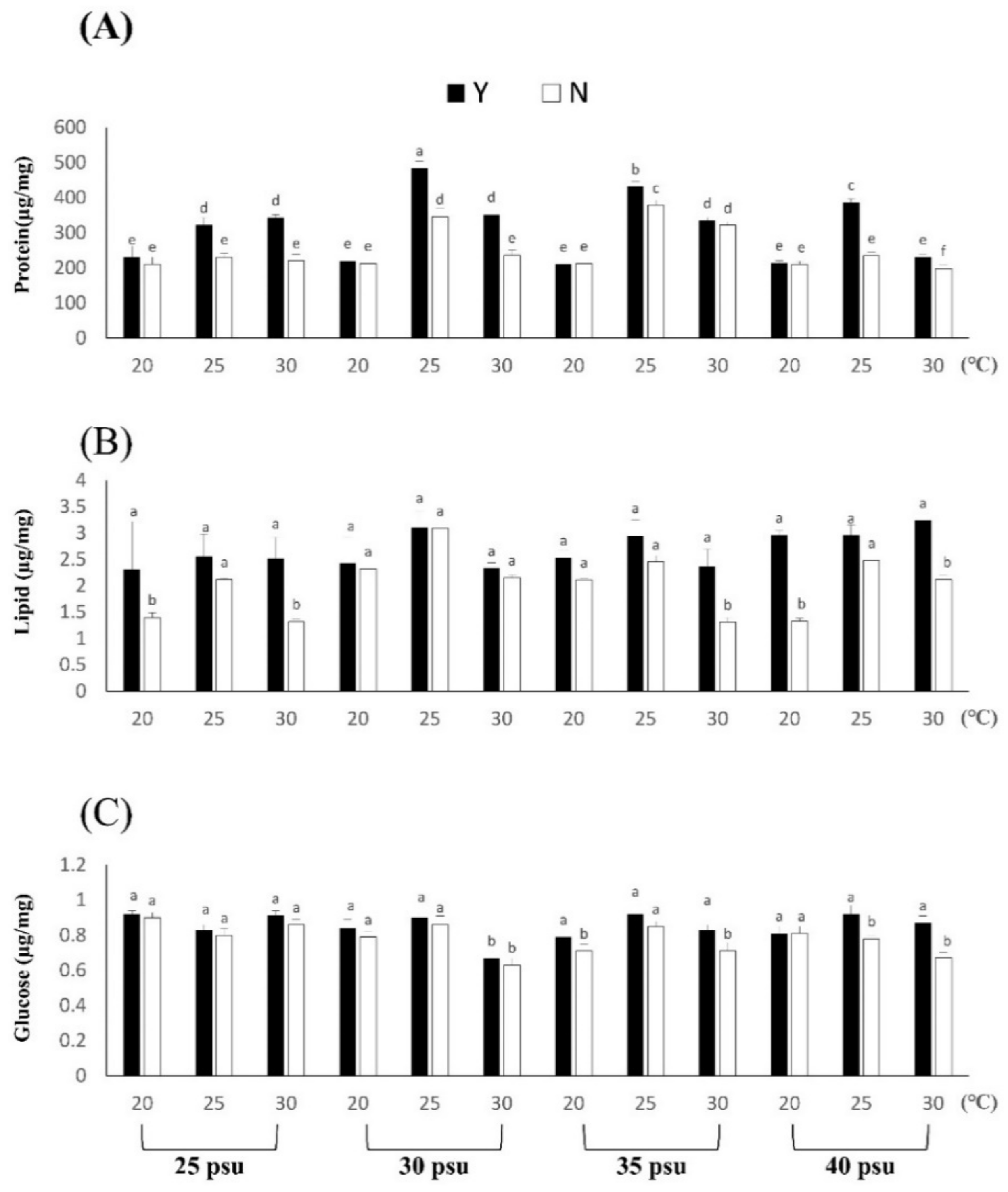
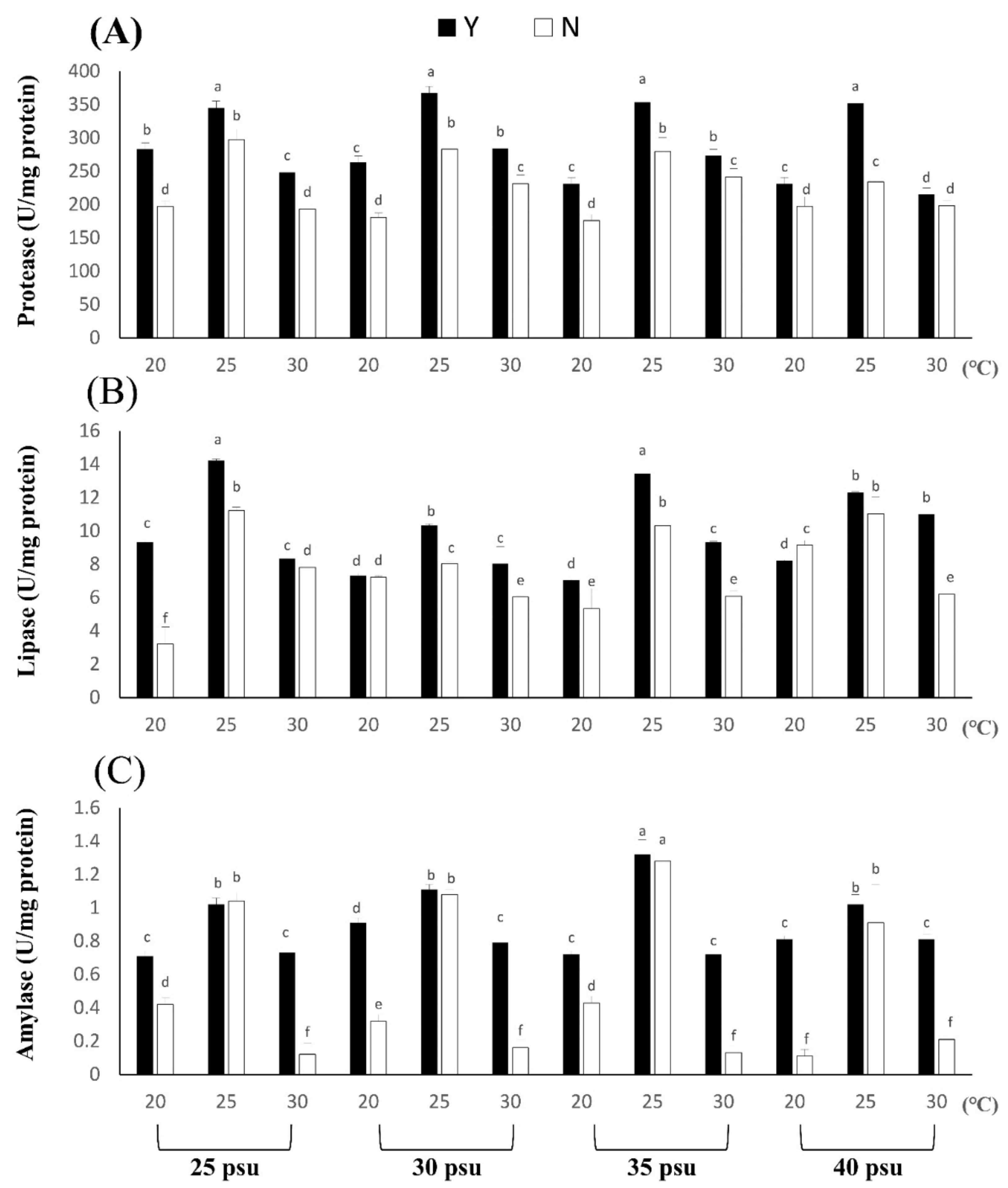
3.1.3. Effects of Temperature and Salinity Stress on Body Composition and Digestive Enzymes
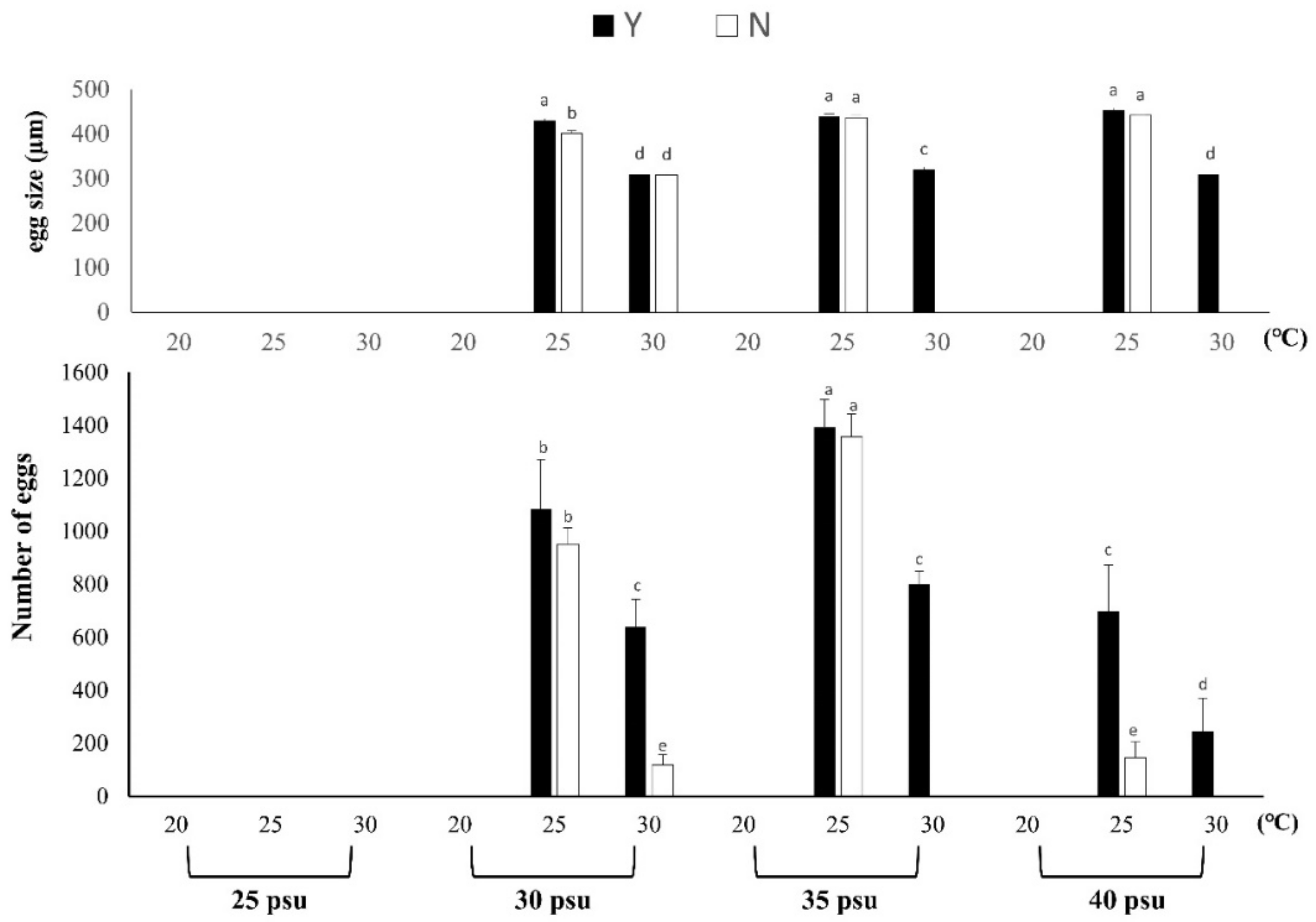
3.1.4. Spawning Volume and Egg Size
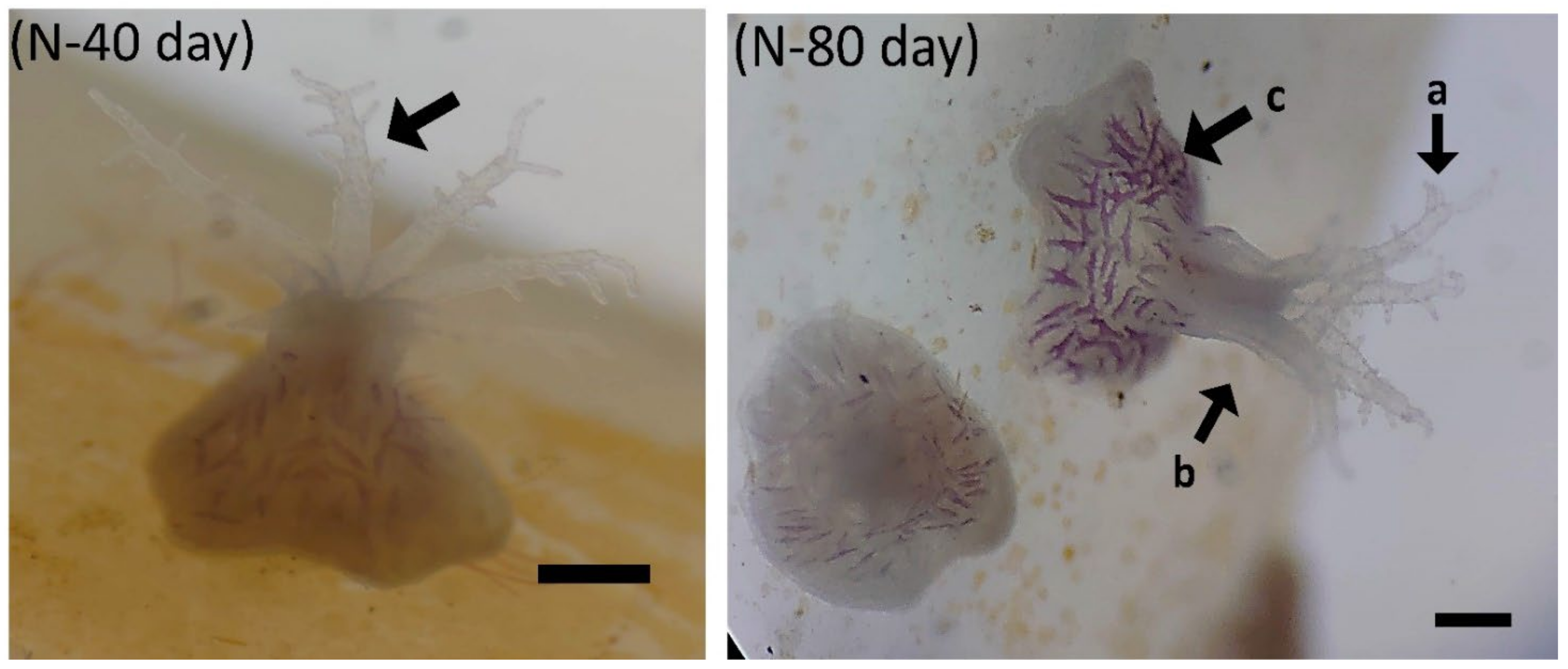
3.2. Experiment 2: Larval Settlement and Larval Development of B. violacea
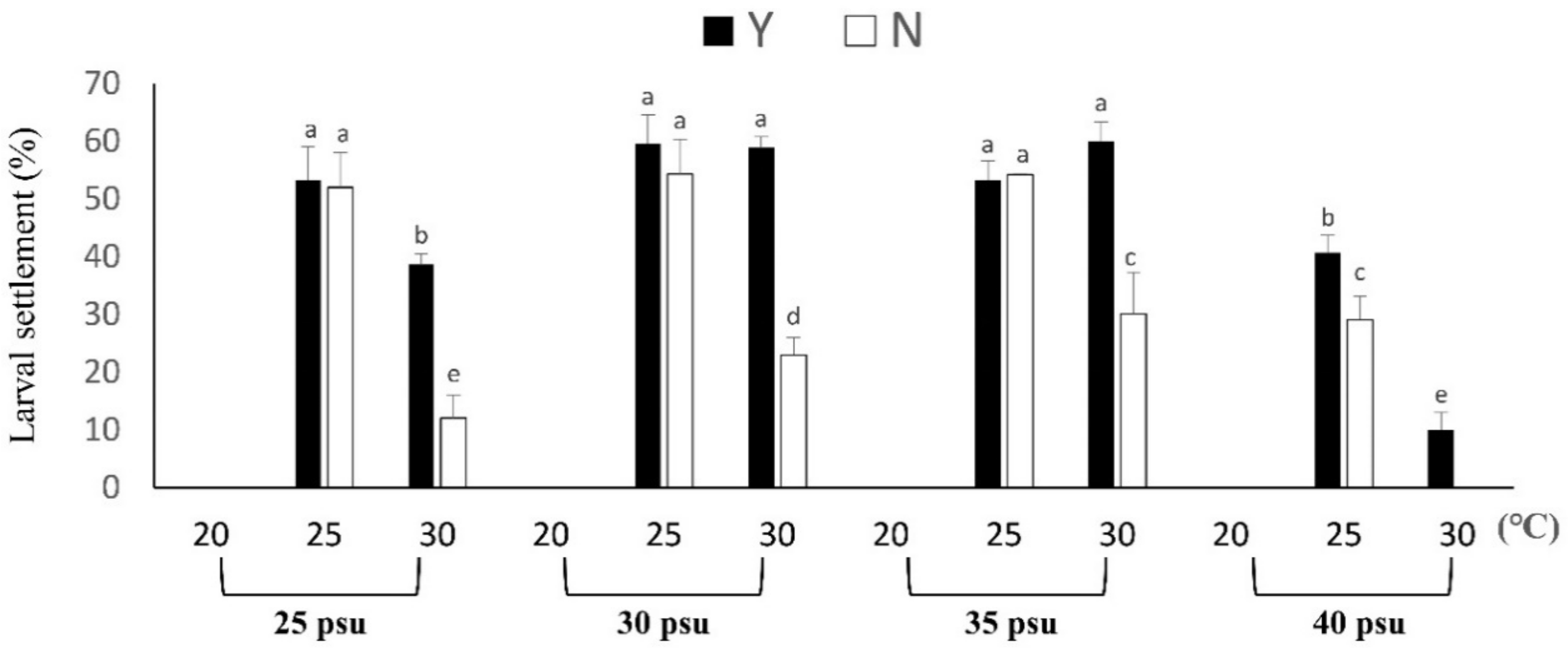
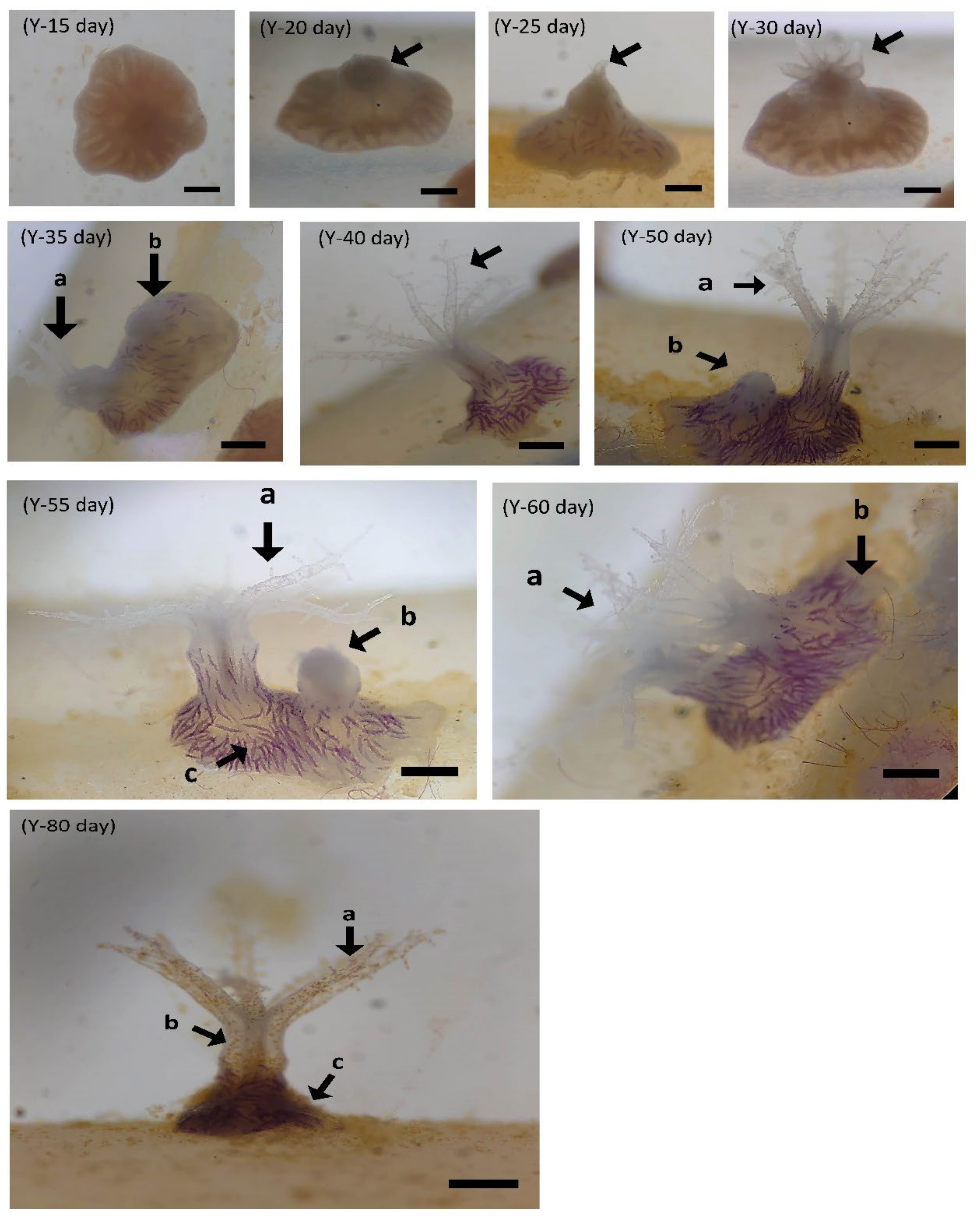
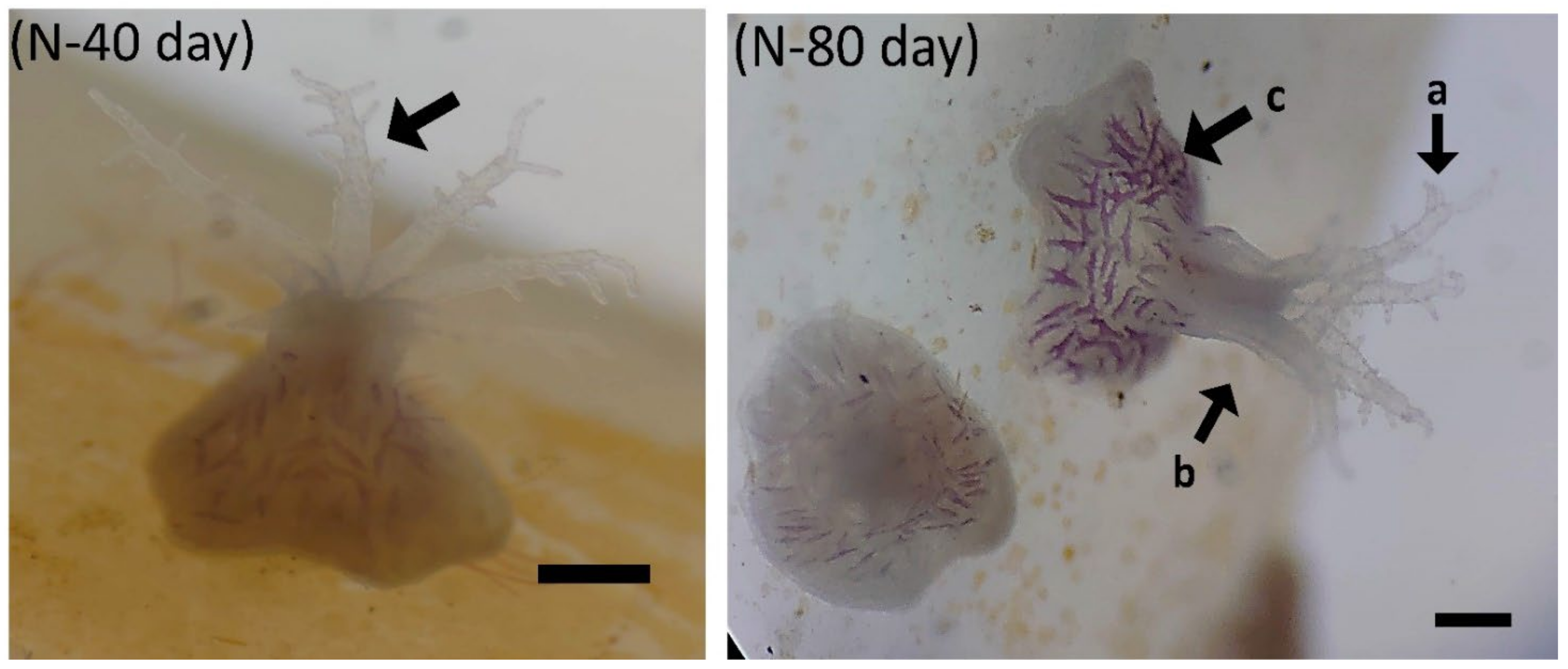
3.2.1. Larval Settlement
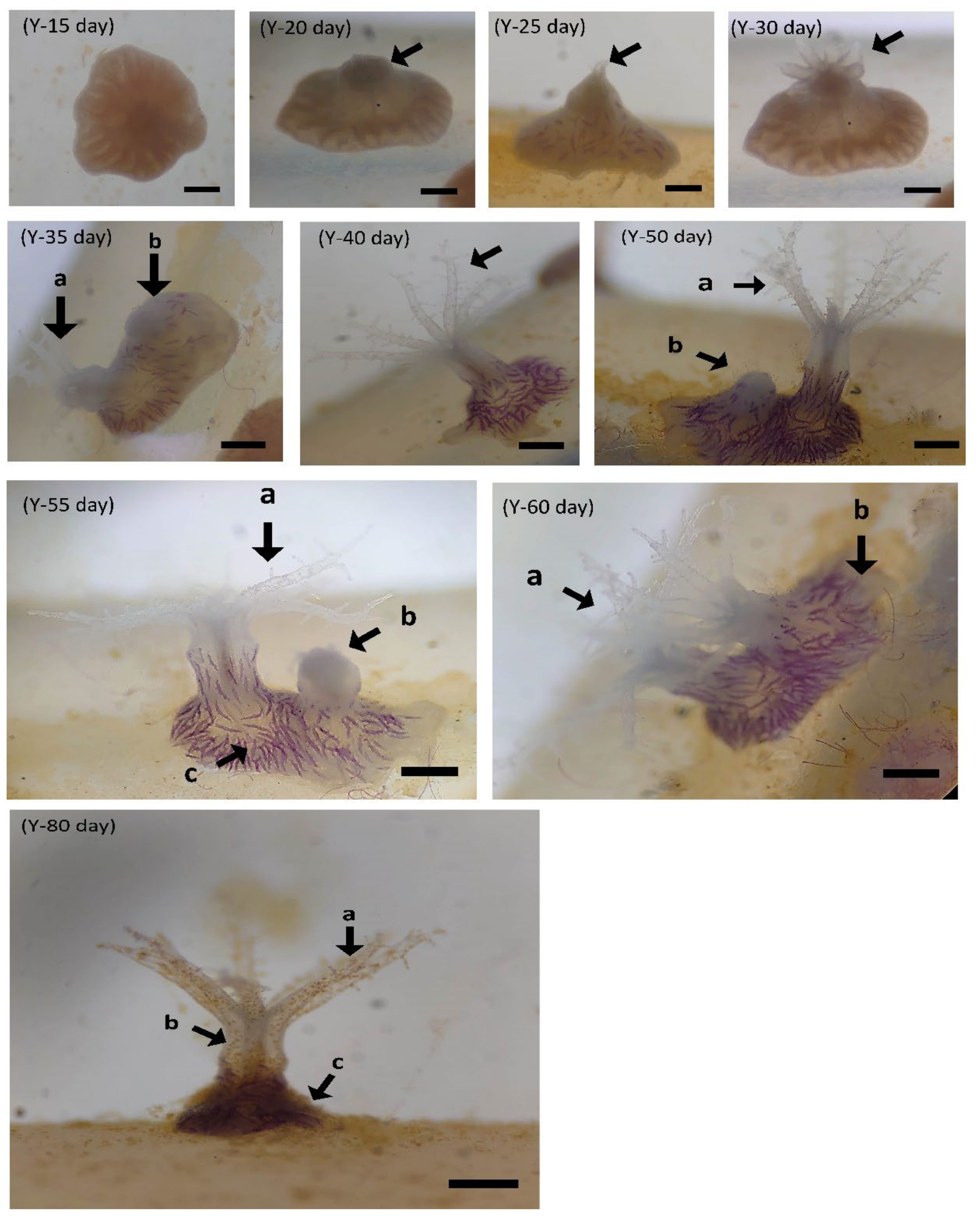
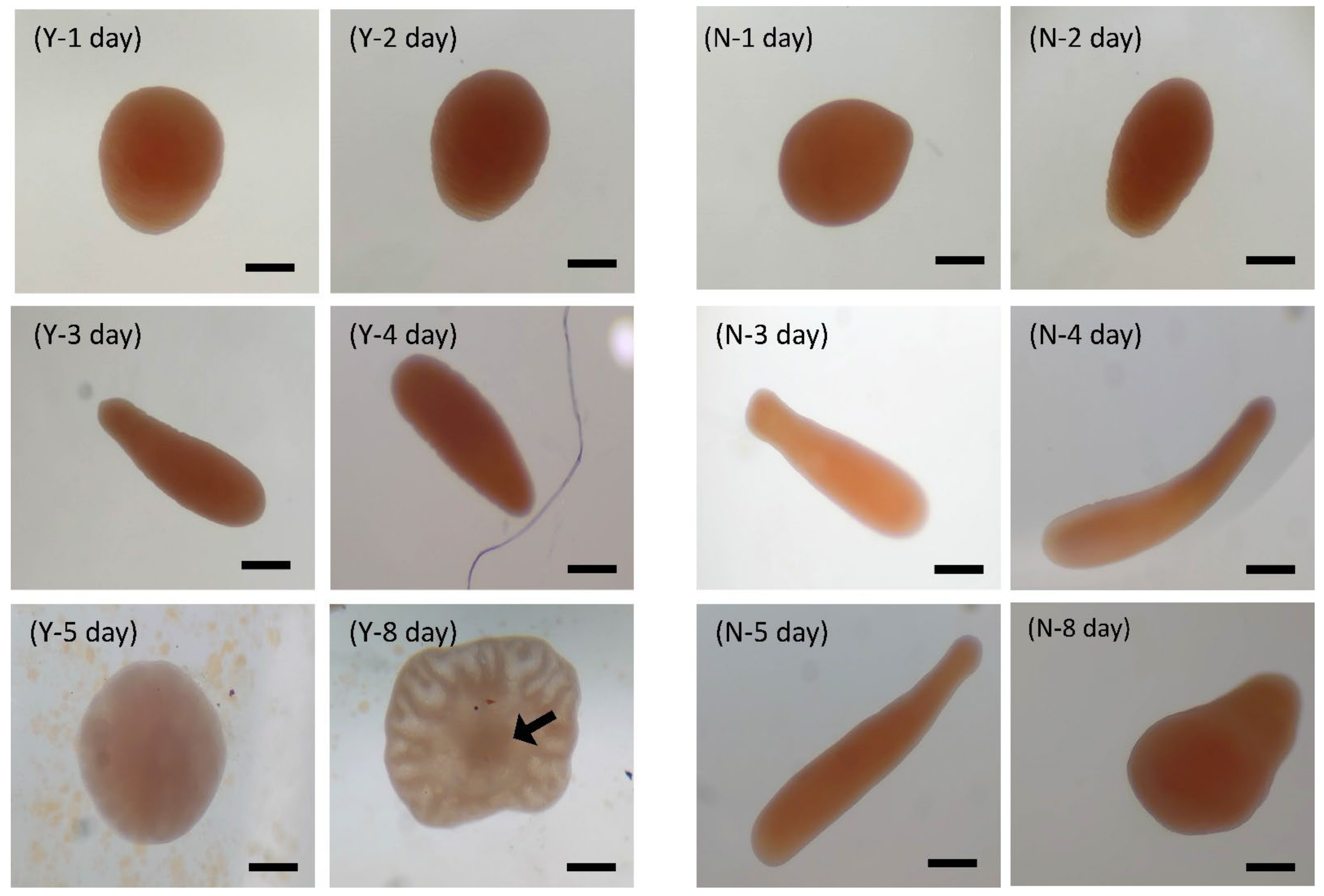
3.2.2. Larval Development of B. violacea
3.2.3. Larval Zooxanthellae and Chlorophyll a
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, K.E.; Abrar, M.; Aeby, G.; Aronson, R.B.; Banks, S.; Bruckner, A.; Chiriboga, A.; Cortés, J.; Delbeek, J.C.; DeVantier, L.; et al. One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 2008, 321, 560–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veron, J.; Hoegh-Guldberg, O.; Lenton, T.; Lough, J.; Obura, D.; Pearce-Kelly, P.; Sheppard, C.; Spalding, M.; Stafford-Smith, M.; Rogers, A. The coral reef crisis: The critical importance of <350 ppm CO2. Mar. Pollut. Bull. 2009, 58, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Smith, G.J. The effect of sudden changes in temperature, light and salinity on the population density and export of zooxanthellae from the reef corals Stylophora pistillata Esper and Seriatopora hystrix Dana. J. Exp. Mar. Biol. Ecol. 1989, 129, 279–303. [Google Scholar] [CrossRef]
- Ferrier-Pages, C.; Gattuso, J.-P.; Jaubert, J. Effect of small variations in salinity on the rates of photosynthesis and respiration of the zooxanthellate coral Stylophora pistillata. Mar. Ecol. Prog. Ser. 1999, 181, 309–314. [Google Scholar] [CrossRef]
- True, J.D. Salinity as a Structuring Force for near Shore Coral Communities. In Proceedings of the 12th International Coral Reef Symposium, Cairns, Australia, 9–13 July 2012. [Google Scholar]
- Bellwood, D.R.; Hughes, T.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Berkelmans, R.; Jones, A.M.; Schaffelke, B. Salinity thresholds of Acropora spp. on the Great Barrier Reef. Coral Reefs 2012, 31, 1103–1110. [Google Scholar] [CrossRef]
- Hédouin, L.; Pilon, R.; Puisay, A. Hyposalinity stress compromises the fertilization of gametes more than the survival of coral larvae. Mar. Environ. Res. 2015, 104, 1–9. [Google Scholar] [CrossRef]
- Reynaud, S.; Leclercq, N.; Romaine-Lioud, S.; Ferrier-Pagés, C.; Jaubert, J.; Gattuso, J.-P. Interacting effects of CO2 partial pressure and temperature on photosynthesis and calcification in a scleractinian coral. Glob. Change Biol. 2003, 9, 1660–1668. [Google Scholar] [CrossRef] [Green Version]
