


Changes in the Suitable Habitats of Three Endemic Fishes to Climate Change in Tibet


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Climatic Variables
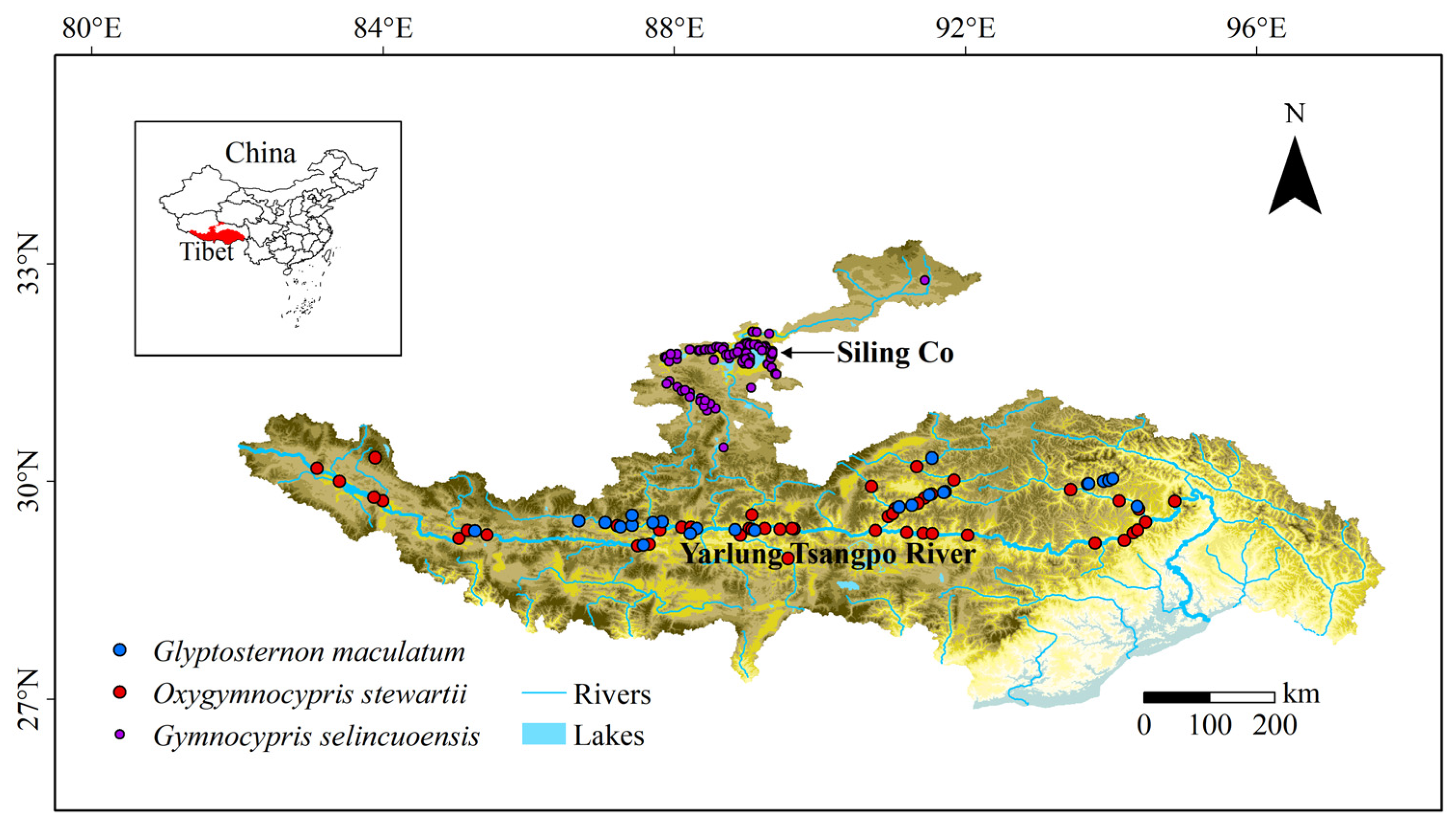
2.2. Occurrence Records
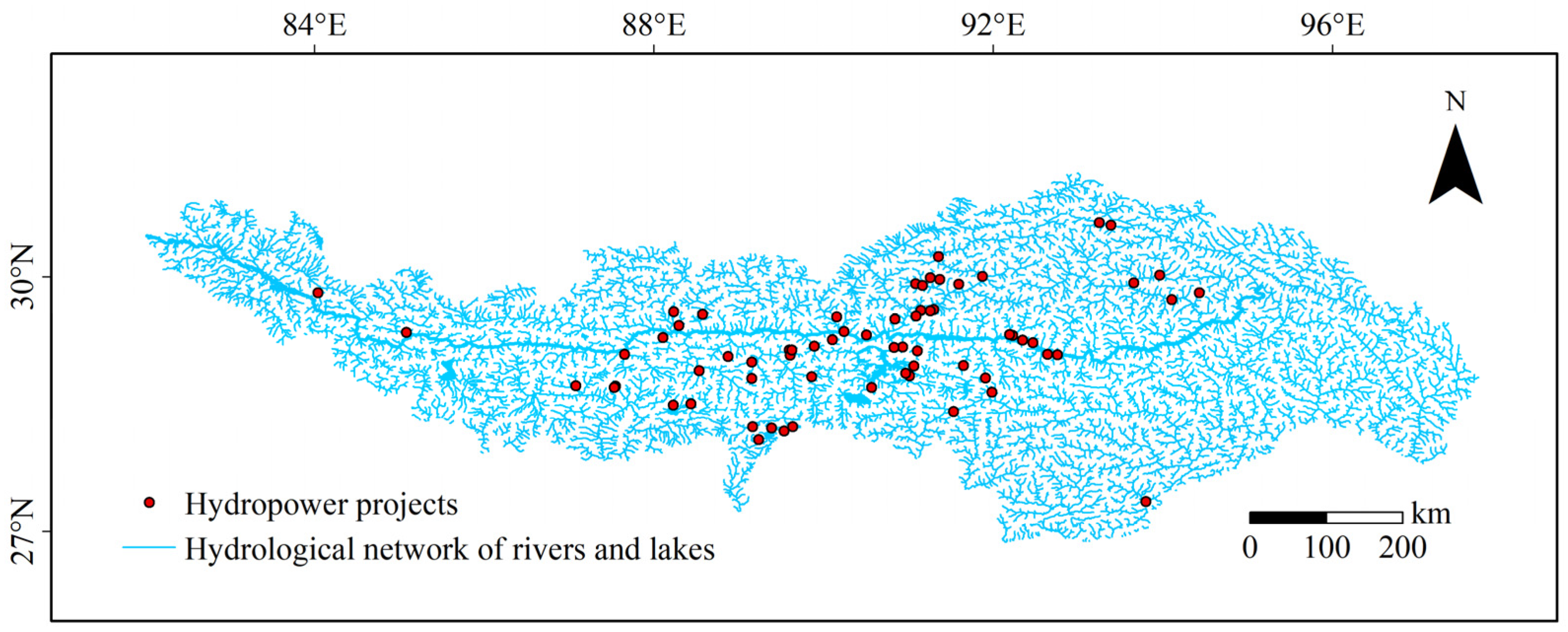
2.3. Hydropower Projects Records
2.4. MaxEnt Modeling
2.5. Processing of Model Results
3. Results
3.1. Model Evaluation and Importance of Environmental Variables
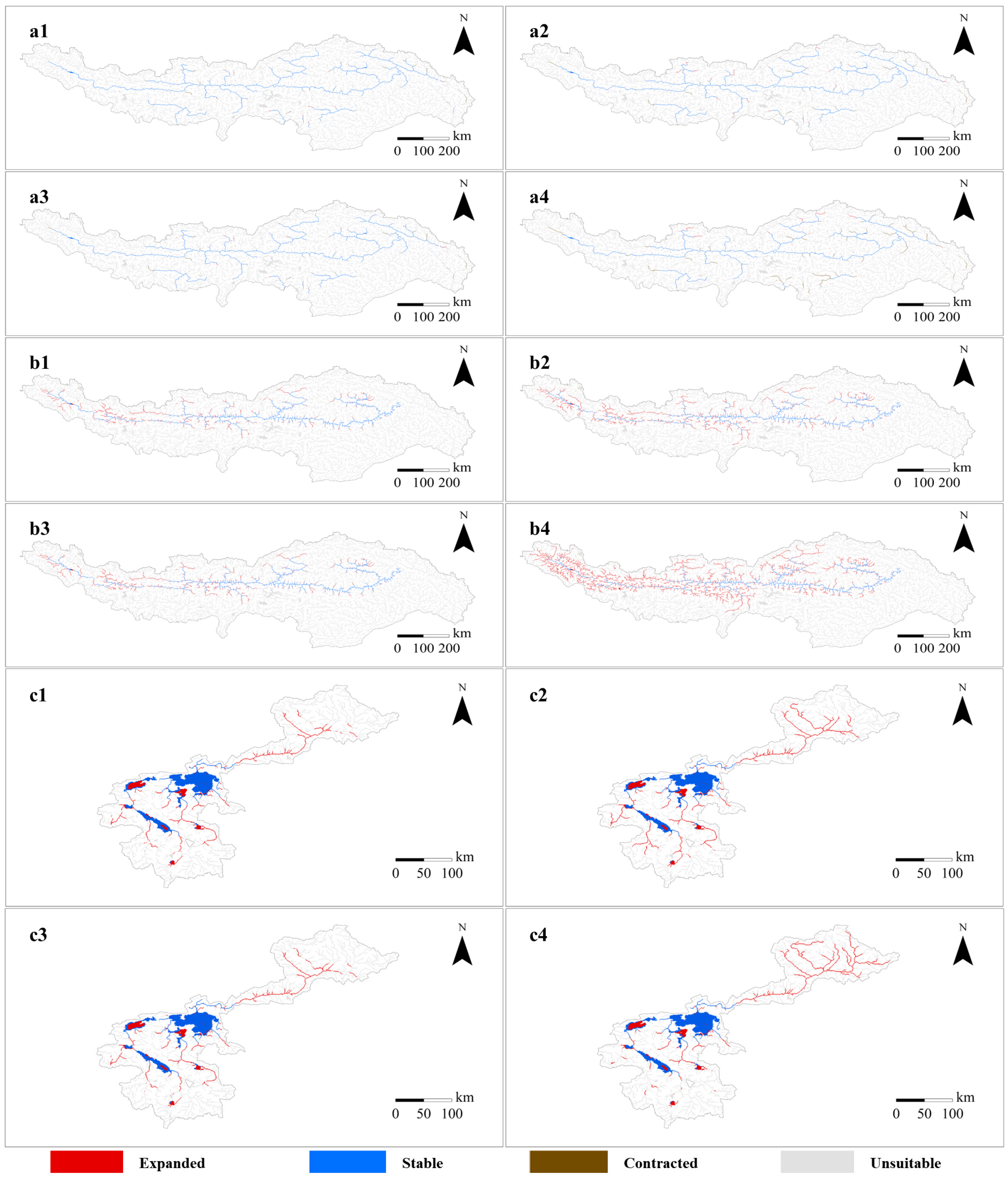
3.2. Changes of Fish Habitats to Climate Change
3.3. Changes in Average Elevation of Fish Suitable Habitats to Climate Change
3.4. Impact of River Fragmentation on Fish Caused by Hydropower Projects Construction
4. Discussion
4.1. Impact of Climate Change on the Range of Suitable Habitats of Three Species
4.2. Impact of Climate Change on Migration of Suitable Habitats of Three Species
4.3. Impact of Hydropower Projects on Changes of Suitable Habitats of Three Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, S.; Xu, Y.; You, Q.; Flügel, W.-A.; Pepin, N.; Yao, T. Review of climate and cryospheric change in the Tibetan Plateau. Environ. Res. Lett. 2010, 5, 015101. [Google Scholar] [CrossRef]
- Tao, J.; Chen, Y.; He, D.; Ding, C. Relationships between climate and growth of Gymnocypris selincuoensis in the Tibetan Plateau. Ecol. Evol. 2015, 5, 1693–1701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yao, T.; Xie, H.; Zhang, K.; Zhu, F. Lakes’ state and abundance across the Tibetan Plateau. Chin. Sci. Bull. 2014, 59, 3010–3021. [Google Scholar] [CrossRef]
- Cuo, L.; Zhang, Y.; Zhu, F.; Liang, L. Characteristics and changes of streamflow on the Tibetan Plateau: A review. J. Hydrol. Reg. Stud. 2014, 2, 49–68. [Google Scholar] [CrossRef]
- Comte, L.; Grenouillet, G. Do stream fish track climate change? Assessing distribution shifts in recent decades. Ecography 2013, 36, 1236–1246. [Google Scholar] [CrossRef]
- Markovic, D.; Carrizo, S.; Freyhof, J.; Cid, N.; Lengyel, S.; Scholz, M.; Kasperdius, H.; Darwall, W. Europe’s freshwater biodiversity under climate change: Distribution shifts and conservation needs. Divers. Distrib. 2014, 20, 1097–1107. [Google Scholar] [CrossRef]
