A Change in Conservation Status of Pachyphytum caesium (Crassulaceae), a Threatened Species from Central Mexico Based on Genetic Studies

 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
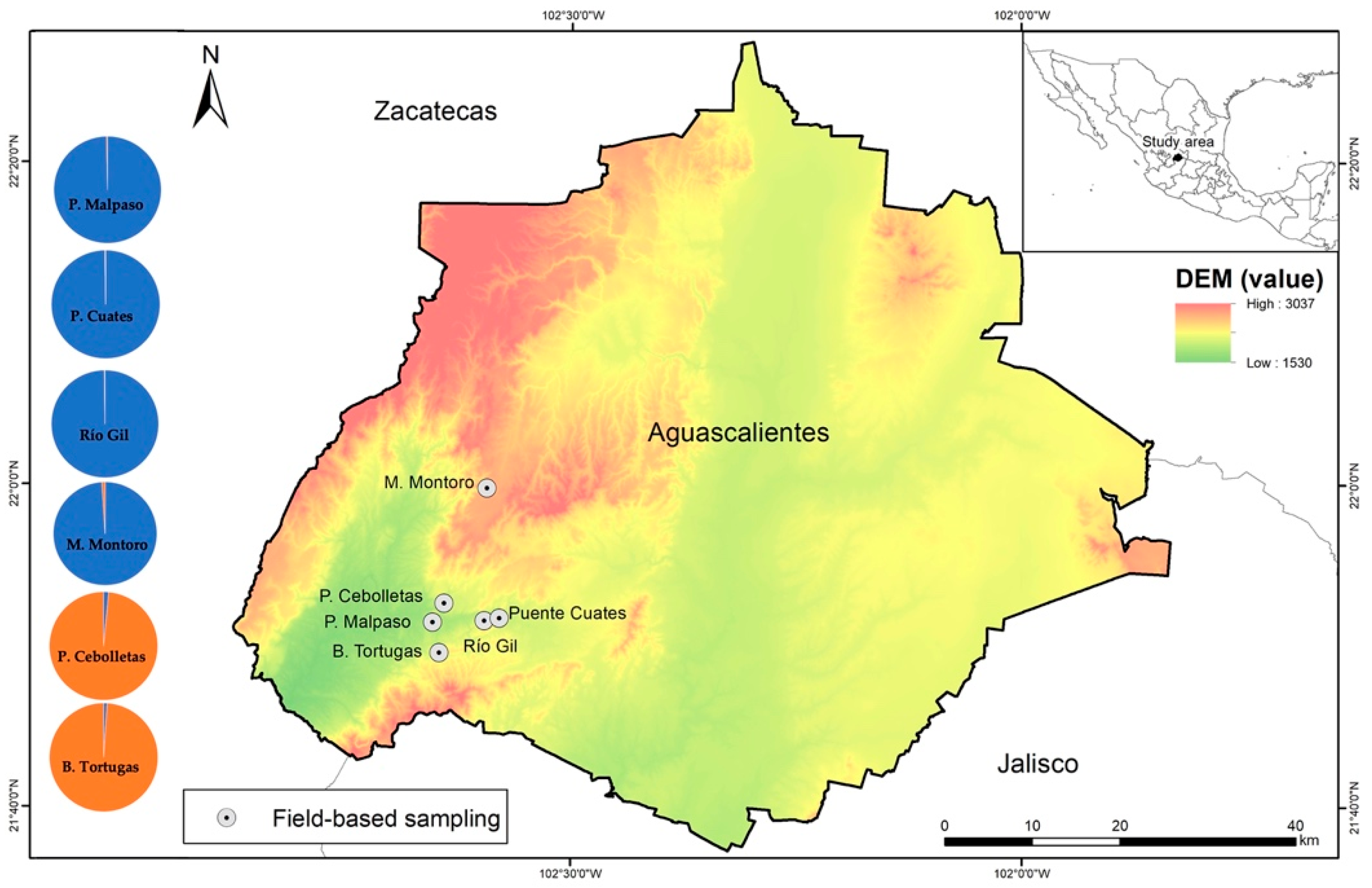
2.1. Study Area
2.2. Species Description
2.3. Biological Tissue Collection
2.4. Genetic Analysis
2.4.1. Allelic and Genetic Diversity
2.4.2. Population Structure and Isolation by Distance
2.4.3. Simulation to Obtain Genetic Diversity and Structure
2.5. Conservation Status
3. Results
3.1. Allelic and Genetic Diversity
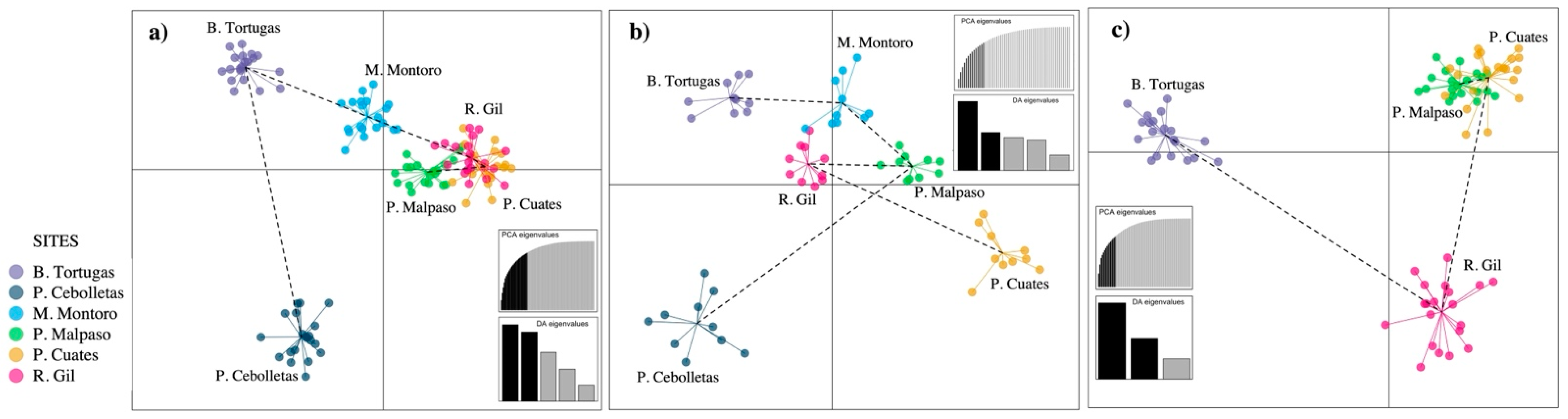
3.2. Population Structure and Isolation by Distance
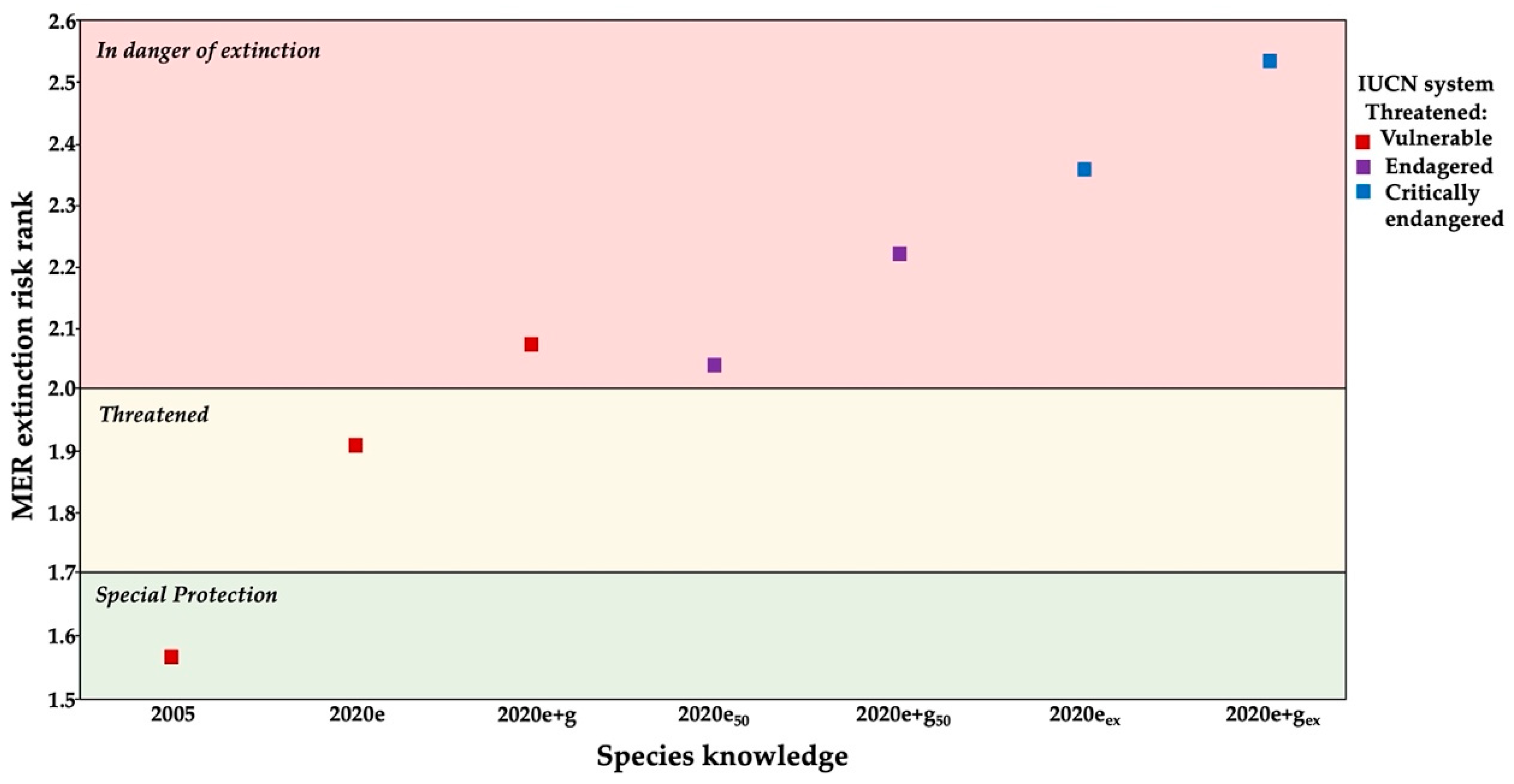
3.3. Conservation Status
4. Discussion
4.1. Allelic and Genetic Diversity
4.2. Population Structure and Isolation by Distance
4.3. Genetics Studies and Conservation Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Primack, R.B. A Primer of Conservation Biology; Sinauer Associates Inc.: Sunderland, MA, USA, 2012. [Google Scholar]
- Van Dyke, F.; Lamb, R.L. Conservation Biology Foundations, Concepts, Applications; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Pimm, S.L.; Joppa, L.N. How many plant species are there, where are they, and at what rate are they going extinct? Ann. Missouri Bot. Gard. 2015, 100, 170–176. [Google Scholar] [CrossRef]
- Médail, F.; Baumel, A. Using phylogeography to define conservation priorities: The case of narrow endemic plants in the Mediterranean Basin hotspot. Biol. Conserv. 2018, 224, 258–266. [Google Scholar] [CrossRef]
- Coker, O.M. Importance of genetics in conservation of biodiversity. Niger. J. Wildl. Manage. 2017, 1, 11–18. [Google Scholar]