- Langdon, C.; Atkinson, M.J. Effect of elevated pCO2 on photosynthesis and calcification of corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. J. Geophys. Res. Oceans 2005, 110, 1–54. [Google Scholar] [CrossRef]
- Anthony, K.R.N.; Kline, D.; Diaz-Pulido, G.; Dove, S.; Hoegh-Guldberg, O. Ocean acidification causes bleaching and productivity loss in coral reef builders. Proc. Natl. Acad. Sci. USA 2008, 105, 17442–17446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Katwijk, M.; Meier, N.F.; Van Loon, R.; Van Hove, E.M.; Giesen, W.B.J.T.; Van Der Velde, G.; Hartog, C.D. Sabaki River sediment load and coral stress: Correlation between sediments and condition of the Malindi-Watamu reefs in Kenya (Indian Ocean). Mar. Biol. 1993, 117, 675–683. [Google Scholar] [CrossRef]
- Torres, R.; Chiappone, M.; Geraldes, F.; Rodriguez, Y.; Vega, M. Sedimentation as an important environmental influence on Dominican Republic reefs. Bull. Mar. Sci. 2001, 69, 805–818. [Google Scholar]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Harriott, V.; Banks, S. Latitudinal variation in coral communities in eastern Australia: A qualitative biophysical model of factors regulating coral reefs. Coral Reefs 2002, 21, 83–94. [Google Scholar] [CrossRef]
- Chui, A.P.Y.; Ang, P. Elevated temperature enhances normal early embryonic development in the coral Platygyra acuta under low salinity conditions. Coral Reefs 2015, 34, 461–469. [Google Scholar] [CrossRef]
- Coles, S.; Jokiel, P.L. Synergistic effects of temperature, salinity and light on the hermatypic coral Montipora verrucosa. Mar. Biol. 1978, 49, 187–195. [Google Scholar] [CrossRef]
- Kuanui, P.; Chavanich, S.; Viyakarn, V.; Omori, M.; Lin, C. Effects of temperature and salinity on survival rate of cultured corals and photosynthetic efficiency of zooxanthellae in coral tissues. Ocean Sci. J. 2015, 50, 263–268. [Google Scholar] [CrossRef]
- Burke, L.; Reytar, K.; Spalding, M.; Perry, A. Reefs at Risk Revisited; World Resources Institute: Washington, DC, USA, 2011. [Google Scholar]
- Wilkinson, C. Status of Coral Reefs of the World: 2008; Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre: Townsville, Australia, 2008. [Google Scholar]
- Amador, M.L.; Jimeno, J.; Paz-Ares, L.; Cortes-Funes, H.; Hidalgo, M. Progress in the development and acquisition of anticancer agents from marine sources. Ann. Oncol. 2003, 14, 1607–1615. [Google Scholar] [CrossRef]
- Fenical, W.; Jensen, P.; Palladino, M.A.; Lam, K.S.; Lloyd, G.K.; Potts, B.C. Discovery and development of the anticancer agent salinosporamide A (NPI-0052). Bioorg. Med. Chem. 2009, 17, 2175–2180. [Google Scholar] [CrossRef] [Green Version]
