

Commensal Intestinal Protozoa—Underestimated Members of the Gut Microbial Community

Abstract
Simple Summary
Abstract
1. Introduction
2. Intestinal Protozoa—Pathogenic, Commensal, or Beneficial?
2.1. Blastocystis spp.
2.2. Dientamoeba fragilis
2.3. Entamoeba spp.
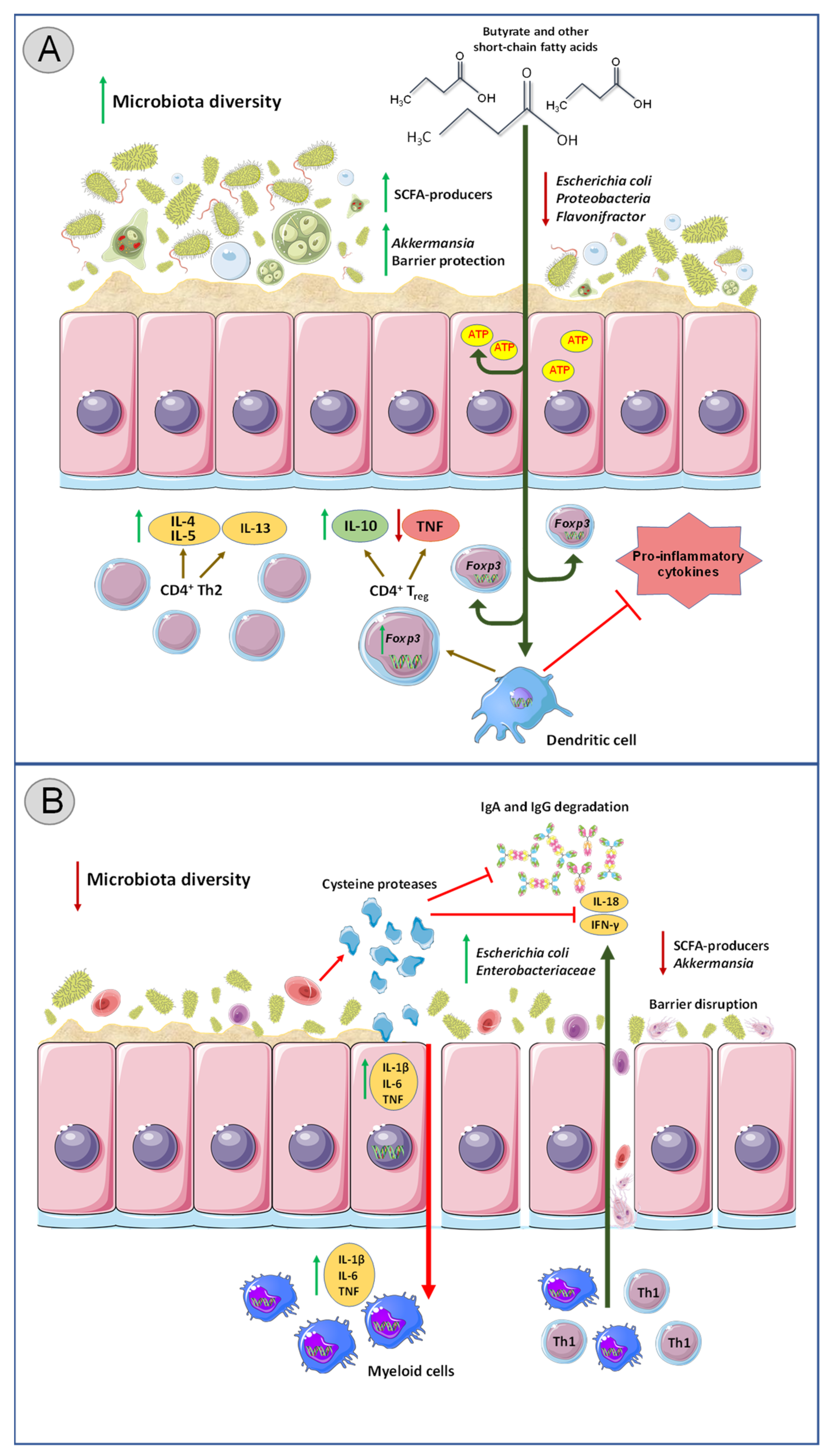
3. Protozoa–Microbiota Interactions

{kind=link}
| Protozoa spp. | Detection Method | Alterations to the Gut Bacterial Microbiota in Colonized Individuals | Ref. | Characteristics of the Enriched Bacterial Species |
|---|---|---|---|---|
| Blastocystis | Real-time PCR | Increase of bacterial genera: Acetanaerobacterium, Acetivibrio, Coprococcus, Hespellia, Oscillibacter, Papillibacter, Sporobacter, Ruminococcus, Prevotella, Roseburia, Faecalibacterium Decrease of bacterial families: Enterococcaceae, Streptococcaceae, Lactobacillaceae Enterobacteriaceae | [22] | Acetanaerobacterium: anaerobic, fermentation of acetate ethanol [116] Coprococcus: anaerobic, vitamin B, butyrate- and acetate production [111,117] Hespellia: anaerobic, butyrate production [118] Oscillibacter: anaerobic, glucose oxidation [119] Papillibacter: anaerobic, butyrate production [120] Ruminococcus: anaerobic, metabolism of complex polysaccharides [121] Prevotella: anaerobic, metabolism of polysaccharides [122] Roseburia: anaerobic, butyrate-production [120] Feacalibacterium: anaerobic, butyrate and other SCFA production [120] |
| Blastocystis (ST1-6) | Metagenomics | General increase of Firmicutes phyla and Clostridia order. Decrease of Bacteroides genus Increase of bacterial species: Methanobrevibacter smithii, Akkermansia muciniphila, Butyrivibrio crossotus, Eubacterium siraeum, Coprococcus catus, Prevotella copri, Eubacterium rectale, Bifidobacterium adolescentis, Faecalibacterium prausnitzii, Treponema succinifaciens Decrease of bacterial species: Ruminococcus gnavus, Dialister invisus, Escherichia coli, Bacteroides thetaiotamicron, Bacteroides fragilis, Bacteroides vulgatus, Bacteroides uniformis, Bacteroidesovatus, Bacteroides stercoris | [71] | Methanobrevibacter: methanogen, anaerobic, SCFA-production [96] Akkermansia municiphila: anaerobic, mucin degrading, SCFA-production [104] Butyrivibrio crossotus: anaerobic, butyrate-production [120] Eubacterium siraeum: anaerobic, degradation of xylans and ferulic acid production [123] Coprococcus catus: anaerobic, SCFA-production [117] Eubacterium rectale: anaerobic, SCFA-production [120] Bifidobacterium adolescentis: anaerobic, SCFA- and folate production [124] Treponema succinifaciens: anaerobic, SCFA-production [125] |
| Blastocystis (ST3) | Microscopic evaluation and real-time PCR | General increase of Prevotellaceae, Methanobacteriaceae, Clostridiaceae Lachnospiraceae, Erysipelotrichaceae and Pasteurellaceae family. Decrease of Bacteroidaceae and Veillonecellaceae family. Increase of bacterial genera: Prevotella, Methanobrevibacter, Ruminococcus Decrease of bacterial genera: Bacteroides | [126] | Prevotella: anaerobic, metabolism of polysaccharides [122] Methanobrevibacter: methanogen, anaerobic, SCFA-production [96] Ruminococcus: anaerobic, metabolism of complex polysaccharides [121] |
| D. fragilis | Microscopic evaluation, multiplex qPCR and real-time PCR | General Decrease of Bacteroides. Increase of bacterial genera: Akkermansia muciniphila, Methanobrevibacter smithii, Butyrivibrio crossotus, Alistipes, Victivallis, Oscillibacter, Eubacterium, Coproccus, Bifidobacterium adolescentis, Bifidobacterium longum, Ruminococcus bromii, Prevotella copri, Decrease of bacterial genera: Flavonifractor, Parabacteroides distasonis, Bacteroides fragilis,Clostridium leptum, | [73,82] | Methanobrevibacter: methanogen, anaerobic, SCFA-production [96] Akkermansia municiphila: anaerobic, mucin degrading, SCFA-production [104] Victivallis: anaerobic and sugar fermenting [127] Oscillibacter: anaerobic, glucose oxidation [119] Coprococcus: anaerobic, vitamin B, butyrate- and acetate production [111,117] Bifidobacterium adolescentis: anaerobic, SCFA- and folate production [124] Eubacterium siraeum: anaerobic, degradation of xylans and ferulic acid production [123] |
| Entamoeba spp. | Microscopic evaluation and metagenomics | General increase of taxa Clostridiales, Ruminococcaceae. Decrease of Bacteroides, Prevotella and Fusobacteria Increase of bacterial genera Akkarmensia municiphila, Coprococcus, Alistepes Decrease of bacterial genera: Blautia, Streptococcus | [21,48,86] | Alistepes: anaerobic, hydrolysis of tryptophan to indole [128] Coprococcus: anaerobic, vitamin B, butyrate- and acetate production [111,117] Akkermansia municiphila: anaerobic, mucin degrading, SCFA-production [104] |
| Entamoeba and Blastocystis | Nested-PCR | Increase of Faecalibacterium prausnitziim. Decrease of Escherichia coli | [87] | Feacalibacterium prausnitziim: anaerobic, butyrate and other SCFA production [120] |
4. The Impact of Commensal Gut Protozoa on the Host Immune System
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome—A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef]