- Tao, J.; He, D.; Kennard, M.J.; Ding, C.; Bunn, S.E.; Liu, C.; Jia, Y.; Che, R.; Chen, Y. Strong evidence for changing fish reproductive phenology under climate warming on the Tibetan Plateau. Glob. Chang. Biol. 2018, 24, 2093–2104. [Google Scholar] [CrossRef]
- Holt, R.E.; Jørgensen, C. Climate change in fish: Effects of respiratory constraints on optimal life history and behaviour. Biol. Lett. 2015, 11, 20141032. [Google Scholar] [CrossRef]
- Comte, L.; Grenouillet, G. Distribution shifts of freshwater fish under a variable climate: Comparing climatic, bioclimatic and biotic velocities. Divers. Distrib. 2015, 21, 1014–1026. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Herrera, R.G.; Oberdorff, T.; Anderson, E.P.; Brosse, S.; Carvajal-Vallejos, F.M.; Frederico, R.G.; Hidalgo, M.; Jezequel, C.; Maldonado, M.; Maldonado-Ocampo, J.A.; et al. The combined effects of climate change and river fragmentation on the distribution of Andean Amazon fishes. Glob. Chang. Biol. 2020, 26, 5509–5523. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Radinger, J.; Hölker, F.; Horký, P.; Slavík, O.; Dendoncker, N.; Wolter, C. Synergistic and antagonistic interactions of future land use and climate change on river fish assemblages. Glob. Chang. Biol. 2016, 22, 1505–1522. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. Going with the flow: Global warming and the challenge of sustaining river ecosystems in monsoonal Asia. Water Sci. Technol. Water Supply 2007, 7, 69–80. [Google Scholar] [CrossRef]
- Carvajal-Quintero, J.D.; Januchowski-Hartley, S.R.; Maldonado-Ocampo, J.A.; Jézéquel, C.; Delgado, J.; Tedesco, P.A. Damming fragments species’ ranges and heightens extinction risk. Conserv. Lett. 2017, 10, 708–716. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef]
- Gibson-Reinemer, D.K.; Rahel, F.J.; Albeke, S.E.; Fitzpatrick, R.M. Natural and anthropogenic barriers to climate tracking in river fishes along a mountain–plains transition zone. Divers. Distrib. 2017, 23, 761–770. [Google Scholar] [CrossRef]
- Sun, H.; He, D.; Sui, X.; Chen, Y. Predicting impacts of future climate change and hydropower development towards habitats of native and non-native fishes. Sci. Total Environ. 2020, 707, 135419. [Google Scholar] [CrossRef]