- Rocha, J.; Peixe, L.; Gomes, N.C.; Calado, R. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.E. Worldwide death of corals—Natural cyclical events or man-made pollution? Mar. Pollut. Bull. 1987, 18, 9–13. [Google Scholar] [CrossRef]
- Chavanich, S.; Siripong, A.; Sojisuporn, P.; Menasveta, P. Impact of tsunami on the seafloor and corals in Thailand. Coral Reefs 2005, 24, 535. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, K.R.T.; Miller, J.; Sanchez, O.; Patterson, M. Impact of the 2005 coral bleaching event on Porites porites and Colpophyllia natans at Tektite Reef, U.S. Virgin Islands. Coral Reefs 2007, 26, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Eakin, C.; Lough, J.; Heron, S. Climate Variability and Change: Monitoring Data and Evidence for Increased Coral Bleaching Stress. In Coral Bleaching; Springer: Berlin/Heidelberg, Germany, 2009; pp. 41–67. [Google Scholar]
- Brown, B.E.; Phongsuwan, N. Delayed mortality in bleached massive corals on intertidal reef flats around Phuket, Andaman Sea, Thailand. Phuket Mar. Biol. Cent. Res. Bull. 2012, 71, 43–48. [Google Scholar]
- Chavanich, S.; Viyakarn, V. Reef communities after the 2010 mass coral bleaching at Racha Yai Island in the Andaman Sea and Koh Tao in the Gulf of Thailand. Phuket Mar. Biol. Cent. Res. Bull. 2012, 71, 103–110. [Google Scholar]
- Phongsuwan, N.; Chansang, H. Repeated coral bleaching in the Andaman Sea, Thailand, during the last two decades. Phuket Mar. Biol. Cent. Res. Bull. 2012, 71, 19–41. [Google Scholar]
- Baird, A.H.; Bhagooli, R.; Ralph, P.; Takahashi, S. Coral bleaching: The role of the host. Trends Ecol. Evol. 2009, 24, 16–20. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Lesser, M.P.; Stochaj, W.R.; Tapley, D.W.; Shick, J.M. Bleaching in coral reef anthozoans: Effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 1990, 8, 225–232. [Google Scholar] [CrossRef]
- Dykens, J.A.; Shick, J.M.; Benoit, C.; Buettner, G.R.; Winston, G.W. Oxygen radical production in the sea anemone Anthopleura elegantissima and its endosymbiotic algae. J. Exp. Biol. 1992, 168, 219–241. [Google Scholar] [CrossRef]
- Downs, C.; Fauth, E.J.; Halas, J.C.; Dustan, P.; Bemiss, J.; Woodley, C.M. Oxidative stress and seasonal coral bleaching. Free Radic. Biol. Med. 2002, 33, 533–543. [Google Scholar] [CrossRef]
- Higuchi, T.; Fujimura, H.; Arakaki, T.; Oomori, T. Activities of Antioxidant Enzymes (SOD and CAT) in the Coral Galaxea fascicularis against Increased Hydrogen Peroxide Concentrations in Seawater. In Proceedings of the 11th International Coral Reef Symposium, Fort Lauderdale, FL, USA, 7–11 July 2008. [Google Scholar]