- Xiong, J.; Hu, H.; Xu, C.; Yin, J.; Liu, M.; Zhang, L.; Duan, Y.; Huang, Y. Development of gut microbiota along with its metabolites of preschool children. BMC Pediatr. 2022, 22, 25. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef]
- Maeda, Y.; Takeda, K. Host-microbiota interactions in rheumatoid arthritis. Exp. Mol. Med. 2019, 51, 1–6. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, K.; Wu, Y.; Yang, Y.; Tso, P.; Wu, Z. Interactions between Intestinal Microbiota and Host Immune Response in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 942. [Google Scholar] [CrossRef]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef]
- Maiuolo, J.; Gliozzi, M.; Musolino, V.; Carresi, C.; Scarano, F.; Nucera, S.; Scicchitano, M.; Oppedisano, F.; Bosco, F.; Ruga, S.; et al. The Contribution of Gut Microbiota-Brain Axis in the Development of Brain Disorders. Front. Neurosci. 2021, 15, 616883. [Google Scholar] [CrossRef]
- Caruso, R.; Lo, B.C.; Núñez, G. Host-microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef]
- García, M.J.; Pascual, M.; Del Pozo, C.; Díaz-González, A.; Castro, B.; Rasines, L.; Crespo, J.; Rivero, M. Impact of immune-mediated diseases in inflammatory bowel disease and implications in therapeutic approach. Sci. Rep. 2020, 10, 10731. [Google Scholar] [CrossRef]
- Argollo, M.; Gilardi, D.; Peyrin-Biroulet, C.; Chabot, J.F.; Peyrin-Biroulet, L.; Danese, S. Comorbidities in inflammatory bowel disease: A call for action. Lancet Gastroenterol. Hepatol. 2019, 4, 643–654. [Google Scholar] [CrossRef]
- Matos, R.; Lencastre, L.; Rocha, V.; Torres, S.; Vieira, F.; Barbosa, M.R.; Ascenção, J.; Guerra, M.P. Quality of life in patients with inflammatory bowel disease: The role of positive psychological factors. Health Psychol. Behav. Med. 2021, 9, 989–1005. [Google Scholar] [CrossRef]
- Silangcruz, K.; Nishimura, Y.; Czech, T.; Kimura, N.; Hagiya, H.; Koyama, T.; Otsuka, F. Impact of the World Inflammatory Bowel Disease Day and Crohn’s and Colitis Awareness Week on Population Interest Between 2016 and 2020: Google Trends Analysis. JMIR Infodemiol. 2021, 1, e32856. [Google Scholar] [CrossRef]
- Cui, G.; Liu, H.; Xu, G.; Laugsand, J.B.; Pang, Z. Exploring Links Between Industrialization, Urbanization, and Chinese Inflammatory Bowel Disease. Front. Med. 2021, 8, 757025. [Google Scholar] [CrossRef]
- Lukeš, J.; Stensvold, C.R.; Jirků-Pomajbíková, K.; Wegener Parfrey, L. Are Human Intestinal Eukaryotes Beneficial or Commensals? PLoS Pathog. 2015, 11, e1005039. [Google Scholar] [CrossRef]
- Chudnovskiy, A.; Mortha, A.; Kana, V.; Kennard, A.; Ramirez, J.D.; Rahman, A.; Remark, R.; Mogno, I.; Ng, R.; Gnjatic, S.; et al. Host-Protozoan Interactions Protect from Mucosal Infections through Activation of the Inflammasome. Cell 2016, 167, 444–456.e414. [Google Scholar] [CrossRef]
- Wei, Y.; Gao, J.; Kou, Y.; Meng, L.; Zheng, X.; Liang, M.; Sun, H.; Liu, Z.; Wang, Y. Commensal Bacteria Impact a Protozoan’s Integration into the Murine Gut Microbiota in a Dietary Nutrient-Dependent Manner. Appl. Environ. Microbiol. 2020, 86, e00303-20. [Google Scholar] [CrossRef]
- Issa, R. Non-pathogenic protozoa. Int. J. Pharm. Pharm. Sci. 2014, 6, 30–40. [Google Scholar]
- Cavalier-Smith, T. Protist phylogeny and the high-level classification of Protozoa. Eur. J. Protistol. 2003, 39, 338–348. [Google Scholar] [CrossRef]
- Ocaña, K.A.; Dávila, A.M. Phylogenomics-based reconstruction of protozoan species tree. Evol. Bioinform. Online 2011, 7, 107–121. [Google Scholar] [CrossRef]
- Even, G.; Lokmer, A.; Rodrigues, J.; Audebert, C.; Viscogliosi, E.; Ségurel, L.; Chabé, M. Changes in the Human Gut Microbiota Associated With Colonization by Blastocystis sp. and Entamoeba spp. in Non-Industrialized Populations. Front. Cell. Infect. Microbiol. 2021, 11, 533528. [Google Scholar] [CrossRef]
- Audebert, C.; Even, G.; Cian, A.; Safadi, D.E.; Certad, G.; Delhaes, L.; Pereira, B.; Nourrisson, C.; Poirier, P.; Wawrzyniak, I.; et al. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef] [PubMed]
- von Huth, S.; Thingholm, L.B.; Kofoed, P.-E.; Bang, C.; Rühlemann, M.C.; Franke, A.; Holmskov, U. Intestinal protozoan infections shape fecal bacterial microbiota in children from Guinea-Bissau. PLoS Negl. Trop. Dis. 2021, 15, e0009232. [Google Scholar] [CrossRef]
- Chabé, M.; Lokmer, A.; Ségurel, L. Gut Protozoa: Friends or Foes of the Human Gut Microbiota? Trends Parasitol. 2017, 33, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Maritz, J.M.; Land, K.M.; Carlton, J.M.; Hirt, R.P. What is the importance of zoonotic trichomonads for human health? Trends Parasitol. 2014, 30, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Gascoigne, N.R.J.; Peng, G.; Tan, K.S.W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 2021, 17, e1009253. [Google Scholar] [CrossRef]
- Wawrzyniak, I.; Poirier, P.; Viscogliosi, E.; Dionigia, M.; Texier, C.; Delbac, F.; Alaoui, H.E. Blastocystis, an unrecognized parasite: An overview of pathogenesis and diagnosis. Ther. Adv. Infect. Dis. 2013, 1, 167–178. [Google Scholar] [CrossRef]