- Hedrick, P.W. Genetics of Populations; Jones & Bartlett Learning: Burlington, MA, USA, 2011. [Google Scholar]
- Dunham, J.; Peacock, M.; Tracy, C.R.; Nielsen, J.; Vinyard, G. Assessing extinction risk: Integrating genetic information. Conserv. Ecol. 1999, 3. [Google Scholar] [CrossRef]
- Garner, B.A.; Hoban, S.; Luikart, G. IUCN Red List and the value of integrating genetics. Conserv. Genet. 2020, 21, 795–801. [Google Scholar] [CrossRef]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Ramírez, A.P.; Torres-Morán, M.I.; Molina-Moret, S.; Sánchez-González, J.D.J.; Santacruz-Ruvalcaba, F. Efficiency of RAPD, ISSR, AFLP and ISTR markers for the detection of polymorphisms and genetic relationships in camote de cerro (Dioscorea spp.). Electron. J. Biotechnol. 2014, 17, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Idrees, M.; Irshad, M. Molecular markers in plants for analysis of genetic diversity: A review. Eur. J. Acad. Res. 2014, 2, 1513–1540. [Google Scholar]
- Loera-Sánchez, M.; Studer, B.; Kölliker, R. DNA-based assessment of genetic diversity in grassland plant species: Challenges, approaches, and applications. Agronomy 2019, 9, 881. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.L.; Yorkson, M.; Morden, C. Population genetics of the endemic Hawaiian species Chrysodracon hawaiiensis and Chrysodracon auwahiensis (Asparagaceae): Insights from RAPD and ISSR variation. Int. J. Mol. Sci. 2016, 1, 1341. [Google Scholar] [CrossRef]