- Shick, J.M.; Lesser, M.P.; Dunlap, W.C.; Stochaj, W.R.; Chalker, B.E.; Won, J.W. Depth-dependent responses to solar ultraviolet radiation and oxidative stress in the zooxanthellate coral Acropora microphthalma. Mar. Biol. 1995, 122, 41–51. [Google Scholar] [CrossRef]
- Ding, D.-S.; Sun, W.-T.; Pan, C.-H. Feeding of a Scleractinian Coral, Goniopora columna, on Microalgae, Yeast, and Artificial Feed in Captivity. Animals 2021, 11, 3009. [Google Scholar] [CrossRef]
- Johnson, C.R.; Preece, A.L. Damage, Scale and Recovery in Model Coral Communities: The Importance of System State. In Proceedings of the 7th International Coral Reef Symposium, Guam, Micronesia, 22–27 June 1992. [Google Scholar]
- Connell, J. Disturbance and recovery of coral assemblages. Coral Reefs 1997, 16, S101–S113. [Google Scholar] [CrossRef]
- Sammarco, P.W. Polyp bail-out: An escape response to environmental stress and a new means of reproduction in corals. Mar. Ecol. Prog. Ser. 1982, 10, 57–65. [Google Scholar] [CrossRef]
- Dias, M.; Madeira, C.; Jogee, N.; Ferreira, A.; Gouveia, R.; Cabral, H.; Diniz, M.; Vinagre, C. Oxidative stress on scleractinian coral fragments following exposure to high temperature and low salinity. Ecol. Indic. 2019, 107, 105586. [Google Scholar] [CrossRef]
- Bonaldo, R.M.; Hay, M.E. Hay, Seaweed-coral interactions: Variance in seaweed allelopathy, coral susceptibility, and potential effects on coral resilience. PLoS ONE 2014, 9, e85786. [Google Scholar]
- Rocha, R.J.; Pimentel, T.; Serôdio, J.; Rosa, R.; Calado, R. Comparative performance of light emitting plasma (LEP) and light emitting diode (LED) in ex situ aquaculture of scleractinian corals. Aquaculture 2013, 402, 38–45. [Google Scholar] [CrossRef]
- Schutter, M.; van Velthoven, B.; Janse, M.; Osinga, R.; Janssen, M.; Wijffels, R.; Verreth, J. The effect of irradiance on long-term skeletal growth and net photosynthesis in Galaxea fascicularis under four light conditions. J. Exp. Mar. Biol. Ecol. 2008, 367, 75–80. [Google Scholar] [CrossRef]
- AOAC. Official Methods for the Analysis, 14th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1984. [Google Scholar]
- Duben-Engelkirk, J.L.; Fody, E.P. Clinical Chemistry: Principles, Procedures, Correlations; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Tietze, W. Fundamentals of Clinical Chemistry, 2nd ed.; WB Saunders Co.: New York, NY, USA, 1982; pp. 994–995. [Google Scholar]