- Leung, J.M.; Graham, A.L.; Knowles, S.C.L. Parasite-Microbiota Interactions With the Vertebrate Gut: Synthesis Through an Ecological Lens. Front. Microbiol. 2018, 9, 843. [Google Scholar] [CrossRef]
- Panthee, B.; Gyawali, S.; Panthee, P.; Techato, K. Environmental and Human Microbiome for Health. Life 2022, 12, 456. [Google Scholar] [CrossRef]
- Thompson, R.C.A.; Ash, A. Molecular epidemiology of Giardia and Cryptosporidium infections. Infect. Genet. Evol. 2016, 40, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Kurenzvi, L.; Sebunya, T.K.; Coetzee, T.; Paganotti, G.M.; Teye, M.V. Prevalence of Cryptosporidium parvum, Giardia intestinalis and molecular characterization of group A rotavirus associated with diarrhea in children below five years old in Gaborone, Botswana. Pan. Afr. Med. J. 2020, 37, 159. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Padalia, J.; Moonah, S. Tissue Destruction Caused by Entamoeba histolytica Parasite: Cell Death, Inflammation, Invasion, and the Gut Microbiome. Curr. Clin. Microbiol. Rep. 2019, 6, 51–57. [Google Scholar] [CrossRef]
- Guérin, A.; Striepen, B. The Biology of the Intestinal Intracellular Parasite Cryptosporidium. Cell Host. Microbe. 2020, 28, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.D. Giardia duodenalis: Biology and Pathogenesis. Clin. Microbiol. Rev. 2021, 34, e00024-19. [Google Scholar] [CrossRef]
- Marie, C.; Petri, W.A., Jr. Regulation of virulence of Entamoeba histolytica. Annu. Rev. Microbiol. 2014, 68, 493–520. [Google Scholar] [CrossRef]
- Lokmer, A.; Cian, A.; Froment, A.; Gantois, N.; Viscogliosi, E.; Chabé, M.; Ségurel, L. Use of shotgun metagenomics for the identification of protozoa in the gut microbiota of healthy individuals from worldwide populations with various industrialization levels. PLoS ONE 2019, 14, e0211139. [Google Scholar] [CrossRef]
- Parfrey, L.W.; Walters, W.A.; Lauber, C.L.; Clemente, J.C.; Berg-Lyons, D.; Teiling, C.; Kodira, C.; Mohiuddin, M.; Brunelle, J.; Driscoll, M.; et al. Communities of microbial eukaryotes in the mammalian gut within the context of environmental eukaryotic diversity. Front. Microbiol. 2014, 5, 298. [Google Scholar] [CrossRef]
- Schuster, F.L.; Ramirez-Avila, L. Current world status of Balantidium coli. Clin. Microbiol. Rev. 2008, 21, 626–638. [Google Scholar] [CrossRef]
- Andersen, L.O.B.; Stensvold, C.R. Blastocystis in Health and Disease: Are We Moving from a Clinical to a Public Health Perspective? J. Clin. Microbiol. 2016, 54, 524–528. [Google Scholar] [CrossRef]
- Mathison, B.A.; Pritt, B.S. Parasites of the Gastrointestinal Tract. In Encyclopedia of Infection and Immunity; Rezaei, N., Ed.; Elsevier: Oxford, UK, 2022; pp. 136–203. [Google Scholar] [CrossRef]
- Dubey, J.P.; Almeria, S. Cystoisospora belli infections in humans: The past 100 years. Parasitology 2019, 146, 1490–1527. [Google Scholar] [CrossRef]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium species in humans and animals: Current understanding and research needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef]
- Almeria, S.; Cinar, H.N.; Dubey, J.P. Cyclospora cayetanensis and Cyclosporiasis: An Update. Microorganisms 2019, 7, 317. [Google Scholar] [CrossRef]
- Stark, D.; Barratt, J.; Chan, D.; Ellis, J.T. Dientamoeba fragilis, the Neglected Trichomonad of the Human Bowel. Clin. Microbiol. Rev. 2016, 29, 553–580. [Google Scholar] [CrossRef]
- Poulsen, C.S.; Stensvold, C.R. Systematic review on Endolimax nana: A less well studied intestinal ameba. Trop. Parasitol. 2016, 6, 8–29. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, C.A. Entamoeba bangladeshi: An insight. Trop. Parasitol. 2014, 4, 96–98. [Google Scholar] [CrossRef] [PubMed]
- Alzate, J.F.; Toro-Londoño, M.; Cabarcas, F.; Garcia-Montoya, G.; Galvan-Diaz, A. Contrasting microbiota profiles observed in children carrying either Blastocystis spp. or the commensal amoebas Entamoeba coli or Endolimax nana. Sci. Rep. 2020, 10, 15354. [Google Scholar] [CrossRef]
- Pysova, I.; Tumova, P.; Tolarova, V.; Nohynkova, E. Nonpathogenic Entamoeba dispar quickly outgrows pathogenic Entamoeba histolytica in mixed xenic cultures. Lett. Appl. Microbiol. 2009, 48, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Winiecka-Krusnell, J.; Lier, T.; Lebbad, M. Evaluation of a PCR Method for Detection of Entamoeba polecki, with an Overview of Its Molecular Epidemiology. J. Clin. Microbiol. 2018, 56, e00154-18. [Google Scholar] [CrossRef]
- Singh, A.; Banerjee, T.; Khan, U.; Shukla, S.K. Epidemiology of clinically relevant Entamoeba spp. (E. histolytica/dispar/moshkovskii/bangladeshi): A cross sectional study from North India. PLoS Negl. Trop. Dis. 2021, 15, e0009762. [Google Scholar] [CrossRef]