- Xia, T.; Chen, S.L.; Chen, S.Y.; Ge, X.J. Genetic variation within and among populations of Rhodiola alsia (Crassulaceae) native to the Tibetan Plateau as detected by ISSR markers. Biochem. Genet. 2005, 43, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Laikre, L.; Nilsson, T.; Primmer, C.R.; Ryman, N.; Allendorf, F.W. Importance of genetics in the interpretation of favourable conservation status. Conserv. Biol. 2009, 23, 1378–1381. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, L.C.; Souza, U.B.; Jardim, T.P.F.A.; Ballesteros–Mejia, L. Towards inclusion of genetic diversity measures into IUCN assessments: A case study on birds. Anim. Biodivers. Conserv. 2019, 42, 317–335. [Google Scholar] [CrossRef]
- Lanes, É.C.; Pope, N.S.; Alves, R.; Carvalho Filho, N.M.; Giannini, T.C.; Giulietti, A.M.; Imperatriz-Fonseca, V.L.; Monteiro, W.; Oliveira, G.; Silva, A.R.; et al. Landscape genomic conservation assessment of a narrow-endemic and a widespread morning glory from Amazonian savannas. Front. Plant Sci. 2018, 9, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1; IUCN: Gland, Switzerland; Cambridge, UK, 2012. [Google Scholar]
- Clark-Tapia, R.; Alfonso-Corrado, C.; Eguiarte, L.E.; Molina-Freaner, F. Clonal diversity and distribution in Stenocereus eruca (Cactaceae), a narrow endemic cactus of the Sonoran Desert. Am. J. Bot. 2005, 92, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Pingarroni, A.; Molina-Garay, C.; Rosas-Osorio, C.; Alfonso-Corrado, C.; Clark-Tapia, R.; Monsalvo-Reyes, A.; Campos, J.E. Abundancia y diversidad genética de Quercus mulleri, especie mocroendémica amenazada de Oaxaca. Madera Bosques 2020, 26, e2611782. [Google Scholar] [CrossRef]
- Clark-Tapia, R.; González-Adame, G.; Campos, J.E.; Aguirre-Hidalgo, V.; Pacheco-Cruz, N.; Von Thaden, J.J.; Campista-Léon, S.; Peinado-Guevara, L.I.; Alfonso-Corrado, C. Effects of habitats loss on the ecology of Pachyphytum caesium (Crassulaceae), a specialized cliff-dwelling endemic species in central Mexico. Diversity 2021, 13, 421. [Google Scholar] [CrossRef]