- Radinger, J.; Essl, F.; Hölker, F.; Horký, P.; Slavík, O.; Wolter, C. The future distribution of river fish: The complex interplay of climate and land use changes, species dispersal and movement barriers. Glob. Chang. Biol. 2017, 23, 4970–4986. [Google Scholar] [CrossRef]
- Araújo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef]
- Li, X.; Tian, H.; Wang, Y.; Li, R.; Song, Z.; Zhang, F.; Xu, M.; Li, D. Vulnerability of 208 endemic or endangered species in China to the effects of climate change. Reg. Environ. Chang. 2013, 13, 843–852. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, X.; Jia, Y.; Chen, Y.; He, D. Predicting fish invasions in the Yarlung Tsangpo River of the Qinghai-Tibet Plateau, China. Am. Fish. Soc. Symp. 2016, 84, 139–167. [Google Scholar]
- Hansen, G.J.; Read, J.S.; Hansen, J.F.; Winslow, L.A. Projected shifts in fish species dominance in Wisconsin lakes under climate change. Glob. Chang. Biol. 2017, 23, 1463–1476. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; He, D.; Jia, Y.; Sun, H.; Chen, Y. Phylogeographic studies of schizothoracine fishes on the central Qinghai-Tibet Plateau reveal the highest known glacial microrefugia. Sci. Rep. 2017, 7, 10983. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sui, X.; Lü, Z.; Chen, Y. A prediction of the global habitat of two invasive fishes (Pseudorasbora parvaandCarassius auratus) from East Asia using Maxent. Biodivers. Sci. 2014, 22, 182–188. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Reddy, M.; Begum, H.; Sunil, N.; Pandravada, S.; Sivaraj, N.; Kumar, S. Mapping the climate suitability using MaxEnt modeling approach for Ceylon spinach (Basella alba L.) cultivation in India. J. Agric. Sci. 2015, 10, 87–97. [Google Scholar] [CrossRef]
- Gao, X.; Huang, Y.; Zheng, J.; Xiao, J.; Wang, H. Impact of meteorological and geographical factors on the distribution of leishmaniasis’s vector in mainland China. Pest Manag. Sci. 2020, 76, 961–966. [Google Scholar] [CrossRef] [PubMed]
- National Forestry and Grassland Administration. List of National Key Protected Wild Animals. Chin. J. Wildl. 2021, 42, 36. [Google Scholar]