- Li, W.C.; Ying, M.; Gong, P.T.; Li, J.H.; Yang, J.; Li, H.; Zhang, X.C. Pentatrichomonas hominis: Prevalence and molecular characterization in humans, dogs, and monkeys in Northern China. Parasitol. Res. 2016, 115, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; van der Giezen, M. Associations between Gut Microbiota and Common Luminal Intestinal Parasites. Trends Parasitol. 2018, 34, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Nourrisson, C.; Scanzi, J.; Pereira, B.; NkoudMongo, C.; Wawrzyniak, I.; Cian, A.; Viscogliosi, E.; Livrelli, V.; Delbac, F.; Dapoigny, M.; et al. Blastocystis is associated with decrease of fecal microbiota protective bacteria: Comparative analysis between patients with irritable bowel syndrome and control subjects. PLoS ONE 2014, 9, e111868. [Google Scholar] [CrossRef]
- Poirier, P.; Wawrzyniak, I.; Vivarès, C.P.; Delbac, F.; El Alaoui, H. New insights into Blastocystis spp.: A potential link with irritable bowel syndrome. PLoS Pathog. 2012, 8, e1002545. [Google Scholar] [CrossRef]
- Shafiei, Z.; Esfandiari, F.; Sarkari, B.; Rezaei, Z.; Fatahi, M.R.; Hosseini Asl, S.M.K. Parasitic infections in irritable bowel syndrome patients: Evidence to propose a possible link, based on a case-control study in the south of Iran. BMC Res. Notes 2020, 13, 264. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Lepczyńska, M.; Kot, K.; Szkup, M.; Łanocha-Arendarczyk, N.; Dzika, E.; Grochans, E. Prevalence, subtypes and risk factors of Blastocystis spp. infection among pre- and perimenopausal women. BMC Infect. Dis. 2021, 21, 1125. [Google Scholar] [CrossRef]
- Petersen, A.M.; Stensvold, C.R.; Mirsepasi, H.; Engberg, J.; Friis-Møller, A.; Porsbo, L.J.; Hammerum, A.M.; Nordgaard-Lassen, I.; Nielsen, H.V.; Krogfelt, K.A. Active ulcerative colitis associated with low prevalence of Blastocystis and Dientamoeba fragilis infection. Scand. J. Gastroenterol. 2013, 48, 638–639. [Google Scholar] [CrossRef] [PubMed]
- Leder, K.; Hellard, M.E.; Sinclair, M.I.; Fairley, C.K.; Wolfe, R. No correlation between clinical symptoms and Blastocystis hominis in immunocompetent individuals. J. Gastroenterol. Hepatol. 2005, 20, 1390–1394. [Google Scholar] [CrossRef]
- Krogsgaard, L.R.; Engsbro, A.L.; Stensvold, C.R.; Nielsen, H.V.; Bytzer, P. The prevalence of intestinal parasites is not greater among individuals with irritable bowel syndrome: A population-based case-control study. Clin. Gastroenterol. Hepatol. 2015, 13, 507–513.e502. [Google Scholar] [CrossRef]
- Gassama, A.; Sow, P.S.; Fall, F.; Camara, P.; Guèye-N’diaye, A.; Seng, R.; Samb, B.; M’Boup, S.; Aïdara-Kane, A. Ordinary and opportunistic enteropathogens associated with diarrhea in Senegalese adults in relation to human immunodeficiency virus serostatus. Int. J. Infect. Dis. 2001, 5, 192–198. [Google Scholar] [CrossRef]
- Sarzhanov, F.; Dogruman-Al, F.; Santin, M.; Maloney, J.G.; Gureser, A.S.; Karasartova, D.; Taylan-Ozkan, A. Investigation of neglected protists Blastocystis sp. and Dientamoeba fragilis in immunocompetent and immunodeficient diarrheal patients using both conventional and molecular methods. PLoS Negl. Trop. Dis. 2021, 15, e0009779. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.D.; Sánchez, L.V.; Bautista, D.C.; Corredor, A.F.; Flórez, A.C.; Stensvold, C.R. Blastocystis subtypes detected in humans and animals from Colombia. Infect. Genet. Evol. 2014, 22, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Stensvold, C.R.; Rajilić-Stojanović, M.; Heilig, H.G.; De Vos, W.M.; O’Toole, P.W.; Cotter, P.D. The microbial eukaryote Blastocystis is a prevalent and diverse member of the healthy human gut microbiota. FEMS Microbiol. Ecol. 2014, 90, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Sulżyc-Bielicka, V.; Kołodziejczyk, L.; Adamska, M.; Skotarczak, B.; Jaczewska, S.; Safranow, K.; Bielicki, P.; Kładny, J.; Bielicki, D. Colorectal cancer and Blastocystis sp. infection. Parasit. Vectors 2021, 14, 200. [Google Scholar] [CrossRef]
- Dogruman-Al, F.; Dagci, H.; Yoshikawa, H.; Kurt, O.; Demirel, M. A possible link between subtype 2 and asymptomatic infections of Blastocystis hominis. Parasitol. Res. 2008, 103, 685–689. [Google Scholar] [CrossRef]
- Khademvatan, S.; Masjedizadeh, R.; Yousefi-Razin, E.; Mahbodfar, H.; Rahim, F.; Yousefi, E.; Foroutan, M. PCR-based molecular characterization of Blastocystis hominis subtypes in southwest of Iran. J. Infect. Public Health 2018, 11, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.S.; Whipps, C.M.; Ganac, R.D.; Hudson, N.R.; Boorom, K. Association of Blastocystis subtype 3 and 1 with patients from an Oregon community presenting with chronic gastrointestinal illness. Parasitol. Res. 2009, 104, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.H.; Ismail, M.A.M.; El-Badry, A.A.; Abu-Sarea, E.Y.; Dewidar, A.M.; Hamdy, D.A. An Association Between Blastocystis Subtypes and Colorectal Cancer Patients: A Significant Different Profile from Non-cancer Individuals. Acta Parasitol. 2022, 67, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Kodio, A.; Coulibaly, D.; Koné, A.K.; Konaté, S.; Doumbo, S.; Guindo, A.; Bittar, F.; Gouriet, F.; Raoult, D.; Thera, M.A.; et al. Blastocystis Colonization Is Associated with Increased Diversity and Altered Gut Bacterial Communities in Healthy Malian Children. Microorganisms 2019, 7, 649. [Google Scholar] [CrossRef]