- Ma, Y.P.; Chen, G.; Grumbine, R.E.; Dao, Z.; Sun, W.; Guo, H. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Yang, J.; Gao, Z.; Sun, W.; Zhang, C. High regional genetic differentiation of an endangered relict plant Craigia yunnanensis and implications for its conservation. Plant Divers. 2016, 38, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Trejo, I.; Dirzo, R. Deforestation of seasonally dry tropical forest: A national and local analysis in Mexico. Biol. Conserv. 2000, 94, 133–142. [Google Scholar] [CrossRef]
- Siqueiros-Delgado, M.E.; Rodríguez-Avalos, J.A.; Martínez-Ramírez, J.; Sierra-Muñoz, J.C. Situación actual de la vegetación del estado de Aguascalientes México. Bot. Sci. 2016, 94, 455–470. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Calix, E. Crassulaceae Flora de Bajío y de Regiones Adyacentes; Instituto de Ecología A.C.: Michoacán, Mexico, 2008. [Google Scholar]
- Kimnach, M.; Moran, R. Pachyphytum caesium a new species from Aguascalientes México. Cactus Succul. J. 1993, 65, 59–62. [Google Scholar]
- Meyrán-García, J.; López-Chávez, L. Las Crasuláceas de México; Sociedad Mexicana de Cactología A.C.: Distrito Federal, Mexico, 2003. [Google Scholar]
- Etter, J.; Kristen, M. Mexican Cliff-dwellers. Cactus Succul. J. 2006, 78, 251–260. [Google Scholar] [CrossRef]
- SEMARNAT. Norma Oficial Mexicana NOM-059-SEMARNAT-2010; Protección Ambiental-Especies Nativas de México de Flora y Fauna Silvestres-Categorías de Riesgo y Especificaciones para su Inclusión, Exclusión o Cambio-Lista de Especies en Riesgo; Semarnat: Distrito Federal, Mexico, 2010; Available online: https://dof.gob.mx/nota_detalle_popup.php?codigo=5173091 (accessed on 21 November 2021).