- Yue, P.; Shan, X.; Lin, R. Fauna Sinica, Osteichthyes, Cypriniformes III; China Scientific Book Services Co., Ltd.: Beijing, China, 2000. [Google Scholar]
- Ng, H. The IUCN Red List of Threatened Species 2010: e.T168408A6487005. Available online: https://dx.doi.org/10.2305/IUCN.UK.2010-4.RLTS.T168408A6487005.en (accessed on 27 May 2022).
- Chen, Y. Systematics and Resource Biology of the Schizothoracine Fishes. Ph.D. Dissertation, Chinese Academy of Science, Wuhan, China, 2000. (In Chinese). [Google Scholar]
- Domisch, S.; Amatulli, G.; Jetz, W. Near-global freshwater-specific environmental variables for biodiversity analyses in 1 km resolution. Sci. Data 2015, 2, 150073. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of a problematic alien invasive species—The American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Brzozowski, M.; Pełechaty, M.; Bogawski, P. A winner or a loser in climate change? Modelling the past, current, and future potential distributions of a rare charophyte species. Glob. Ecol. Conserv. 2022, 34, e02038. [Google Scholar] [CrossRef]
- Fekrat, L.; Farashi, A. Impacts of climatic changes on the worldwide potential geographical dispersal range of the leopard moth, Zeuzera pyrina (L.) (Lepidoptera: Cossidae). Glob. Ecol. Conserv. 2022, 34, e02050. [Google Scholar] [CrossRef]
- Zhang, J.; Lun, Y.; Liu, l.; Liu, Y.; Li, X.; Xu, Z. CMIP6 evaluation and projection of climate change in Tibetan Plateau. J. Beijing Norm. Univ. (Nat. Sci.) 2022, 58, 77–89. [Google Scholar]
- Xu, J.; Zhang, X.; Zhang, W.; Hou, N.; Feng, C.; Yang, S.; Jia, K.; Yao, Y.; Xie, X.; Jiang, B.; et al. Assessment of surface downward longwave radiation in CMIP6 with comparison to observations and CMIP5. Atmos. Res. 2022, 270, 106056. [Google Scholar] [CrossRef]
- Shang, W.; Duan, K.; Li, S.; Ren, X.; Huang, B. Simulation of the dipole pattern of summer precipitation over the Tibetan Plateau by CMIP6 models. Environ. Res. Lett. 2021, 16, 014047. [Google Scholar] [CrossRef]
- Song, C.; Fan, C.; Zhu, J.; Wang, J.; Sheng, Y.; Liu, K.; Chen, T.; Zhan, P.; Luo, S.; Yuan, C. A comprehensive geospatial database of nearly 100 000 reservoirs in China. Earth Syst. Sci. Data 2022, 14, 4017–4034. [Google Scholar] [CrossRef]