- Sun, J.-Y.; Du, J.; Qian, L.-C.; Jing, M.-Y.; Weng, X.-Y. Distribution and characteristics of endogenous digestive enzymes in the red-eared slider turtle, Trachemys cripta elegans. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Borlongan, I.G. Studies on the digestive lipases of milkfish, Chanos chanos. Aquaculture 1990, 89, 315–325. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases, α and β. Methods Enzymol. 1955, 1, 149–158. [Google Scholar]
- Titlyanov, E.; Titlyanova, T.; Yamazato, K.; van Woesik, R. Photo-acclimation of the hermatypic coral Stylophora pistillata while subjected to either starvation or food provisioning. J. Exp. Mar. Biol. Ecol. 2001, 257, 163–181. [Google Scholar] [CrossRef]
- Lajeunesse, T.C.; Parkinson, J.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [Green Version]
- Levy, O.; Dubinsky, Z.; Achituv, Y. Photobehavior of stony corals: Responses to light spectra and intensity. J. Exp. Biol. 2003, 206, 4041–4049. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Liñán-Cabello, M.A.; Flores-Ramirez, L.A.; Zeneto-Savin, T.; Olguin-Monroy, N.O.; Sosa-Avalos, R.; Patino-Barragan, M.; Olivos-Ortiz, A. Seasonal changes of antioxidant and oxidative parameters in the coral Pocillopora capitata on the Pacific coast of Mexico. Mar. Ecol. 2010, 31, 407–417. [Google Scholar]
- Cheng, C.-M.; Cheng, Y.-R.; Ding, D.-S.; Chen, Y.-T.; Sun, W.-T.; Pan, C.-H. Effects of Ciliate Infection on the Activities of Two Antioxidant Enzymes (SOD and CAT) in Captive Coral (Goniopora columna) and Evaluation of Drug Therapy. Biology 2021, 10, 1216. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Main, W.; Ross, C.; Bielmyer, G. Copper accumulation and oxidative stress in the sea anemone, Aiptasia pallida, after waterborne copper exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Paxton, C.W.; Baria, M.V.B.; Weis, V.M.; Harii, S. Effect of elevated temperature on fecundity and reproductive timing in the coral Acropora digitifera. Zygote 2016, 24, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Moorsel, G. Reproductive strategies in two closely related stony corals (Agaricia, Scleractinia). Mar. Ecol. Prog. Ser. 1983, 13, 273–283. [Google Scholar] [CrossRef]
- Miller, K.; Mundy, C. Rapid settlement in broadcast spawning corals: Implications for larval dispersal. Coral Reefs 2003, 22, 99–106. [Google Scholar] [CrossRef]
- Jin, Y.K.; Kininmonth, S.; Lundgren, P.B.; Van Oppen, M.J.H.; Willis, B.L. Predicting the spatial distribution of allele frequencies for a gene associated with tolerance to eutrophication and high temperature in the reef-building coral, Acropora millepora, on the Great Barrier Reef. Coral Reefs 2020, 39, 147–158. [Google Scholar] [CrossRef]