- Beghini, F.; Pasolli, E.; Truong, T.D.; Putignani, L.; Cacciò, S.M.; Segata, N. Large-scale comparative metagenomics of Blastocystis, a common member of the human gut microbiome. ISME J. 2017, 11, 2848–2863. [Google Scholar] [CrossRef]
- Rossen, N.G.; Bart, A.; Verhaar, N.; van Nood, E.; Kootte, R.; de Groot, P.F.; D’Haens, G.R.; Ponsioen, C.Y.; van Gool, T. Low prevalence of Blastocystis sp. in active ulcerative colitis patients. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Krogsgaard, L.R.; Andersen, L.O.B.; Johannesen, T.B.; Engsbro, A.L.; Stensvold, C.R.; Nielsen, H.V.; Bytzer, P. Characteristics of the bacterial microbiome in association with common intestinal parasites in irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2018, 9, e161. [Google Scholar] [CrossRef] [PubMed]
- Tito, R.Y.; Chaffron, S.; Caenepeel, C.; Lima-Mendez, G.; Wang, J.; Vieira-Silva, S.; Falony, G.; Hildebrand, F.; Darzi, Y.; Rymenans, L.; et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut 2019, 68, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Billy, V.; Lhotská, Z.; Jirků, M.; Kadlecová, O.; Frgelecová, L.; Parfrey, L.W.; Pomajbíková, K.J. Blastocystis Colonization Alters the Gut Microbiome and, in Some Cases, Promotes Faster Recovery From Induced Colitis. Front. Microbiol. 2021, 12, 641483. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Png, C.W.; Koh, E.Y.; Aung, T.T.; Kioh, D.Y.Q.; Chan, E.C.Y.; Malleret, B.; Zhang, Y.; Peng, G.; et al. Experimental colonization with Blastocystis ST4 is associated with protective immune responses and modulation of gut microbiome in a DSS-induced colitis mouse model. Cell Mol. Life Sci. 2022, 79, 245. [Google Scholar] [CrossRef]
- Lepczyńska, M.; Białkowska, J.; Dzika, E.; Piskorz-Ogórek, K.; Korycińska, J. Blastocystis: How do specific diets and human gut microbiota affect its development and pathogenicity? Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1531–1540. [Google Scholar] [CrossRef] [PubMed]
- Cacciò, S.M. Molecular epidemiology of Dientamoeba fragilis. Acta Trop. 2018, 184, 73–77. [Google Scholar] [CrossRef]
- Röser, D.; Simonsen, J.; Nielsen, H.V.; Stensvold, C.R.; Mølbak, K. Dientamoeba fragilis in Denmark: Epidemiological experience derived from four years of routine real-time PCR. Eur. J. Clin. Microbiol. 2013, 32, 1303–1310. [Google Scholar] [CrossRef]
- Bruijnesteijn van Coppenraet, L.E.; Dullaert-de Boer, M.; Ruijs, G.J.; van der Reijden, W.A.; van der Zanden, A.G.; Weel, J.F.; Schuurs, T.A. Case-control comparison of bacterial and protozoan microorganisms associated with gastroenteritis: Application of molecular detection. Clin. Microbiol. Infect. 2015, 21, 592.e9–592.e19. [Google Scholar] [CrossRef]
- Brands, M.R.; Van de Vijver, E.; Haisma, S.M.; Heida, A.; van Rheenen, P.F. No association between abdominal pain and Dientamoeba in Dutch and Belgian children. Arch. Dis. Child. 2019, 104, 686–689. [Google Scholar] [CrossRef]
- Gotfred-Rasmussen, H.; Stensvold, C.R.; Ingham, A.C.; Johannesen, T.B.; Andersen, L.O.B.; Röser, D.; Nielsen, H.V. Impact of Metronidazole Treatment and Dientamoeba Fragilis Colonization on Gut Microbiota Diversity. J. Pediatr. Gastroenterol. Nutr. 2021, 73, 23–29. [Google Scholar] [CrossRef] [PubMed]
- López, M.C.; León, C.M.; Fonseca, J.; Reyes, P.; Moncada, L.; Olivera, M.J.; Ramírez, J.D. Molecular Epidemiology of Entamoeba: First Description of Entamoeba moshkovskii in a Rural Area from Central Colombia. PLoS ONE 2015, 10, e0140302. [Google Scholar] [CrossRef] [PubMed][Green Version]
- El-Dib, N.A.; Khater, M.M. Entamoeba. In Encyclopedia of Infection and Immunity; Rezaei, N., Ed.; Elsevier: Oxford, UK, 2022; pp. 492–512. [Google Scholar] [CrossRef]
- Cui, Z.; Li, J.; Chen, Y.; Zhang, L. Molecular epidemiology, evolution, and phylogeny of Entamoeba spp. Infect. Genet. Evol. 2019, 75, 104018. [Google Scholar] [CrossRef] [PubMed]
- Morton, E.R.; Lynch, J.; Froment, A.; Lafosse, S.; Heyer, E.; Przeworski, M.; Blekhman, R.; Ségurel, L. Variation in Rural African Gut Microbiota Is Strongly Correlated with Colonization by Entamoeba and Subsistence. PLoS Genet. 2015, 11, e1005658. [Google Scholar] [CrossRef] [PubMed]
- Valerio, I.; Floriana, S.; Valentina, T.; Fabrizio, P.; Anatole, M.; Veronica Di, C.; David Di, C.; Serena, S.; Federica, B.; Rossella, D.A. Gut microbiota related to Giardia duodenalis, Entamoeba spp. and Blastocystis hominis infections in humans from Côte d’Ivoire. J. Infect. Dev. Ctries. 2016, 10, 8179. [Google Scholar] [CrossRef]
- Mejia, R.; Damania, A.; Jeun, R.; Bryan, P.E.; Vargas, P.; Juarez, M.; Cajal, P.S.; Nasser, J.; Krolewiecki, A.; Lefoulon, E.; et al. Impact of intestinal parasites on microbiota and cobalamin gene sequences: A pilot study. Parasit. Vectors 2020, 13, 200. [Google Scholar] [CrossRef]