- Kano, Y.; Dudgeon, D.; Nam, S.; Samejima, H.; Watanabe, K.; Grudpan, C.; Grudpan, J.; Magtoon, W.; Musikasinthorn, P.; Nguyen, P.T. Impacts of dams and global warming on fish biodiversity in the Indo-Burma hotspot. PLoS ONE 2016, 11, e0160151. [Google Scholar] [CrossRef]
- Varela, S.; Anderson, R.P.; García-Valdés, R.; Fernández-González, F. Environmental filters reduce the effects of sampling bias and improve predictions of ecological niche models. Ecography 2014, 37, 1084–1091. [Google Scholar] [CrossRef]
- Tesfamariam, B.G.; Gessesse, B.; Melgani, F. MaxEnt-based modeling of suitable habitat for rehabilitation of Podocarpus forest at landscape-scale. Environ. Syst. Res. 2022, 11, 4. [Google Scholar] [CrossRef]
- Zhang, Y.; Hughes, A.C.; Zhao, Z.; Li, Z.; Qin, Y. Including climate change to predict the global suitable area of an invasive pest: Bactrocera correcta (Diptera: Tephritidae). Glob. Ecol. Conserv. 2022, 34, e02021. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Zhang, C. Fish Diversity Patterns in the Mekong River Basin and Its Respense to Climate Change and River Fragmentation. Institute of Hydrobiology; Chinese Academy of Science: Wuhan, China, 2021. [Google Scholar]
- Chu, X.; Zheng, B.; Dai, D. Fauna Sinica Osteichthyes: Siluriformes; China Scientific Book Services Co., Ltd.: Beijing, China, 1999. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- You, Q.; Kang, S.; Wu, Y.; Yan, Y. Climate change over the Yarlung Tsangpo river basin during 1961–2005. J. Geogr. Sci. 2007, 17, 409–420. [Google Scholar] [CrossRef]
- Duan, Z.; Wang, M.; Chang, X.; Gao, W. Response of river-lake hydrologic regimes to local climate change in the Yunnan-Guizhou Plateau region, China. Reg. Environ. Chang. 2020, 20, 122. [Google Scholar] [CrossRef]
- Pacifici, M.; Foden, W.B.; Visconti, P.; Watson, J.E.; Butchart, S.H.; Kovacs, K.M.; Scheffers, B.R.; Hole, D.G.; Martin, T.G.; Akçakaya, H.R. Assessing species vulnerability to climate change. Nat. Clim. Chang. 2015, 5, 215–224. [Google Scholar] [CrossRef]