- Humphrey, C.; Weber, M.; Lott, C.; Cooper, T.; Fabricius, K. Effects of suspended sediments, dissolved inorganic nutrients and salinity on fertilisation and embryo development in the coral Acropora millepora (Ehrenberg, 1834). Coral Reefs 2008, 27, 837–850. [Google Scholar] [CrossRef] [Green Version]
- Howells, E.J.; Ketchum, R.N.; Bauman, A.G.; Mustafa, Y.; Watkins, K.D.; Burt, J. Species-specific trends in the reproductive output of corals across environmental gradients and bleaching histories. Mar. Pollut. Bull. 2016, 105, 532–539. [Google Scholar] [CrossRef]
- Mos, B.; Byrne, M.; Dworjanyn, S.A. Effects of low and high pH on sea urchin settlement, implications for the use of alkali to counter the impacts of acidification. Aquaculture 2020, 528, 735618. [Google Scholar] [CrossRef]
- Babcock, R.; Heyward, A.J. Larval development of certain gamete-spawning scleractinian corals. Coral Reefs 1986, 5, 111–116. [Google Scholar] [CrossRef]
- Shlesinger, Y.; Loya, Y. Larval Development and Survivorship in the Corals Favia favus and Platygyra lamellina. In Coelenterate Biology: Recent Research on Cnidaria and Ctenophora; Springer: Dordrecht, Germany, 1991; pp. 101–108. [Google Scholar]
- Nozawa, Y.; Harrison, P.L. Effects of elevated temperature on larval settlement and post-settlement survival in scleractinian corals, Acropora solitaryensis and Favites chinensis. Mar. Biol. 2007, 152, 1181–1185. [Google Scholar] [CrossRef]
- Yakovleva, I.M.; Baird, A.H.; Yamamoto, H.H.; Bhagooli, R.; Nonaka, M.; Hidaka, M. Algal symbionts increase oxidative damage and death in coral larvae at high temperatures. Mar. Ecol. Prog. Ser. 2009, 378, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Nesa, B.; Baird, A.H.; Harii, S.; Yakovleva, I.; Hidaka, M. Algal symbionts increase DNA damage in coral planulae exposed to sunlight. Zool. Stud. 2012, 51, 12–17. [Google Scholar]
- Haryanti, D.; Yasuda, N.; Harii, S.; Hidaka, M. High tolerance of symbiotic larvae of Pocillopora damicornis to thermal stress. Zool. Stud. 2015, 54, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baria, M.V.B.; Kurihara, H.; Harii, S. Tolerance to elevated temperature and ocean acidification of the larvae of the solitary corals Fungia fungites (Linnaues, 1758) and Lithophyllon repanda (Dana, 1846). Zool. Sci. 2015, 32, 447–454. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Quality Conditions | Water Quality Data |
|---|---|
| pH | 8.0 ± 0.31 |
| Ammonia nitrogen | 0.04 ± 0.06 mg/L |
| Nitrous acid | 0.02 ± 0.04 mg/L |
| Nitric acid | 0.10 ± 0.01 mg/L |
| Calcium | 400 ± 44.05 ppm |
| Magnesium | 1280 ± 75.03 ppm |
| Phosphate | 0.02 ± 0.01 ppm |
| Nutritional Indicators | Protein | Lipid | Glucose |
|---|---|---|---|
| (µg/mg) | (µg/mg) | (µg/mg) | |
| Nutrient composition | 76.69 a | 6.04 c | 19.00 b |