- Chihi, A.; O’Brien Andersen, L.; Aoun, K.; Bouratbine, A.; Stensvold, C.R. Amplicon-based next-generation sequencing of eukaryotic nuclear ribosomal genes (metabarcoding) for the detection of single-celled parasites in human faecal samples. Parasit. Epidemiol. Control. 2022, 17, e00242. [Google Scholar] [CrossRef]
- Weber, A.A.T.; Pawlowski, J. Can Abundance of Protists Be Inferred from Sequence Data: A Case Study of Foraminifera. PLoS ONE 2013, 8, e56739. [Google Scholar] [CrossRef]
- Vieira-Silva, S.; Falony, G.; Darzi, Y.; Lima-Mendez, G.; Garcia Yunta, R.; Okuda, S.; Vandeputte, D.; Valles-Colomer, M.; Hildebrand, F.; Chaffron, S.; et al. Species–function relationships shape ecological properties of the human gut microbiome. Nat. Microbiol. 2016, 1, 16088. [Google Scholar] [CrossRef]
- O’Brien Andersen, L.; Karim, A.B.; Roager, H.M.; Vigsnæs, L.K.; Krogfelt, K.A.; Licht, T.R.; Stensvold, C.R. Associations between common intestinal parasites and bacteria in humans as revealed by qPCR. Eur. J. Clin. Microbiol. 2016, 35, 1427–1431. [Google Scholar] [CrossRef]
- Baldelli, V.; Scaldaferri, F.; Putignani, L.; Del Chierico, F. The Role of Enterobacteriaceae in Gut Microbiota Dysbiosis in Inflammatory Bowel Diseases. Microorganisms 2021, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Vester-Andersen, M.K.; Mirsepasi-Lauridsen, H.C.; Prosberg, M.V.; Mortensen, C.O.; Träger, C.; Skovsen, K.; Thorkilgaard, T.; Nøjgaard, C.; Vind, I.; Krogfelt, K.A.; et al. Increased abundance of proteobacteria in aggressive Crohn’s disease seven years after diagnosis. Sci. Rep. 2019, 9, 13473. [Google Scholar] [CrossRef] [PubMed]
- Camara, A.; Konate, S.; Tidjani Alou, M.; Kodio, A.; Togo, A.H.; Cortaredona, S.; Henrissat, B.; Thera, M.A.; Doumbo, O.K.; Raoult, D.; et al. Clinical evidence of the role of Methanobrevibacter smithii in severe acute malnutrition. Sci. Rep. 2021, 11, 5426. [Google Scholar] [CrossRef]
- Geirnaert, A.; Calatayud, M.; Grootaert, C.; Laukens, D.; Devriese, S.; Smagghe, G.; De Vos, M.; Boon, N.; Van de Wiele, T. Butyrate-producing bacteria supplemented in vitro to Crohn’s disease patient microbiota increased butyrate production and enhanced intestinal epithelial barrier integrity. Sci. Rep. 2017, 7, 11450. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Tong, M.; Li, X.; Wegener Parfrey, L.; Roth, B.; Ippoliti, A.; Wei, B.; Borneman, J.; McGovern, D.P.B.; Frank, D.N.; Li, E.; et al. A Modular Organization of the Human Intestinal Mucosal Microbiota and Its Association with Inflammatory Bowel Disease. PLoS ONE 2013, 8, e80702. [Google Scholar] [CrossRef]
- Sokol, H.; Seksik, P.; Furet, J.P.; Firmesse, O.; Nion-Larmurier, I.; Beaugerie, L.; Cosnes, J.; Corthier, G.; Marteau, P.; Doré, J. Low Counts of Faecalibacterium prausnitzii in Colitis Microbiota. Inflamm. Bowel. Dis. 2009, 15, 1183–1189. [Google Scholar] [CrossRef]
- Caudet, J.; Trelis, M.; Cifre, S.; Soriano, J.M.; Rico, H.; Merino-Torres, J.F. Interplay between Intestinal Bacterial Communities and Unicellular Parasites in a Morbidly Obese Population: A Neglected Trinomial. Nutrients 2022, 14, 3211. [Google Scholar] [CrossRef]
- Yao, S.; Zhao, Z.; Wang, W.; Liu, X. Bifidobacterium Longum: Protection against Inflammatory Bowel Disease. J. Immunol. Res. 2021, 2021, 8030297. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Belzer, C.; Chia, L.W.; Aalvink, S.; Chamlagain, B.; Piironen, V.; Knol, J.; de Vos, W.M. Microbial Metabolic Networks at the Mucus Layer Lead to Diet-Independent Butyrate and Vitamin B(12) Production by Intestinal Symbionts. mBio 2017, 8, e00770-17. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Lee, J.W.J.; Tan, K.S.W. Infection with pathogenic Blastocystis ST7 is associated with decreased bacterial diversity and altered gut microbiome profiles in diarrheal patients. Parasit. Vectors 2022, 15, 312. [Google Scholar] [CrossRef] [PubMed]
- Freeman, C.D.; Klutman, N.E.; Lamp, K.C. Metronidazole. A therapeutic review and update. Drugs 1997, 54, 679–708. [Google Scholar] [CrossRef]
- Gupta, A.; Dhakan, D.B.; Maji, A.; Saxena, R.; P K, V.P.; Mahajan, S.; Pulikkan, J.; Kurian, J.; Gomez, A.M.; Scaria, J.; et al. Association of Flavonifractor plautii, a Flavonoid-Degrading Bacterium, with the Gut Microbiome of Colorectal Cancer Patients in India. mSystems 2019, 4, e00438-19. [Google Scholar] [CrossRef]
- Yang, Y.; Du, L.; Shi, D.; Kong, C.; Liu, J.; Liu, G.; Li, X.; Ma, Y. Dysbiosis of human gut microbiome in young-onset colorectal cancer. Nat. Commun. 2021, 12, 6757. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Y.; Gu, J. Clinical Implications of the Associations Between Intestinal Microbiome and Colorectal Cancer Progression. Cancer Manag. Res. 2020, 12, 4117–4128. [Google Scholar] [CrossRef]
- Doumatey, A.P.; Adeyemo, A.; Zhou, J.; Lei, L.; Adebamowo, S.N.; Adebamowo, C.; Rotimi, C.N. Gut Microbiome Profiles Are Associated With Type 2 Diabetes in Urban Africans. Front. Cell. Infect. Microbiol. 2020, 10, 63. [Google Scholar] [CrossRef]