- Arroyo, T.P.F.; Olson, M.E.; García-Mendoza, A.B.I.S.A.Í.; Solano, E. A GIS-based comparison of the Mexican national and IUCN methods for determining extinction risk. Conserv. Biol. 2009, 23, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Estadística, Geografía e Informática (INEGI)-Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO)-Instituto Nacional de Ecología (INE). Ecorregiones Terr. México; 2008. Available online: http://geoportal.conabio.gob.mx/metadatos/doc/html/ecort08gw.html (accessed on 2 January 2022).
- Wu, W.; Chen, F.; Yeh, K.; Chen, J. ISSR analysis of genetic diversity and structure of plum varieties cultivated in southern China. Biology 2019, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meirmans, P.G. Genodive version 3.0: Easy-to-use software for the analysis of genetic data of diploids and polyploids. Mol. Ecol. Resour. 2020, 20, 1126–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, M.; Rousset, F. An exact test for population differentiation. Evolution 1995, 49, 1280–1283. [Google Scholar] [CrossRef]
- Lynch, M.; Milligan, B.G. Analysis of population genetic structure with RAPD markers. Mol. Ecol. 1994, 3, 91–99. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.501: Genetic analysis in excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Kamvar, N.K.; Brooks, J.C.; Grünwald, N.J. Novel R tools for the analysis of genome-wide population genetic data with emphasis on clonality. Front. Genet. 2015, 6, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 10 April 2021).