- Whitney, J.E.; Whittier, J.B.; Paukert, C.P.; Olden, J.D.; Strecker, A.L. Forecasted range shifts of arid-land fishes in response to climate change. Rev. Fish Biol. Fish. 2017, 27, 463–479. [Google Scholar] [CrossRef]
- Stocker, T. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Tibet Fisheries Bureau. Fishes and Fish Resources in Xizang, China; China Agriculture Press: Beijing, China, 1995. [Google Scholar]
- Huo, B. Study on the Biology and Population Dynamics of Oxygymnocypris stewartii. Doctoral Dissertation, Huazhong Agriculture University, Wuhan, China, 2014. [Google Scholar]
- Winemiller, K.O.; McIntyre, P.B.; Castello, L.; Fluet-Chouinard, E.; Giarrizzo, T.; Nam, S.; Baird, I.G.; Darwall, W.; Lujan, N.K.; Harrison, I. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 2016, 351, 128–129. [Google Scholar] [CrossRef]
- Comte, L.; Murienne, J.; Grenouillet, G. Species traits and phylogenetic conservatism of climate-induced range shifts in stream fishes. Nat. Commun. 2014, 5, 5053. [Google Scholar] [CrossRef]
- Chevalier, M.; Comte, L.; Laffaille, P.; Grenouillet, G. Interactions between species attributes explain population dynamics in stream fishes under changing climate. Ecosphere 2018, 9, e02061. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Environmental Variables | Contribution (%) | Permutation Importance |
|---|---|---|---|
| Glyptosternon maculatum | Flow accumulation | 81.4 | 77.3 |
| Mean temperature of driest quarter (bio9) | 10.1 | 9.1 | |
| Precipitation of coldest quarter (bio19) | 5.5 | 5.5 | |
| Precipitation of wettest month(bio13) | 1.4 | 7.3 | |
| Slope | 1.2 | 1.2 | |
| Isothermality (bio3) | 0.4 | 0.4 | |
| Oxygymnocypris stewartii | Flow length | 66.3 | 32.6 |
| Max temperature of warmest month (bio5) | 9.2 | 36.2 | |
| Mean temperature of wettest quarter (bio8) | 8.7 | 11.6 | |
| Mean temperature of driest quarter (bio9) | 7.5 | 12.8 | |
| Slope | 5.4 | 6.7 | |
| Precipitation seasonality (bio15) | 3 | 0.2 | |
| Gymnocypris selincuoensis | Flow length | 66.5 | 34.3 |
| Isothermality (bio3) | 12.7 | 21.9 | |
| Max temperature of warmest month (bio5) | 6.9 | 32.7 | |
| Slope | 6.8 | 6 | |
| Temperature seasonality (bio4) | 5.5 | 3.6 | |
| Elevation | 1.6 | 1.4 |