- Kort, R.; Schlösser, J.; Vazquez, A.R.; Atukunda, P.; Muhoozi, G.K.M.; Wacoo, A.P.; Sybesma, W.F.H.; Westerberg, A.C.; Iversen, P.O.; Schoen, E.D. Model Selection Reveals the Butyrate-Producing Gut Bacterium Coprococcus eutactus as Predictor for Language Development in 3-Year-Old Rural Ugandan Children. Front. Microbiol. 2021, 12, 681485. [Google Scholar] [CrossRef]
- Sobhani, I.; Bergsten, E.; Couffin, S.; Amiot, A.; Nebbad, B.; Barau, C.; de’Angelis, N.; Rabot, S.; Canoui-Poitrine, F.; Mestivier, D.; et al. Colorectal cancer-associated microbiota contributes to oncogenic epigenetic signatures. Proc. Natl. Acad. Sci. USA 2019, 116, 24285–24295. [Google Scholar] [CrossRef]
- Charania, R.; Wade, B.E.; McNair, N.N.; Mead, J.R. Changes in the Microbiome of Cryptosporidium-Infected Mice Correlate to Differences in Susceptibility and Infection Levels. Microorganisms 2020, 8, 879. [Google Scholar] [CrossRef] [PubMed]
- Olajide, J.S.; Cai, J. Perils and Promises of Pathogenic Protozoan Extracellular Vesicles. Front. Cell. Infect. Microbiol. 2020, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- Espinoza-Vergara, G.; Hoque, M.M.; McDougald, D.; Noorian, P. The Impact of Protozoan Predation on the Pathogenicity of Vibrio cholerae. Front. Microbiol. 2020, 11, 17. [Google Scholar] [CrossRef]
- Chen, S.Y.; Niu, L.L.; Dong, X.Z. Hydrogen production from glucose by Acetanaerobacterium elongatum. Wei Sheng Wu Xue Bao 2006, 46, 233–237. [Google Scholar] [PubMed]
- Nogal, A.; Louca, P.; Zhang, X.; Wells, P.M.; Steves, C.J.; Spector, T.D.; Falchi, M.; Valdes, A.M.; Menni, C. Circulating Levels of the Short-Chain Fatty Acid Acetate Mediate the Effect of the Gut Microbiome on Visceral Fat. Front. Microbiol. 2021, 12, 711359. [Google Scholar] [CrossRef]
- Ghosh, S.; Pramanik, S. Structural diversity, functional aspects and future therapeutic applications of human gut microbiome. Arch. Microbiol. 2021, 203, 5281–5308. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kim, H.-E.; Park, J.I.; Cho, H.; Kwak, M.-J.; Kim, B.-Y.; Yang, S.H.; Lee, J.P.; Kim, D.K.; Joo, K.W.; et al. The Association between Gut Microbiota and Uremia of Chronic Kidney Disease. Microorganisms 2020, 8, 907. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Belzer, C.; Goossens, M.; Kleerebezem, M.; De Vos, W.M.; Thas, O.; De Weirdt, R.; Kerckhof, F.-M.; Van de Wiele, T. Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 2013, 7, 949–961. [Google Scholar] [CrossRef] [PubMed]
- La Reau, A.J.; Suen, G. The Ruminococci: Key symbionts of the gut ecosystem. J. Microbiol. 2018, 56, 199–208. [Google Scholar] [CrossRef]
- Precup, G.; Vodnar, D.C. Gut Prevotella as a possible biomarker of diet and its eubiotic versus dysbiotic roles: A comprehensive literature review. Br. J. Nutr. 2019, 122, 131–140. [Google Scholar] [CrossRef]
- Duncan, S.H.; Russell, W.R.; Quartieri, A.; Rossi, M.; Parkhill, J.; Walker, A.W.; Flint, H.J. Wheat bran promotes enrichment within the human colonic microbiota of butyrate-producing bacteria that release ferulic acid. Environ. Microbiol. 2016, 18, 2214–2225. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Tanes, C.; Walker, E.M.; Slisarenko, N.; Gerrets, G.L.; Grasperge, B.F.; Qin, X.; Jazwinski, S.M.; Bushman, F.D.; Bittinger, K.; Rout, N. Gut Microbiome Changes Associated with Epithelial Barrier Damage and Systemic Inflammation during Antiretroviral Therapy of Chronic SIV Infection. Viruses 2021, 13, 1567. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, S.; Furzi, F.; Fontanelli Sulekova, L.; Taliani, G.; Mattiucci, S. Occurrence of Blastocystis-subtypes in patients from Italy revealed association of ST3 with a healthy gut microbiota. Parasite Epidemiol. Control 2020, 9, e00134. [Google Scholar] [CrossRef]
- van Passel, M.W.; Kant, R.; Palva, A.; Lucas, S.; Copeland, A.; Lapidus, A.; Glavina del Rio, T.; Dalin, E.; Tice, H.; Bruce, D.; et al. Genome sequence of Victivallis vadensis ATCC BAA-548, an anaerobic bacterium from the phylum Lentisphaerae, isolated from the human gastrointestinal tract. J. Bacteriol. 2011, 193, 2373–2374. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; van Harsselaar, J.; et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Carretta, M.D.; Quiroga, J.; López, R.; Hidalgo, M.A.; Burgos, R.A. Participation of Short-Chain Fatty Acids and Their Receptors in Gut Inflammation and Colon Cancer. Front. Physiol. 2021, 12, 662739. [Google Scholar] [CrossRef]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Deng, L.; Tan, K.S.W. Interactions between Blastocystis subtype ST4 and gut microbiota in vitro. Parasit. Vectors 2022, 15, 80. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.X.; Png, C.W.; Tay, C.Y.; Teo, J.D.; Jiao, H.; Lehming, N.; Tan, K.S.; Zhang, Y. Differential regulation of proinflammatory cytokine expression by mitogen-activated protein kinases in macrophages in response to intestinal parasite infection. Infect. Immun. 2014, 82, 4789–4801. [Google Scholar] [CrossRef] [PubMed]