| (Feed nutrition) | (1.49) | (0.03) | (0.32) |
| No feeding | 305.12 b | 1.11 b | 2.41 c |
| (Coral body composition) | (14.23) | (0.31) | (0.36) |
| 1 h after being fed | 403.13 a | 1.31 b | 3.72 b |
| (Coral body composition) | (19.20) | (0.08) | (0.24) |
| Salinity and Temperature | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 psu | 30 psu | 35 psu | 40 psu | ||||||||||
| Feeding | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | |
| Y | Net Increase | 0.00 | 3.67 c | 8.00 b | 4.00 c | 13.00 a | 9.00 b | 11.00 ab | 14.33 a | 4.00 c | 2.00 d | 1.67 d | 2.67 d |
| (Number ± 95%) | (0.00) | (0.58) | (1.00) | (1.00) | (2.00) | (1.00) | (1.00) | (1.53) | (1.00) | (1.00) | (0.58) | (0.58) | |
| N | Net Increase | 0.00 | 0.00 | 0.00 | 0.00 | 9.33 a | 8.33 a | 9.00 a | 9.33 a | 0.00 | 0.00 | 0.00 | 0.00 |
| (Number ± 95%) | (0.00) | (0.00) | (0.00) | (0.00) | (1.15) | (1.53) | (1.00) | (0.58) | (0.00) | (0.00) | (0.00) | (0.00) | |
| Salinity and Temperature | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 psu | 30 psu | 35 psu | 40 psu | ||||||||||
| Feeding | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | |
| Y | Zooxanthellae | 0.7 h | 1.7 e | 1.0 g | 1.50 f | 4.3 b | 3.13 d | 3.72 c | 4.82 a | 1.73 e | 0.30 i | 0.15 i | 0.13 i |
| (cell × 107 cm−2) | (0.04) | (0.03) | (0.02) | (0.05) | (0.07) | (0.03) | (0.03) | (0.02) | (0.06) | (0.05) | (0.03) | (0.05) | |
| Chlorophyll a | 1.21 e | 2.67 e | 1.42 e | 14.04 d | 43.70 b | 41.21 b | 32.21 c | 51.23 a | 14.30 d | 1.13 e | 1.19 e | 1.17 e | |
| (μg cm−2) | (0.01) | (0.25) | (0.42) | (1.22) | (2.30) | (1.42) | (1.01) | (1.03) | (1.41) | (1.02) | (0.38) | (0.53) | |
| Survival rate | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |
| (%) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | |
| N | Zooxanthellae | 0.07 d | 0.13 d | 0.12 d | 0.08 d | 4.21 a | 2.30 b | 1.74 c | 4.30 a | 0.13 d | 0.17 d | 0.23 d | 0.17 d |
| (cell × 107 cm−2) | (0.01) | (0.01) | (0.02) | (0.02) | (0.05) | (0.03) | (0.01) | (0.04) | (0.02) | (0.02) | (0.02) | (0.03) | |
| Chlorophyll a | 1.09 f | 0.96 f | 0.95 f | 0.72 f | 42.33 b | 38.21 c | 19.21 d | 49.24 a | 10.03 e | 1.02 f | 1.01 f | 1.04 f | |
| (μg cm−2) | (0.02) | (0.04) | (0.02) | (0.01) | (1.21) | (1.23) | (1.73) | (2.31) | (1.20) | (0.01) | (0.01) | (0.01) | |
| Survival rate | 46.67 b | 53.33 b | 53.33 b | 73.00 ab | 100 a | 100 a | 73.33 a | 100 a | 73.33 ab | 46.67 b | 33.33 bc | 26.67 bc | |
| (%) | (11.55) | (11.55) | (11.55) | (11.55) | (0.00) | (0.00) | (11.55) | (0.00) | (11.55) | (11.55) | (11.55) | (11.55) | |
| Salinity and Temperature | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 psu | 30 psu | 35 psu | 40 psu | ||||||||||