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crow, J.F.; Aoki, K. Group selection for a polygenic behavioral trait: Estimating the degree of population subdivision. Proc Natl. Acad. Sci. USA 1984, 81, 6073–6077. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; Vonholdt, B.M. Structure harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Slatkin, M. A measure of population subdivision based on microsatellite allele frequencies. Genetics 1995, 139, 457–462. [Google Scholar] [CrossRef]
- Austin, J.D.; Jordan, F. Population genetic structure and conservation genetics of threatened Okaloosa darters (Etheostoma okaloosae). Conserv. Genet. 2011, 12, 981–989. [Google Scholar] [CrossRef]
- Szczecinska, M.; Sramko, G.; Wolosz, K.; Sawicki, J. Genetic diversity and population structure of rare and endangered plant species Pulsatilla patens (L.) Mill in East Central Europe. PLoS ONE 2016, 11, e0151730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.Y.; López-Pujol, J.; Chung, M.G. Comparative genetic structure between Sedum ussuriense and S. kamtschaticum (Crassulaceae), two stonecrops co-occurring on rocky cliffs. Am. J. Bot. 2014, 101, 946–956. [Google Scholar] [CrossRef]
- Chung, M.Y.; López-Pujol, J.; Chung, M.G. Population genetic structure of Sedum polytrichoides (Crassulaceae): Insights into barriers to gene flow. Korean J. Plant Taxon. 2016, 46, 361–370. [Google Scholar] [CrossRef]
- György, Z.; Fjelldal, E.; Ladányi, M.; Aspholm, P.E. Genetic diversity of roseroot (Rhodiola rosea) in North-Norway. Biochem. Syst. Ecol. 2013, 50, 361–367. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W. Allozyme diversity in plant species. In Plant Population Genetics, Breeding, and Genetic Resources; Brown, A.H.D., Clegg, M.T., Kahler, A.L., Weir, B.S., Eds.; Sinauer: Sunderland, MA, USA, 1990; pp. 43–63. [Google Scholar]
- Leimu, R.; Mutikainen, P.I.A.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Plenk, K.; Bardy, K.; Höhn, M.; Kropf, M. Long-term survival and successful conservation? Low genetic diversity but no evidence for reduced reproductive success at the north-westernmost range edge of Poa badensis (Poaceae) in Central Europe. Biodivers. Conserv. 2019, 28, 1245–1265. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, B.D.; Charlesworth, N.H.; Barton, N.H. The effects of genetic and geographic structure on neutral variation. Ann. Rev. Ecol. Evol. Syst. 2003, 34, 99–125. [Google Scholar] [CrossRef]
- Honnay, O.; Jacquemyn, H. Susceptibility of common and rare plant species to the genetic consequences of habitat fragmentation. Conserv. Biol. 2007, 3, 823–831. [Google Scholar] [CrossRef]
- Qiao, Q.; Zhang, C.; Milne, R.I. Population’s genetics and breeding system of Tupistra pingbianensis (Liliaceae), a naturally rare plant endemic to SW China. J. Syst. Evol. 2010, 48, 47–57. [Google Scholar] [CrossRef]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogen. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef]
- Kimura, M.; Crow, J.F. The number of alleles that can be maintained in a finite population. Genetics 1964, 49, 725–738. [Google Scholar] [CrossRef]
- McCommas, S.A.; Bryant, E.H. Loss of electrophoretic variation in serially bottlenecked populations. Heredity 1990, 64, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogler, F.; Reisch, C. Genetic variation on the rocks- the impact of climbing on the population ecology of a typical cliff plant. J. Appl. Ecol. 2011, 48, 899–905. [Google Scholar] [CrossRef]
- Ge, X.J.; Zhang, L.B.; Yuan, Y.M.; Hao, G.; Chang, T.Y. Strong genetic differentiation of the East-Himalayan Megacodon stylophorus (Gentianaceae) detected by inter-simple sequence repeats (ISSR). Biodivers. Conserv. 2005, 14, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Klug, W.S.; Cummings, M.R.; Spencer, C.A.; Killian, D.J. Concepts of Genetics; Pearson: London, UK, 2020. [Google Scholar]