| Scenarios | Glyptosternon Maculatum | Oxygymnocypris Stewartia | Gymnocypris Selincuoensis | |||||
|---|---|---|---|---|---|---|---|---|
| Habitat Gain | Habitat Loss | Habitat Net Change | Habitat Gain | Habitat Loss | Habitat Net Change | Habitat Gain (Habitat Net Change) | Habitat Loss | |
| 2050-RCP2.6 | 204 (3.5%) 1 | 318 (5.5%) | −114 (2.0%) | 2788 (60.9%) | 41 (1.0%) | 2747 (60.0%) | 1778 (46.4%) | 0 |
| 2050-RCP2.6-Dam | 200 | 0 | 110 | 2705 | 0 | 2664 | – | – |
| 2050-RCP8.5 | 383 (6.6%) | 628 (10.9%) | −245 (4.2%) | 4446 (97.1%) | 39 (1.0%) | 4407 (96.2%) | 2157 (56.3%) | 0 |
| 2050-RCP8.5-Dam | 352 | 0 | 214 | 4250 | 0 | 4211 | – | – |
| 2090-RCP2.6 | 206 (3.6%) | 417 (7.2%) | −211 (3.7%) | 3231 (70.6%) | 32 (1.0%) | 3199 (69.9%) | 1819 (47.5%) | 0 |
| 2090-RCP2.6-Dam | 201 | 0 | 206 | 3112 | 0 | 3080 | – | – |
| 2090-RCP8.5 | 365 (6.3%) | 1453 (25.1%) | −1088 (18.8%) | 10949 (239.1%) | 39 (1.0%) | 10910 (238.3%) | 2180 (56.9%) | 0 |
| 2090-RCP8.5-Dam | 365 | 0 | −1088 | 9885 | 0 | 9846 | – | – |
| Scenarios | Average Elevation of Habitats (m) | ||
|---|---|---|---|
| Glyptosternon maculatum | Oxygymnocypris stewartia | Gymnocypris selincuoensis | |
| Current | 4891 | 4838 | 4800 |
| 2050-RCP2.6 | 4909 | 4874 | 4847 |
| 2050-RCP8.5 | 4928 | 4872 | 4863 |
| 2090-RCP2.6 | 4914 | 4867 | 4848 |
| 2090-RCP8.5 | 4948 | 4922 | 4873 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, T.; He, D.; Zhu, R.; Sui, X.; Chen, Y. Changes in the Suitable Habitats of Three Endemic Fishes to Climate Change in Tibet. Biology 2022, 11, 1808. https://doi.org/10.3390/biology11121808
Mu T, He D, Zhu R, Sui X, Chen Y. Changes in the Suitable Habitats of Three Endemic Fishes to Climate Change in Tibet. Biology. 2022; 11(12):1808. https://doi.org/10.3390/biology11121808
Chicago/Turabian StyleMu, Tong, Dekui He, Ren Zhu, Xiaoyun Sui, and Yifeng Chen. 2022. "Changes in the Suitable Habitats of Three Endemic Fishes to Climate Change in Tibet" Biology 11, no. 12: 1808. https://doi.org/10.3390/biology11121808
APA StyleMu, T., He, D., Zhu, R., Sui, X., & Chen, Y. (2022). Changes in the Suitable Habitats of Three Endemic Fishes to Climate Change in Tibet. Biology, 11(12), 1808. https://doi.org/10.3390/biology11121808