| Feeding | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | 20 °C | 25 °C | 30 °C | |
| Y | Zooxanthellae | 0.00 | 0.71 g | 1.02 d | 0.00 | 2.13 a | 0.83 f | 0.00 | 2.22 a | 1.43 b | 0.00 | 0.93 e | 0.85 f |
| (cell × 107 cm−2) | (0.00) | (0.01) | (0.02) | (0.00) | (0.03) | (0.03) | (0.00) | (0.02) | (0.06) | (0.00) | (0.03) | (0.02) | |
| Chlorophyll a | 0.00 | 0.07 b | 0.02 c | 0.00 | 0.11 a | 0.05 b | 0.00 | 0.11 a | 0.09 ab | 0.00 | 0.08 ab | 0.07 ab | |
| (μg cm−2) | (0.00) | (0.01) | (0.01) | (0.00) | (0.01) | (0.03) | (0.00) | (0.02) | (0.01) | (0.00) | (0.01) | (0.03) | |
| Survival rate | 0.00 | 33.33 b | 20.00 b | 0.00 | 36.67 b | 33.33 b | 0.00 | 45.00 a | 30.00 b | 0.00 | 33.33 b | 25.00 b | |
| (%) | (0.00) | (5.77) | (10.00) | (0.00) | (5.77) | (5.77) | (0.00) | (5.00) | (10.00) | (0.00) | (5.77) | (5.00) | |
| N | Zooxanthellae | 0.00 | 0.53 b | 0.11 d | 0.00 | 0.54 b | 0.20 c | 0.00 | 1.20 a | 0.13 d | 0.00 | 0.11 d | 0.00 |
| (cell × 107 cm−2) | (0.00) | (0.01) | (0.01) | (0.00) | (0.05) | (0.03) | (0.00) | (0.04) | (0.02) | (0.00) | (0.01) | (0.00) | |
| Chlorophyll a | 0.00 | 0.04 ab | 0.05 ab | 0.00 | 0.04 ab | 0.04 ab | 0.00 | 0.06 a | 0.03 c | 0.00 | 0.05 ab | 0.00 | |
| (μg cm−2) | (0.00) | (0.01) | (0.01) | (0.00) | (0.01) | (0.01) | (0.00) | (0.01) | (0.01) | (0.00) | (0.01) | (0.00) | |
| Survival rate | 0.00 | 25.00 b | 8.33 d | 0.00 | 28.33 b | 15.00 c | 0.00 | 38.33 a | 40.00 a | 0.00 | 33.33 a | 0.00 | |
| (%) | (0.00) | (5.00) | (2.89) | (0.00) | (2.89) | (5.00) | (0.00) | (7.64) | (5.00) | (0.00) | (5.77) | (0.00) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, D.-S.; Wang, S.-H.; Sun, W.-T.; Liu, H.-L.; Pan, C.-H. The Effect of Feeding on Briareum violacea Growth, Survival and Larval Development under Temperature and Salinity Stress. Biology 2022, 11, 410. https://doi.org/10.3390/biology11030410
Ding D-S, Wang S-H, Sun W-T, Liu H-L, Pan C-H. The Effect of Feeding on Briareum violacea Growth, Survival and Larval Development under Temperature and Salinity Stress. Biology. 2022; 11(3):410. https://doi.org/10.3390/biology11030410
Chicago/Turabian StyleDing, De-Sing, Sheng-Hao Wang, Wei-Ting Sun, Huang-Lin Liu, and Chih-Hung Pan. 2022. "The Effect of Feeding on Briareum violacea Growth, Survival and Larval Development under Temperature and Salinity Stress" Biology 11, no. 3: 410. https://doi.org/10.3390/biology11030410
APA StyleDing, D.-S., Wang, S.-H., Sun, W.-T., Liu, H.-L., & Pan, C.-H. (2022). The Effect of Feeding on Briareum violacea Growth, Survival and Larval Development under Temperature and Salinity Stress. Biology, 11(3), 410. https://doi.org/10.3390/biology11030410