- Hou, Y.; Lou, A. Population genetic diversity and structure of a naturally isolated plant species, Rhodiola dumulosa (Crassulaceae). PLoS ONE 2011, 6, e24497. [Google Scholar] [CrossRef] [Green Version]
- Raj, J.X.; Srivastara, R.B. Using Inter-simple sequence repeat markers to analyze the genetic structure of natural Rhodiola imbricata populations from the trans-Himalayan region and implications for species conservation. Madras Agric. J. 2012, 99, 221–227. [Google Scholar]
- Gentili, R.; Fenu, G.; Mattana, E.; Citterio, S.; De Mattia, F.; Bacchetta, G. Conservation genetics of two island endemic Ribes spp. (Grossulariaceae) of Sardinia: Survival or extinction? Plant Biol. 2015, 17, 1085–1094. [Google Scholar] [CrossRef]
- Lombaert, E.; Guillemaud, T.; Deleury, E. Biases of STRUCTURE software when exploring introduction routes of invasive species. Heredity 2018, 120, 485–499. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.E.; Purvis, A.; Gittleman, J.L. Biological correlates of extinction risk in bats. Am. Nat. 2003, 161, 601–614. [Google Scholar] [CrossRef]
- Laikre, L. Genetic diversity is overlooked in international conservation policy implementation. Conserv. Genet. 2010, 11, 349–354. [Google Scholar] [CrossRef]
- Hedrick, P.W.; Garcia–Dorado, A. Understanding inbreeding depression, purging, and genetic rescue. Trends Ecol. Evol. 2016, 31, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Efective population size/adult population size ratios in wildlife: A review. Genet. Res. 1995, 66, 95–107. [Google Scholar] [CrossRef]
- Weiser, E.L.; Grueber, C.E.; Jamieson, I.G. Simulating retention of rare alleles in small populations to assess management options for species with different life histories. Conserv. Biol. 2013, 27, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Caujapé-Castells, J.; Pedrola-Monfort, J. Designing ex-situ conservation strategies through the assessment of neutral genetic 759 markers: Application to the endangered Androcymbium gramineum. Conserv. Genet. 2004, 5, 131–144. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-León, T.; Clark-Tapia, R.; Campos, J.E.; Peinado-Guevara, L.I.; Campista-León, S.; Molina-Freaner, F.; Pacheco-Cruz, N.; González-Adame, G.; Von Thaden Ugalde, J.J.; Alfonso-Corrado, C. A Change in Conservation Status of Pachyphytum caesium (Crassulaceae), a Threatened Species from Central Mexico Based on Genetic Studies. Biology 2022, 11, 379. https://doi.org/10.3390/biology11030379
Martínez-León T, Clark-Tapia R, Campos JE, Peinado-Guevara LI, Campista-León S, Molina-Freaner F, Pacheco-Cruz N, González-Adame G, Von Thaden Ugalde JJ, Alfonso-Corrado C. A Change in Conservation Status of Pachyphytum caesium (Crassulaceae), a Threatened Species from Central Mexico Based on Genetic Studies. Biology. 2022; 11(3):379. https://doi.org/10.3390/biology11030379
Chicago/Turabian StyleMartínez-León, Tania, Ricardo Clark-Tapia, Jorge E. Campos, Luz Isela Peinado-Guevara, Samuel Campista-León, Francisco Molina-Freaner, Nelly Pacheco-Cruz, Gabriel González-Adame, Juan José Von Thaden Ugalde, and Cecilia Alfonso-Corrado. 2022. "A Change in Conservation Status of Pachyphytum caesium (Crassulaceae), a Threatened Species from Central Mexico Based on Genetic Studies" Biology 11, no. 3: 379. https://doi.org/10.3390/biology11030379
APA StyleMartínez-León, T., Clark-Tapia, R., Campos, J. E., Peinado-Guevara, L. I., Campista-León, S., Molina-Freaner, F., Pacheco-Cruz, N., González-Adame, G., Von Thaden Ugalde, J. J., & Alfonso-Corrado, C. (2022). A Change in Conservation Status of Pachyphytum caesium (Crassulaceae), a Threatened Species from Central Mexico Based on Genetic Studies. Biology, 11(3), 379. https://doi.org/10.3390/biology11030379