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine proteases in protozoan parasites. PLoS Negl. Trop. Dis. 2018, 12, e0006512. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-López, S.G.; Herdman, S.; Hirata, K.; Choi, M.H.; Choe, Y.; Craik, C.; Caffrey, C.R.; Hansell, E.; Chávez-Munguía, B.; Chen, Y.T.; et al. Use of recombinant Entamoeba histolytica cysteine proteinase 1 to identify a potent inhibitor of amebic invasion in a human colonic model. Eukaryot. Cell 2007, 6, 1130–1136. [Google Scholar] [CrossRef]
- Takeuchi, A.; Phillips, B.P. Electron Microscope Studies of Experimental Entamoeba Histolytica Infection in the Guinea Pig: I. Penetration of the Intestinal Epithelium by Trophozoites. Am. J. Trop. Med. 1975, 24, 34–48. [Google Scholar] [CrossRef]
- Wu, Z.; Mirza, H.; Tan, K.S.W. Intra-Subtype Variation in Enteroadhesion Accounts for Differences in Epithelial Barrier Disruption and Is Associated with Metronidazole Resistance in Blastocystis Subtype-7. PLoS Negl. Trop. Dis. 2014, 8, e2885. [Google Scholar] [CrossRef]
- Puthia, M.K.; Vaithilingam, A.; Lu, J.; Tan, K.S. Degradation of human secretory immunoglobulin A by Blastocystis. Parasitol. Res. 2005, 97, 386–389. [Google Scholar] [CrossRef]
- Tan, P.; Li, X.; Shen, J.; Feng, Q. Fecal Microbiota Transplantation for the Treatment of Inflammatory Bowel Disease: An Update. Front. Pharmacol. 2020, 11, 574533. [Google Scholar] [CrossRef]
- Terveer, E.M.; van Gool, T.; Ooijevaar, R.E.; Sanders, I.; Boeije-Koppenol, E.; Keller, J.J.; Bart, A.; Kuijper, E.J. Human Transmission of Blastocystis by Fecal Microbiota Transplantation Without Development of Gastrointestinal Symptoms in Recipients. Clin. Infect. Dis. 2020, 71, 2630–2636. [Google Scholar] [CrossRef]

| Intestinal Protozoa | Characteristics | Ref. |
|---|---|---|
| Balantidium coli | Pathogenic ciliates of the human cecum and colon. Causes balantidiosis in humans. | [39] |
| Blastocystis hominis | Questionable pathogenicity. Conflicting effects reported in humans. | [40] |
| Chilomastix mesnilii | Non-pathogenic flagellates of the human large and small intestine. Unknown role and impact on human health. | [41] |
| Cystoisospora belli | Pathogenic coccidians that infects epithelial cells of the intestine. Causes cystoisosporiasis in humans. | [42] |
| Cryptosporidium spp. | Pathogenic coccidians of the small intestine comprising 20 different species identified in humans. Cause cryptosporidiosis in humans. | [43] |
| Cyclospora cayetanensis | Pathogenic coccidians of the human small intestine. Causes cyclosporiasis in humans. | [44] |
| Dientamoeba fragilis | Questionable pathogenicity. Flagellates of the human large intestine. Conflicting effects reported in humans. | [45] |
| Endolimax nana | Non-pathogenic amoebas of the human large intestine. Unknown role and impact on human health. | [46] |
| Entamoeba bangladeshi | Questionable pathogenicity. Amoebas of the human large intestine. Conflicting effects reported in humans. | [47] |
| Entamoeba coli | Non-pathogenic amoebas of the human large intestine. Unknown role and impact on human health. | [48] |
| Entamoeba dispar | Non-pathogenic amoebas of the human large intestine. Unknown role and impact on human health. | [49] |
| Entamoeba hartmanni | Non-pathogenic amoebas of the human large intestine. Unknown role and impact on human health. | [18] |
| Entamoeba histolytica | Pathogenic amoebas of the human intestine. Causes amebiasis in humans. | [50] |
| Entamoeba moshkovskii | Questionable pathogenicity. Amoebas of the human large intestine. Conflicting effects reported in humans. | [51] |
| Entamoeba polecki | Non-pathogenic amoebas of the human large intestine. Unknown role and impact on human health. | [50] |
| Giardia intestinalis | Pathogenic flagellates of the human small intestine. Causes giardiasis in humans. | [35] |
| Iodamoeba butschlii | Non-pathogenic amoebas of the human large intestine. Unknown role and impact on human health | [41] |
| Retortamonas intestinalis | Non-pathogenic flagellates of the human large intestine. Unknown role and impact on human health. | [41] |
| Pentatrichomonas hominis | Questionable pathogenicity. Flagellates of the human large intestine. Unknown role and impact on human health. | [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubik, M.; Pilecki, B.; Moeller, J.B. Commensal Intestinal Protozoa—Underestimated Members of the Gut Microbial Community. Biology 2022, 11, 1742. https://doi.org/10.3390/biology11121742
Dubik M, Pilecki B, Moeller JB. Commensal Intestinal Protozoa—Underestimated Members of the Gut Microbial Community. Biology. 2022; 11(12):1742. https://doi.org/10.3390/biology11121742
Chicago/Turabian StyleDubik, Magdalena, Bartosz Pilecki, and Jesper Bonnet Moeller. 2022. "Commensal Intestinal Protozoa—Underestimated Members of the Gut Microbial Community" Biology 11, no. 12: 1742. https://doi.org/10.3390/biology11121742
APA StyleDubik, M., Pilecki, B., & Moeller, J. B. (2022). Commensal Intestinal Protozoa—Underestimated Members of the Gut Microbial Community. Biology, 11(12), 1742. https://doi.org/10.3390/biology11121742